
Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna


Abstract
:1. Introduction
2. Results
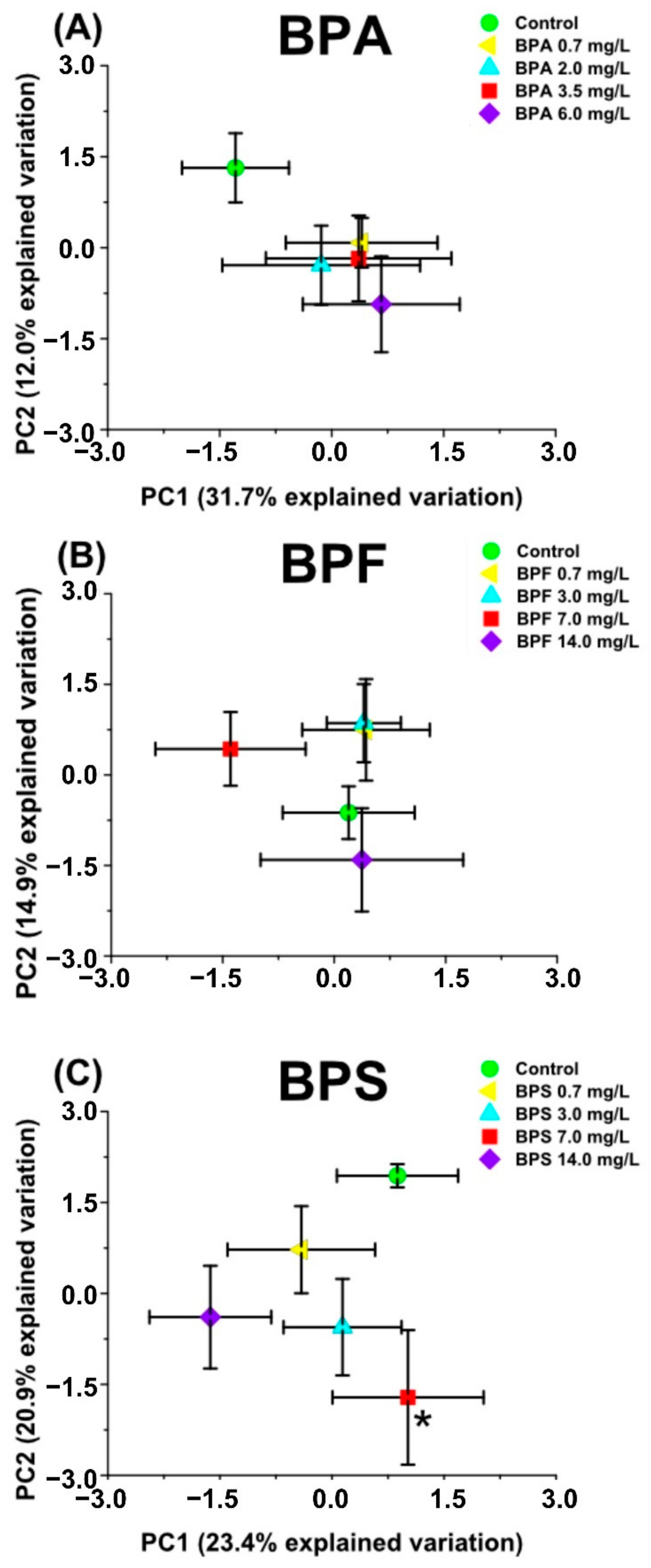
2.1. Multivariate Analysis with Bisphenol Exposure
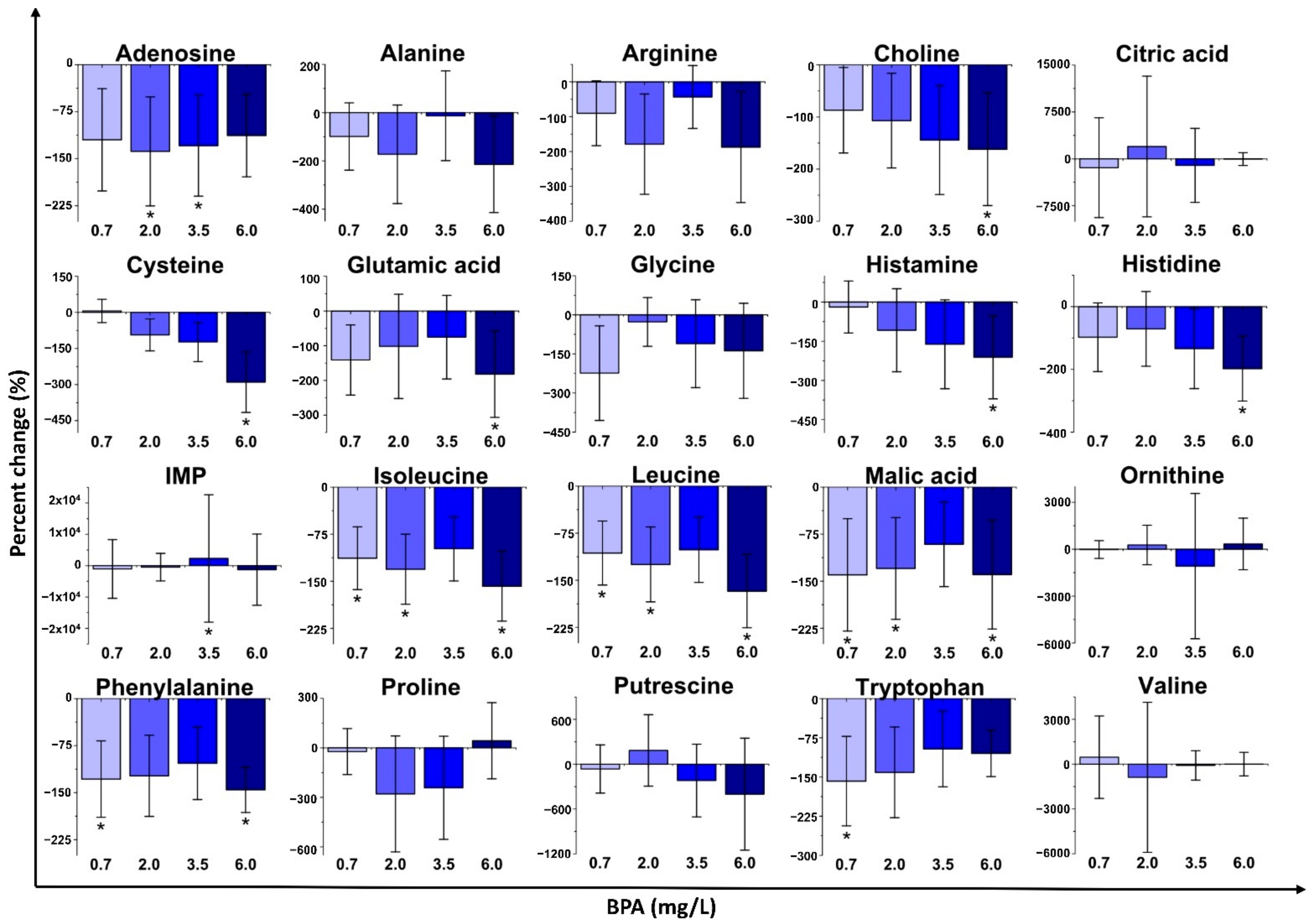
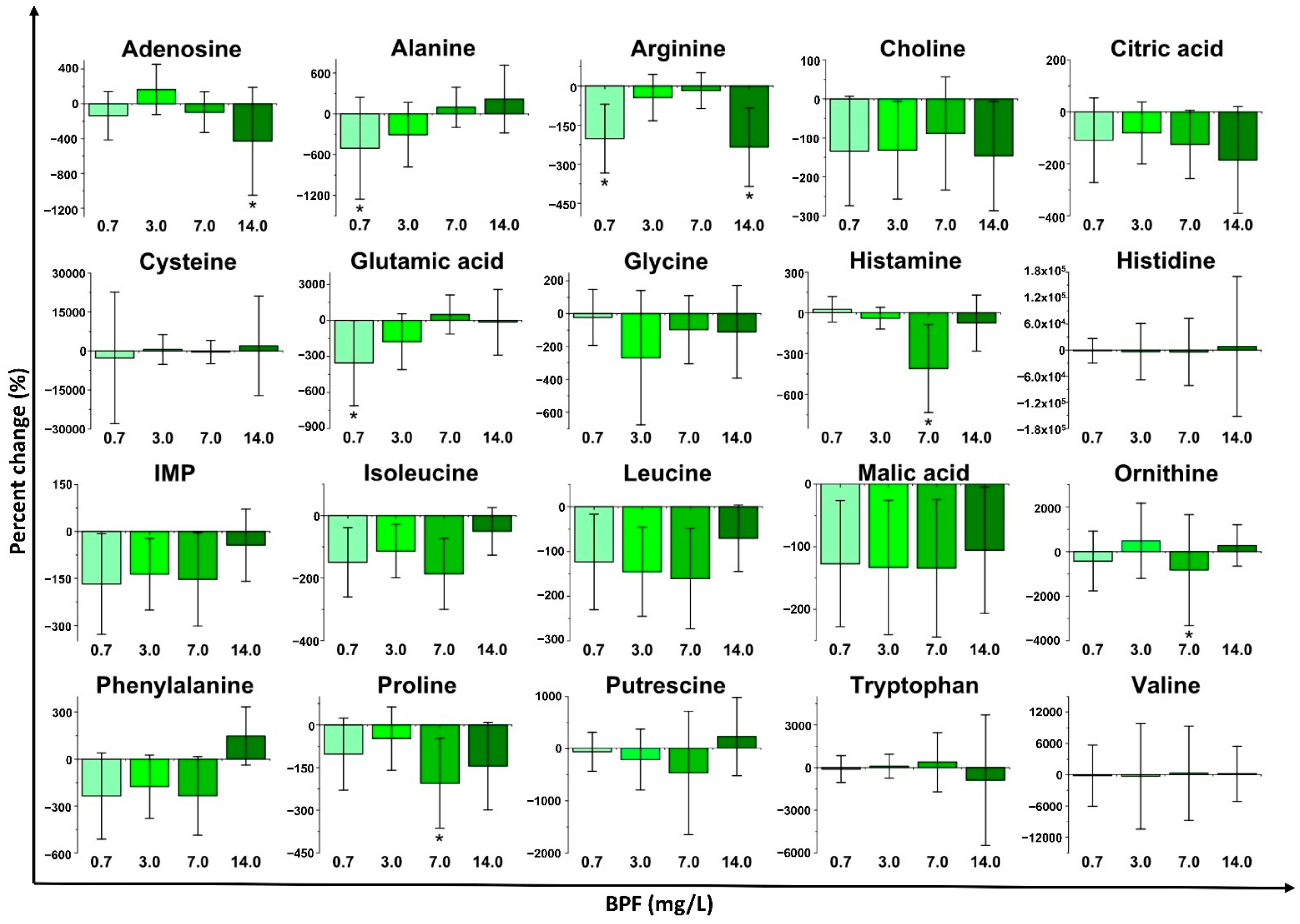
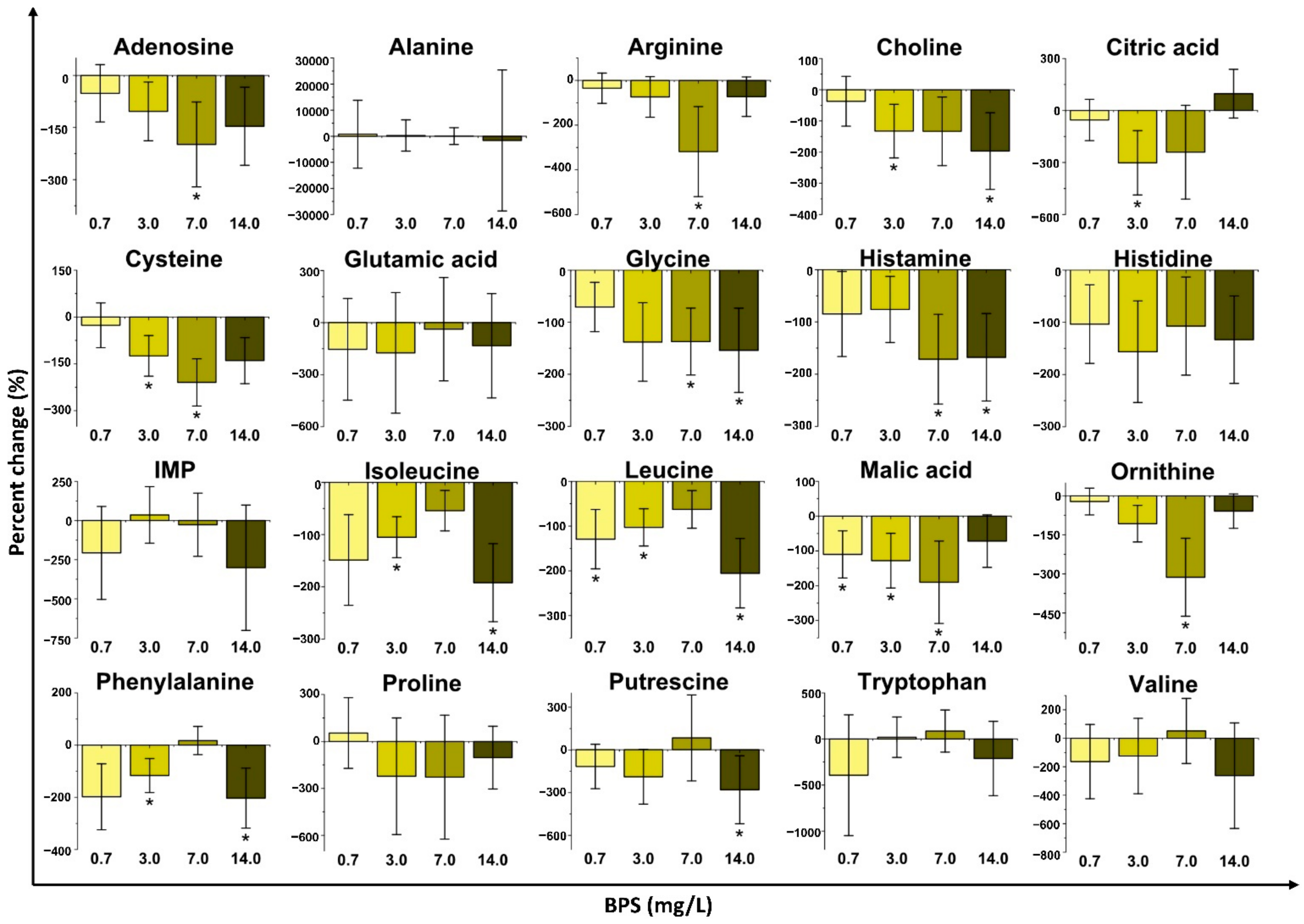
2.2. Changes in Metabolite Concentrations with Bisphenol Exposure
2.2.1. Bisphenol A (BPA) Exposure
2.2.2. Bisphenol F (BPF) Exposure
2.2.3. Bisphenol S (BPS) Exposure
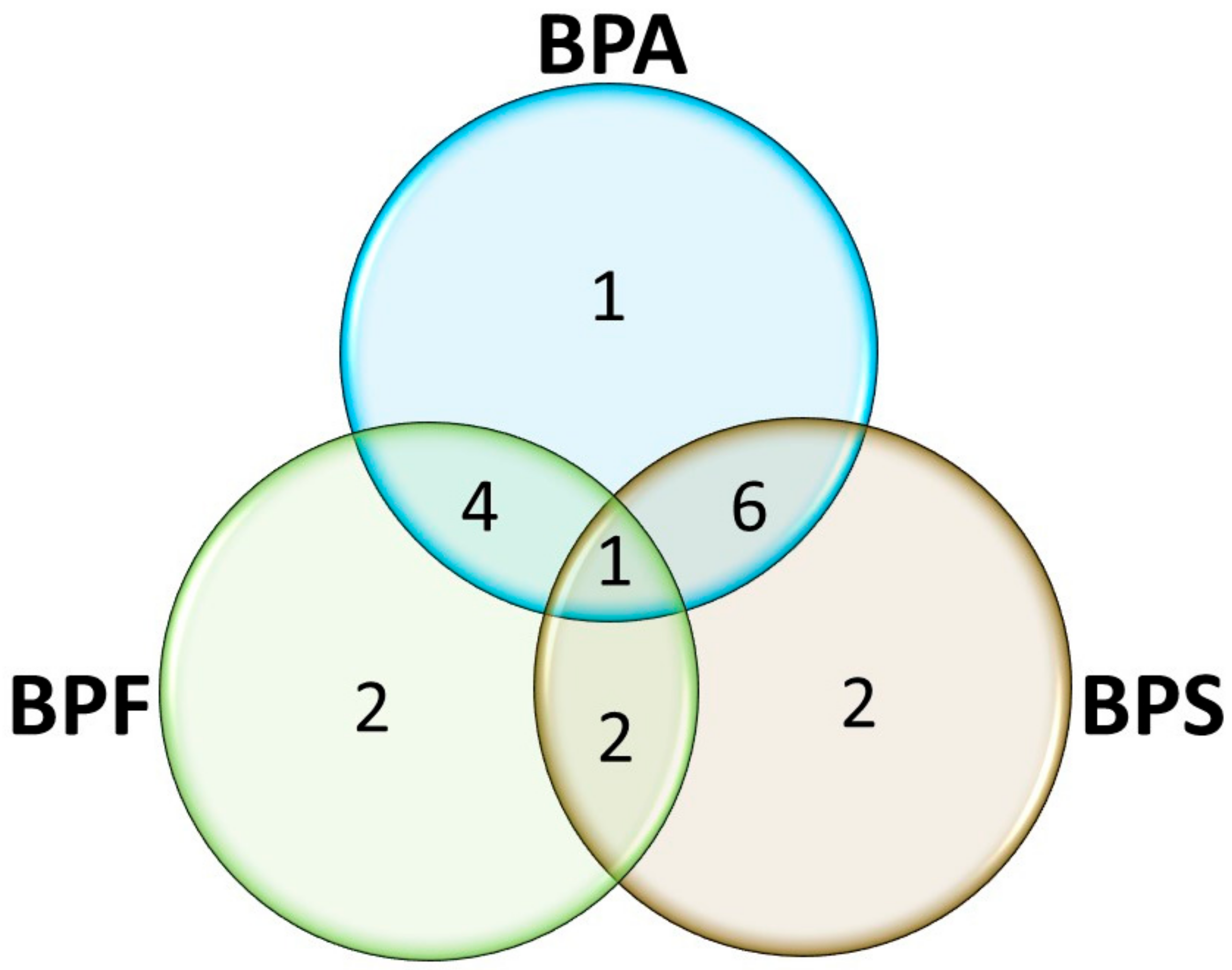
2.3. Pathway Analysis and Metabolic Networks
3. Discussion
4. Materials and Methods
4.1. Daphnia magna Culturing
4.2. Sub-Lethal Bisphenol Exposure
4.3. Metabolite Extraction and Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) Analysis
4.4. Data Processing and Pathway Interpretation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Staples, C.A.; Dome, P.B.; Klecka, G.M.; Oblock, S.T.; Harris, L.R. A review of the environmental fate, effects, and exposures of bisphenol A. Chemosphere 1998, 36, 2149–2173. [Google Scholar] [CrossRef]
- Im, J.; Löffler, F.E. Fate of bisphenol A in terrestrial and aquatic environments. Environ. Sci. Technol. 2016, 50, 8403–8416. [Google Scholar] [CrossRef]
- Vasiljevic, T.; Harner, T. Bisphenol A and its analogues in outdoor and indoor air: Properties, sources and global levels. Sci. Total Environ. 2021, 789, 148013. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Le, H.H.; Carlson, E.M.; Chua, J.P.; Belcher, S.M. Bisphenol A is released from polycarbonate drinking bottles and mimics the neurotoxic actions of estrogen in developing cerebellar neurons. Toxicol. Lett. 2008, 176, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Government of Canada. Proposed Risk Management Approach for Bisphenol A. 2018; pp. 1–19. Available online: http://www.ec.gc.ca/ese-ees/6FA54372-A09E-45CD-8A5F-39EBDD55D13A/batch2_80-05-7_rm_en.pdf (accessed on 23 July 2021).
- The European Commission. Regulation (EU) 2018/213 (on the use of bisphenol A in varnishes and coatings intended to come into contact with food). Off. J. Eur. Union 2018, 2018, 6–12. [Google Scholar]
- Yamazaki, E.; Yamashita, N.; Taniyasu, S.; Lam, J.; Lam, P.K.S.; Moon, H.B.; Jeong, Y.; Kannan, P.; Achyuthan, H.; Munuswamy, N.; et al. Bisphenol A and other bisphenol analogues including BPS and BPF in surface water samples from Japan, China, Korea and India. Ecotoxicol. Environ. Saf. 2015, 122, 565–572. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, L.; Lu, G.; Jiang, R.; Yan, Z.; Li, Y. Occurrence, toxicity and ecological risk of Bisphenol A analogues in aquatic environment—A review. Ecotoxicol. Environ. Saf. 2021, 208, 111481. [Google Scholar] [CrossRef]
- Chen, D.; Kannan, K.; Tan, H.; Zheng, Z.; Feng, Y.-L.; Wu, Y.; Widelka, M. Bisphenol analogues other than BPA: Environmental occurrence, human exposure, and toxicity—a review. Environ. Sci. Technol. 2016, 50, 5438–5453. [Google Scholar] [CrossRef]
- Usman, A.; Ahmad, M. From BPA to its analogues: Is it a safe journey? Chemosphere 2016, 158, 131–142. [Google Scholar] [CrossRef]
- Wang, Y.; Aimuzi, R.; Nian, M.; Zhang, Y.; Luo, K.; Zhang, J. Bisphenol A substitutes and sex hormones in children and adolescents. Chemosphere 2021, 278, 130396. [Google Scholar] [CrossRef]
- Cabaton, N.; Dumont, C.; Severin, I.; Perdu, E.; Zalko, D.; Cherkaoui-Malki, M.; Chagnon, M.C. Genotoxic and endocrine activities of bis(hydroxyphenyl)methane (bisphenol F) and its derivatives in the HepG2 cell line. Toxicology 2009, 255, 15–24. [Google Scholar] [CrossRef]
- Naderi, M.; Wong, M.Y.L.; Gholami, F. Developmental exposure of zebrafish (Danio rerio) to bisphenol-S impairs subsequent reproduction potential and hormonal balance in adults. Aquat. Toxicol. 2014, 148, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, F.; Moon, H.B.; Yamashita, N.; Yun, S.; Kannan, K. Bisphenol analogues in sediments from industrialized areas in the United States, Japan, and Korea: Spatial and temporal distributions. Environ. Sci. Technol. 2012, 46, 11558–11565. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Hu, A.; Li, Y.; He, Y.; Xu, J.; Lu, Z. Determination and occurrence of bisphenol A and thirteen structural analogs in soil. Chemosphere 2021, 277, 130232. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, F.; Alomirah, H.; Loi, V.D.; Mohd, M.A.; Moon, H.B.; Nakata, H.; Kannan, K. Bisphenol S in urine from the United States and seven Asian countries: Occurrence and human exposures. Environ. Sci. Technol. 2012, 46, 6860–6866. [Google Scholar] [CrossRef]
- Cao, P.; Zhong, H.N.; Qiu, K.; Li, D.; Wu, G.; Sui, H.X.; Song, Y. Exposure to bisphenol A and its substitutes, bisphenol F and bisphenol S from canned foods and beverages on Chinese market. Food Control 2021, 120, 107502. [Google Scholar] [CrossRef]
- Liao, C.; Kannan, K. Concentrations and profiles of bisphenol A and other bisphenol analogues in foodstuffs from the United States and their implications for human exposure. J. Agric. Food Chem. 2013, 61, 4655–4662. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, Y.; Li, Y.; Ashfaq, M.; Dai, L.; Xie, X.; Yu, C.-P. Fate and mass balance of bisphenol analogues in wastewater treatment plants in Xiamen City, China. Environ. Pollut. 2017, 225, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Kannan, K. Mass flows and removal of eight bisphenol analogs, bisphenol A diglycidyl ether and its derivatives in two wastewater treatment plants in New York State, USA. Sci. Total Environ. 2019, 648, 442–449. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Yan, K.; Wu, S.; Han, Z.; Guo, R.; Chen, M.; Yang, Q.; Zhang, S.; Chen, J. Bisphenol analogues in surface water and sediment from the shallow Chinese freshwater lakes: Occurrence, distribution, source apportionment, and ecological and human health risk. Chemosphere 2017, 184, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Ocharoen, Y.; Boonphakdee, C.; Boonphakdee, T.; Shinn, A.P.; Moonmangmee, S. High levels of the endocrine disruptors bisphenol-A and 17β-estradiol detected in populations of green mussel, Perna viridis, cultured in the Gulf of Thailand. Aquaculture 2018, 497, 348–356. [Google Scholar] [CrossRef]
- Corrales, J.; Kristofco, L.A.; Baylor Steele, W.; Yates, B.S.; Breed, C.S.; Spencer Williams, E.; Brooks, B.W. Global assessment of bisphenol a in the environment: Review and analysis of its occurrence and bioaccumulation. Dose-Response 2015, 13, 1559325815598308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žalmanová, T.; Hoško, K.; Nevoral, J.; Adámková, K.; Kott, T.; Šulc, M.; Kotíková, Z.; Prokešová, Š.; Jílek, F.; Králíčková, M.; et al. Bisphenol S negatively affects the meotic maturation of pig oocytes. Sci. Rep. 2017, 7, 485. [Google Scholar] [CrossRef] [PubMed]
- Rosenmai, A.K.; Dybdahl, M.; Pedersen, M.; van Vugt-Lussenburg, B.M.A.; Wedebye, E.B.; Taxvig, C.; Vinggaard, A.M. Are structural analogues to bisphenol a safe alternatives? Toxicol. Sci. 2014, 139, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Moreman, J.; Lee, O.; Trznadel, M.; David, A.; Kudoh, T.; Tyler, C.R. Acute toxicity, teratogenic, and estrogenic effects of bisphenol A and its alternative replacements bisphenol S, bisphenol F, and bisphenol AF in zebrafish embryo-larvae. Environ. Sci. Technol. 2017, 51, 12796–12805. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Lee, M.-C.; Yoon, D.-S.; Han, J.; Kim, M.; Hwang, U.-K.; Jung, J.-H.; Lee, J.-S. Effects of bisphenol A and its analogs bisphenol F and S on life parameters, antioxidant system, and response of defensome in the marine rotifer Brachionus koreanus. Aquat. Toxicol. 2018, 199, 21–29. [Google Scholar] [CrossRef]
- Chen, M.Y.; Ike, M.; Fujita, M. Acute toxicity, mutagenicity, and estrogenicity of bisphenol-A and other bisphenols. Environ. Toxicol. 2002, 17, 80–86. [Google Scholar] [CrossRef]
- In, S.; Yoon, H.W.; Yoo, J.W.; Cho, H.; Kim, R.O.; Lee, Y.M. Acute toxicity of bisphenol A and its structural analogues and transcriptional modulation of the ecdysone-mediated pathway in the brackish water flea Diaphanosoma celebensis. Ecotoxicol. Environ. Saf. 2019, 179, 310–317. [Google Scholar] [CrossRef]
- Lee, S.; Kim, C.; Shin, H.; Kho, Y.; Choi, K. Comparison of thyroid hormone disruption potentials by bisphenols A, S, F, and Z in embryo-larval zebrafish. Chemosphere 2019, 221, 115–123. [Google Scholar] [CrossRef]
- Tišler, T.; Krel, A.; Gerželj, U.; Erjavec, B.; Dolenc, M.S.; Pintar, A. Hazard identification and risk characterization of bisphenols A, F and AF to aquatic organisms. Environ. Pollut. 2016, 212, 472–479. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, Z.; Zhang, L.; Deng, Z.; Yuan, J.; Zhang, S.; Chen, J.; Guo, R. Food up-take and reproduction performance of Daphnia magna under the exposure of bisphenols. Ecotoxicol. Environ. Saf. 2019, 170, 47–54. [Google Scholar] [CrossRef]
- Gómez, C.; Gallart-Ayala, H. Metabolomics: A tool to characterize the effect of phthalates and bisphenol A. Environ. Rev. 2018, 26, 351–357. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Nagato, E.G.; Simpson, M.J. NMR-based metabolomics of Daphnia magna: Insights into aquatic ecosystem health. eMagRes 2017, 6, 315–324. [Google Scholar] [CrossRef]
- Størseth, T.R.; Hammer, K.M. Environmental metabolomics of aquatic organisms. In eMagRes; John Wiley & Sons, Ltd.: Chichester, UK, 2013; Volume 2, pp. 541–548. ISBN 9780470034590. [Google Scholar]
- Viant, M.R. Metabolomics of aquatic organisms: The new “omics” on the block. Mar. Ecol. Prog. Ser. 2007, 332, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Nagato, E.G.; Simpson, A.J.; Simpson, M.J. Metabolomics reveals energetic impairments in Daphnia magna exposed to diazinon, malathion and bisphenol-A. Aquat. Toxicol. 2016, 170, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.; Soong, R.; Bermel, W.; Ning, P.; Dutta Majumdar, R.; Tabatabaei-Anaraki, M.; Heumann, H.; Gundy, M.; Bönisch, H.; Liaghati Mobarhan, Y.; et al. Selective amino acid-only in vivo NMR: A powerful tool to follow stress processes. ACS Omega 2019, 4, 9017–9028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garreta-Lara, E.; Gómez-Canela, C.; Campos, B.; Barata, C.; Tauler, R.; Lacorte, S. Combined targeted/untargeted analytical and chemometric approaches in the characterization of Daphnia magna metabolomic changes under bisphenol A exposure. Microchem. J. 2021, 165, 106150. [Google Scholar] [CrossRef]
- Tkaczyk, A.; Bownik, A.; Dudka, J.; Kowal, K.; Ślaska, B. Daphnia magna model in the toxicity assessment of pharmaceuticals: A review. Sci. Total Environ. 2021, 763, 143038. [Google Scholar] [CrossRef] [PubMed]
- Environment Canada. Biological Test Method: Reference Method for Determining Acute Lethality of Effluents to Daphnia Magna; Environmental Technology Centre: Ottawa, ON, Canada, 2000; ISBN 0660616157.
- OECD. Test No. 202: Daphnia sp. Acute Immobilisation Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2004; ISBN 9789264069947. [Google Scholar]
- OECD. Test No. 211: Daphnia Magna Reproduction Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2012; ISBN 9789264185203. [Google Scholar]
- Edison, A.; Hall, R.; Junot, C.; Karp, P.; Kurland, I.; Mistrik, R.; Reed, L.; Saito, K.; Salek, R.; Steinbeck, C.; et al. The time is right to focus on model organism metabolomes. Metabolites 2016, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Vandenbrouck, T.; Jones, O.A.H.; Dom, N.; Griffin, J.L.; De Coen, W. Mixtures of similarly acting compounds in Daphnia magna: From gene to metabolite and beyond. Environ. Int. 2010, 36, 254–268. [Google Scholar] [CrossRef]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. Evaluation of Daphnia magna metabolic responses to organic contaminant exposure with and without dissolved organic matter using 1H nuclear magnetic resonance (NMR)-based metabolomics. Ecotoxicol. Environ. Saf. 2018, 164, 189–200. [Google Scholar] [CrossRef]
- Constantinou, J.K.; Southam, A.D.; Kvist, J.; Jones, M.R.; Viant, M.R.; Mirbahai, L. Characterisation of the dynamic nature of lipids throughout the lifespan of genetically identical female and male Daphnia magna. Sci. Rep. 2020, 10, 5576. [Google Scholar] [CrossRef]
- Labine, L.M.; Simpson, M.J. Targeted metabolomic assessment of the sub-lethal toxicity of halogenated acetic acids (HAAs) to Daphnia magna. Metabolites 2021, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.-Y.; Simpson, M.J. Reproduction stage differentiates the time-course regulation of metabolites in Daphnia magna. Environ. Sci. Technol. 2019, 53, 12764–12773. [Google Scholar] [CrossRef] [PubMed]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A systematic review and comparison of the hormonal activity of bisphenol A substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Peluso, M.E.M.; Munnia, A.; Ceppi, M. Bisphenol-A exposures and behavioural aberrations: Median and linear spline and meta-regression analyses of 12 toxicity studies in rodents. Toxicology 2014, 325, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.M.; Abril, N.; Morales-Prieto, N.; Monterde, J.G.; Lora, A.J.; Ayala, N.; Moyano, R. Evaluation of toxicological endpoints in female zebrafish after bisphenol A exposure. Food Chem. Toxicol. 2018, 112, 19–25. [Google Scholar] [CrossRef]
- Ibba, M.; Söll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef]
- Harper, A.E.; Miller, R.H.; Block, K.P. Branched-chain amino acid metabolism. Annu. Rev. Nutr. 1984, 4, 409–454. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Cross, P.J.; Dobson, R.C.J.; Adams, L.E.; Savka, M.A.; Hudson, A.O. A three-ring circus: Metabolism of the three proteogenic aromatic amino acids and their role in the health of plants and animals. Front. Mol. Biosci. 2018, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef]
- Steuer, R. On the analysis and interpretation of correlations in metabolomic data. Brief. Bioinform. 2006, 7, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Jin, J.; Kurobe, T.; Ramírez-Duarte, W.F.; Bolotaolo, M.B.; Lam, C.H.; Pandey, P.K.; Hung, T.-C.; Stillway, M.E.; Zweig, L.; Caudill, J.; et al. Sub-lethal effects of herbicides penoxsulam, imazamox, fluridone and glyphosate on Delta Smelt (Hypomesus transpacificus). Aquat. Toxicol. 2018, 197, 79–88. [Google Scholar] [CrossRef]
- Marlatt, V.L.; Leung, T.Y.G.; Calbick, S.; Metcalfe, C.; Kennedy, C. Sub-lethal effects of a neonicotinoid, clothianidin, on wild early life stage sockeye salmon (Oncorhynchus nerka). Aquat. Toxicol. 2019, 217, 105335. [Google Scholar] [CrossRef]
- Melvin, S.D.; Habener, L.J.; Leusch, F.D.L.; Carroll, A.R. 1H NMR-based metabolomics reveals sub-lethal toxicity of a mixture of diabetic and lipid-regulating pharmaceuticals on amphibian larvae. Aquat. Toxicol. 2017, 184, 123–132. [Google Scholar] [CrossRef]
- Bouchnak, R.; Steinberg, C.E.W. Algal diets and natural xenobiotics impact energy allocation in cladocerans. I. Daphnia magna. Limnologica 2013, 43, 434–440. [Google Scholar] [CrossRef]
- Sheppard, K.; Yuan, J.; Hohn, M.J.; Jester, B.; Devine, K.M.; Soll, D. From one amino acid to another: tRNA-dependent amino acid biosynthesis. Nucleic Acids Res. 2008, 36, 1813–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Lu, C.; Dong, X.; Zhang, Z.; Yang, M.; Xu, H. Proteomics analysis of zebrafish brain following chronically exposed to bisphenol A. Toxicol. Environ. Chem. 2017, 99, 469–481. [Google Scholar] [CrossRef]
- Yue, S.; Yu, J.; Kong, Y.; Chen, H.; Mao, M.; Ji, C.; Shao, S.; Zhu, J.; Gu, J.; Zhao, M. Metabolomic modulations of HepG2 cells exposed to bisphenol analogues. Environ. Int. 2019, 129, 59–67. [Google Scholar] [CrossRef]
- Sperber, S.; Wahl, M.; Berger, F.; Kamp, H.; Lemke, O.; Starck, V.; Walk, T.; Spitzer, M.; Ravenzwaay, B.V. Metabolomics as read-across tool: An example with 3-aminopropanol and 2-aminoethanol. Regul. Toxicol. Pharmacol. 2019, 108, 104442. [Google Scholar] [CrossRef]
- Yoshizawa, F. Regulation of protein synthesis by branched-chain amino acids in vivo. Biochem. Biophys. Res. Commun. 2004, 313, 417–422. [Google Scholar] [CrossRef]
- Yoon, C.; Yoon, D.; Cho, J.; Kim, S.; Lee, H.; Choi, H.; Kim, S. 1H-NMR-based metabolomic studies of bisphenol A in zebrafish (Danio rerio). J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2017, 52, 282–289. [Google Scholar] [CrossRef]
- Ji, C.; Wei, L.; Zhao, J.; Wu, H. Metabolomic analysis revealed that female mussel Mytilus galloprovincialis was sensitive to bisphenol A exposures. Environ. Toxicol. Pharmacol. 2014, 37, 844–849. [Google Scholar] [CrossRef]
- Zeng, J.; Kuang, H.; Hu, C.; Shi, X.; Yan, M.; Xu, L.; Wang, L.; Xu, C.; Xu, G. Effect of bisphenol A on rat metabolic profiling studied by using capillary electrophoresis time-of-flight mass spectrometry. Environ. Sci. Technol. 2013, 47, 7457–7465. [Google Scholar] [CrossRef]
- Mao, L.; Fang, S.; Zhao, M.; Liu, W.; Jin, H. Effects of bisphenol A and bisphenol S exposure at low doses on the metabolome of adolescent male Sprague–Dawley rats. Chem. Res. Toxicol. 2021, 34, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Reddivari, L.; Veeramachaneni, D.N.R.; Walters, W.A.; Lozupone, C.; Palmer, J.; Hewage, M.K.K.; Bhatnagar, R.; Amir, A.; Kennett, M.J.; Knight, R.; et al. Perinatal bisphenol A exposure induces chronic inflammation in rabbit offspring via modulation of gut bacteria and their metabolites. mSystems 2017, 2, e00093-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.M. Arginine metabolism: Boundaries of our knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.-Z.; Lin, X.-M.; Cheng, Z.-X.; Su, Y.-B.; Li, W.-X.; Ali, F.-M.; Zheng, J.; Peng, B. Identification and efficacy of glycine, serine and threonine metabolism in potentiating kanamycin-mediated killing of Edwardsiella piscicida. J. Proteom. 2018, 183, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Ontario Ministry of the Environment. Daphnia Magna Culturing; Ontario Ministry of the Environment: Toronto, ON, Canada, 2012; Volume SOP DM1.v8, pp. 1–10. [Google Scholar]
- Ontario Ministry of the Environment. Algae Culturing for Use as Food; Ontario Ministry of the Environment: Toronto, ON, Canada, 2012; Volume SOP AL1.v6, pp. 1–10. [Google Scholar]
- Rezaee, M.; Yamini, Y.; Shariati, S.; Esrafili, A.; Shamsipur, M. Dispersive liquid–liquid microextraction combined with high-performance liquid chromatography-UV detection as a very simple, rapid and sensitive method for the determination of bisphenol A in water samples. J. Chromatogr. A 2009, 1216, 1511–1514. [Google Scholar] [CrossRef]
- Coelho, C.M.; de Andrade, J.R.; da Silva, M.G.C.; Vieira, M.G.A. Removal of propranolol hydrochloride by batch biosorption using remaining biomass of alginate extraction from Sargassum filipendula algae. Environ. Sci. Pollut. Res. 2020, 27, 16599–16611. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef]
- Tamone, S.L.; Harrison, J.F. Linking insects with crustacea: Physiology of the pancrustacea: An introduction to the symposium. Integr. Comp. Biol. 2015, 55, 765–770. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways | Compounds | Metabolites Associated |
| Aminoacyl-tRNA biosynthesis | BPA, BPF, and BPS | Alanine, Arginine, Cysteine, Glutamic acid, Glycine, Histidine, Isoleucine, Leucine, Phenylalanine, Tryptophan |
| Valine, leucine, and isoleucine biosynthesis | BPA and BPS | Leucine, Isoleucine |
| Valine, leucine, and isoleucine degradation | BPA and BPS | Leucine, Isoleucine |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | BPA and BPS | Phenylalanine |
| Glyoxylate and dicarboxylate metabolism | BPA and BPS | Citric acid, Malic acid, Glutamic acid |
| Glutathione metabolism | BPA and BPS | Cysteine, Glutamic acid, Glycine, Ornithine, Putrescine |
| Glycine, serine, and threonine metabolism | BPA and BPS | Choline, Cysteine, Glycine |
| Purine metabolism | BPA and BPF | Adenosine, Inosine monophosphate |
| Histidine metabolism | BPA and BPF | Histidine, Histamine |
| Nitrogen metabolism | BPA and BPF | Glutamic acid |
| D-Glutamine and D-glutamate metabolism | BPA and BPF | Glutamic acid |
| Arginine biosynthesis | BPF and BPS | Arginine, Glutamic acid, Ornithine |
| Arginine and proline metabolism | BPF and BPS | Alanine, Arginine, Glutamic acid, Proline, Ornithine |
| Phenylalanine metabolism | BPA | Phenylalanine |
| Alanine, aspartate, and glutamate metabolism | BPF | Alanine, Glutamic acid |
| Butanoate metabolism | BPF | Glutamic acid |
| Citrate cycle (TCA cycle) | BPS | Citric acid, Malic acid |
| Pyruvate metabolism | BPS | Malic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira Pereira, E.A.; Labine, L.M.; Kleywegt, S.; Jobst, K.J.; Simpson, A.J.; Simpson, M.J. Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna. Metabolites 2021, 11, 666. https://doi.org/10.3390/metabo11100666
Oliveira Pereira EA, Labine LM, Kleywegt S, Jobst KJ, Simpson AJ, Simpson MJ. Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna. Metabolites. 2021; 11(10):666. https://doi.org/10.3390/metabo11100666
Chicago/Turabian StyleOliveira Pereira, Erico A., Lisa M. Labine, Sonya Kleywegt, Karl J. Jobst, André J. Simpson, and Myrna J. Simpson. 2021. "Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna" Metabolites 11, no. 10: 666. https://doi.org/10.3390/metabo11100666
APA StyleOliveira Pereira, E. A., Labine, L. M., Kleywegt, S., Jobst, K. J., Simpson, A. J., & Simpson, M. J. (2021). Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna. Metabolites, 11(10), 666. https://doi.org/10.3390/metabo11100666