
Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Effects of Long-Term Nitrogen Application on Tea Yield
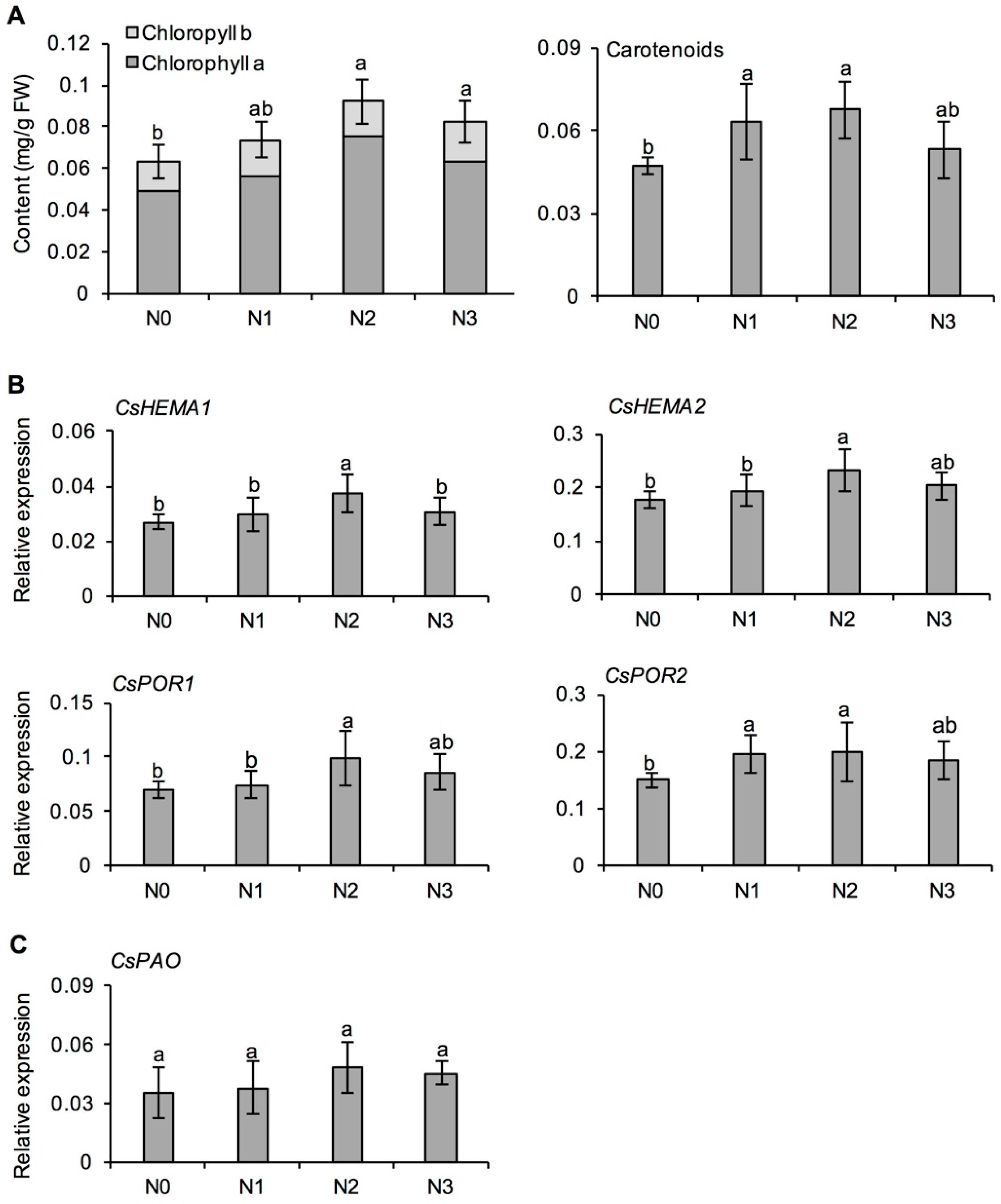
2.2. Effects of Long-Term Nitrogen Application on Chlorophyll Formation in Tea Leaves
2.3. Effects of Long-Term Nitrogen Application on the Formation of Free Amino Acids in Tea Leaves
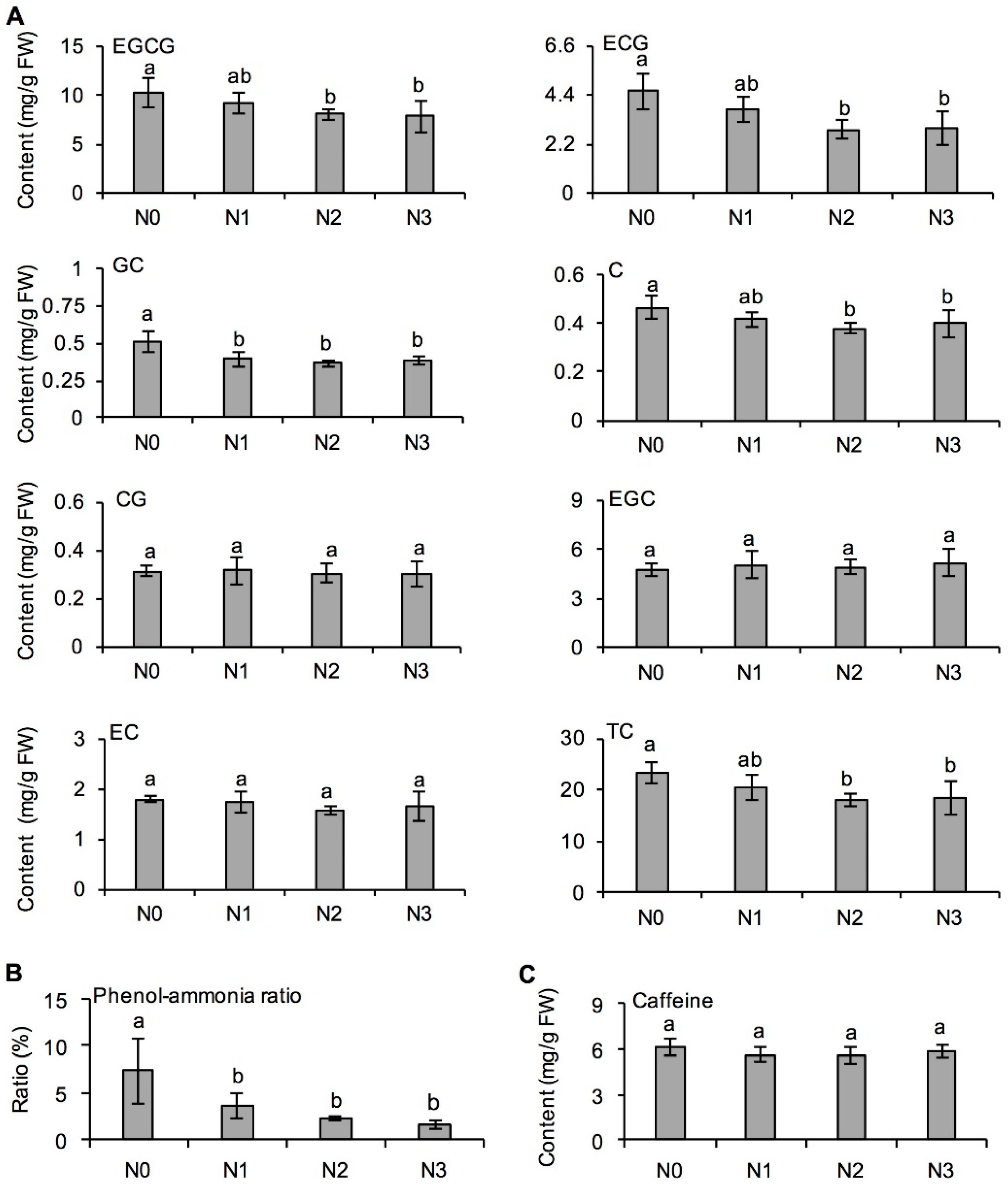
2.4. Effects of Long-Term Nitrogen Application on Catechin and Caffeine Formation in Fresh Tea Leaves
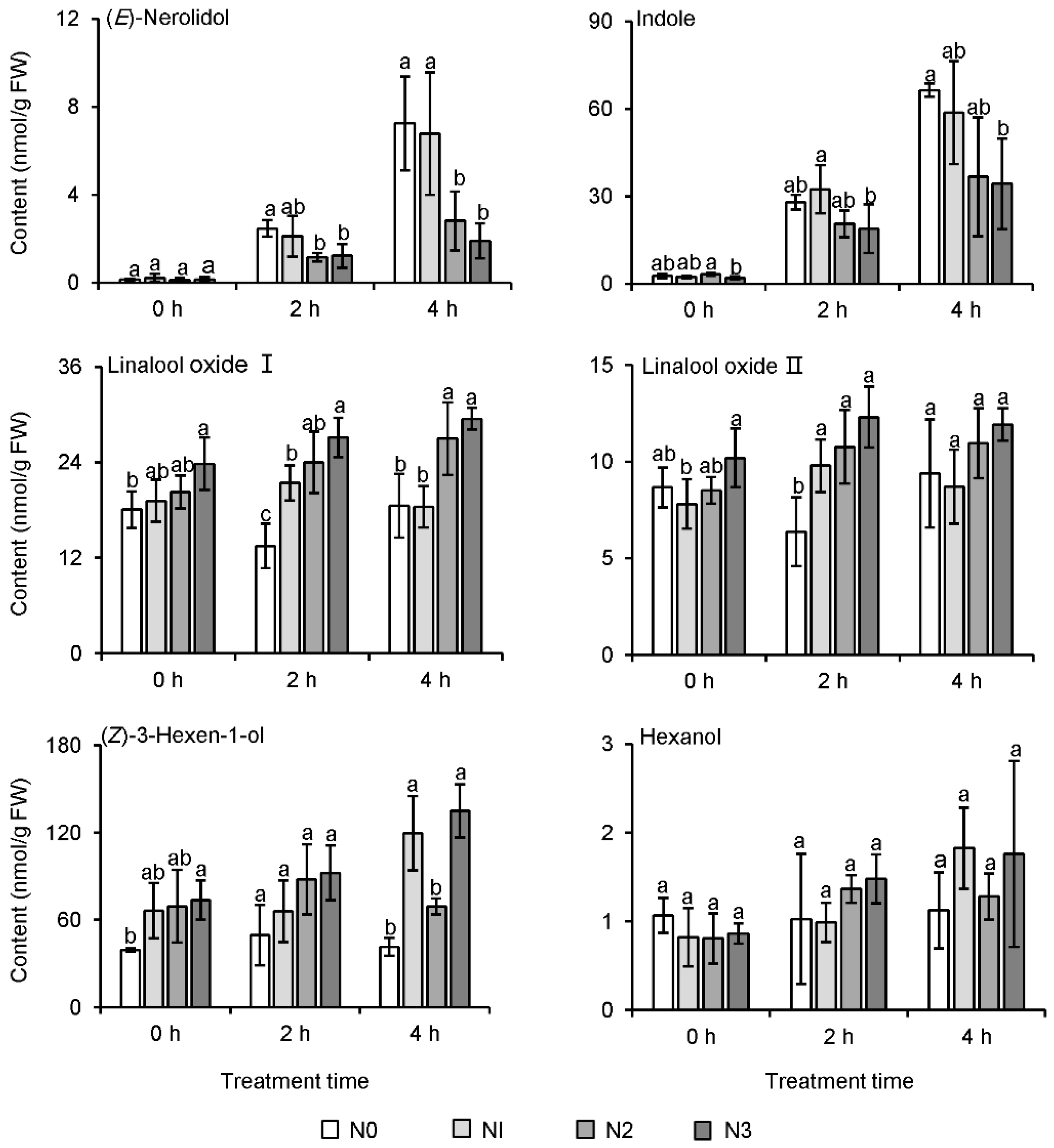
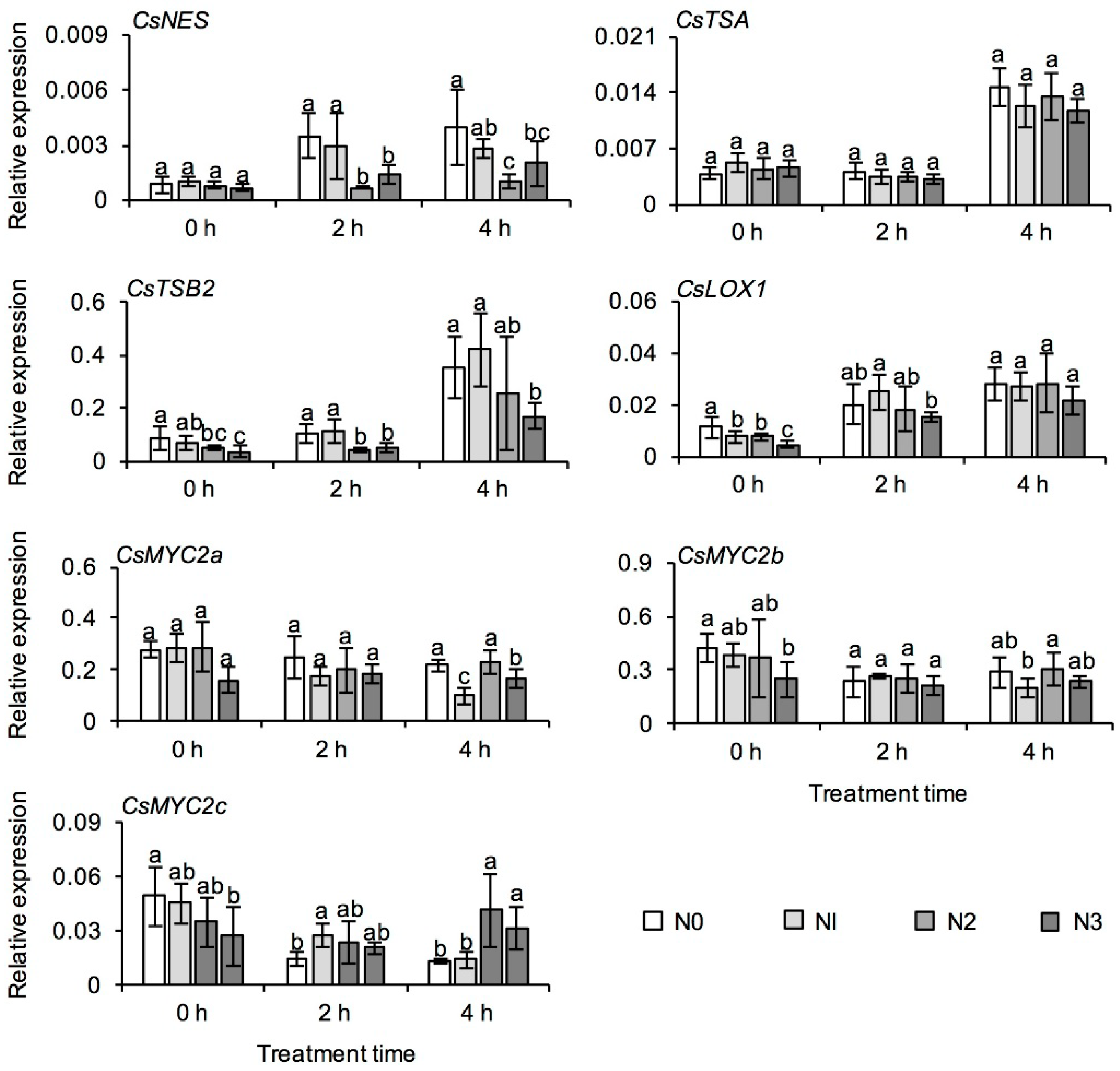
2.5. Effect of Long-Term Nitrogen Application on the Formation of Aroma Compounds in Tea Leaves
3. Discussion
3.1. Effect of Long-Term Nitrogen Application on Chlorophyll Synthesis in Fresh Tea Leaves
3.2. Effect of Long-Term Nitrogen Application in Promoting Amino Acid Synthesis
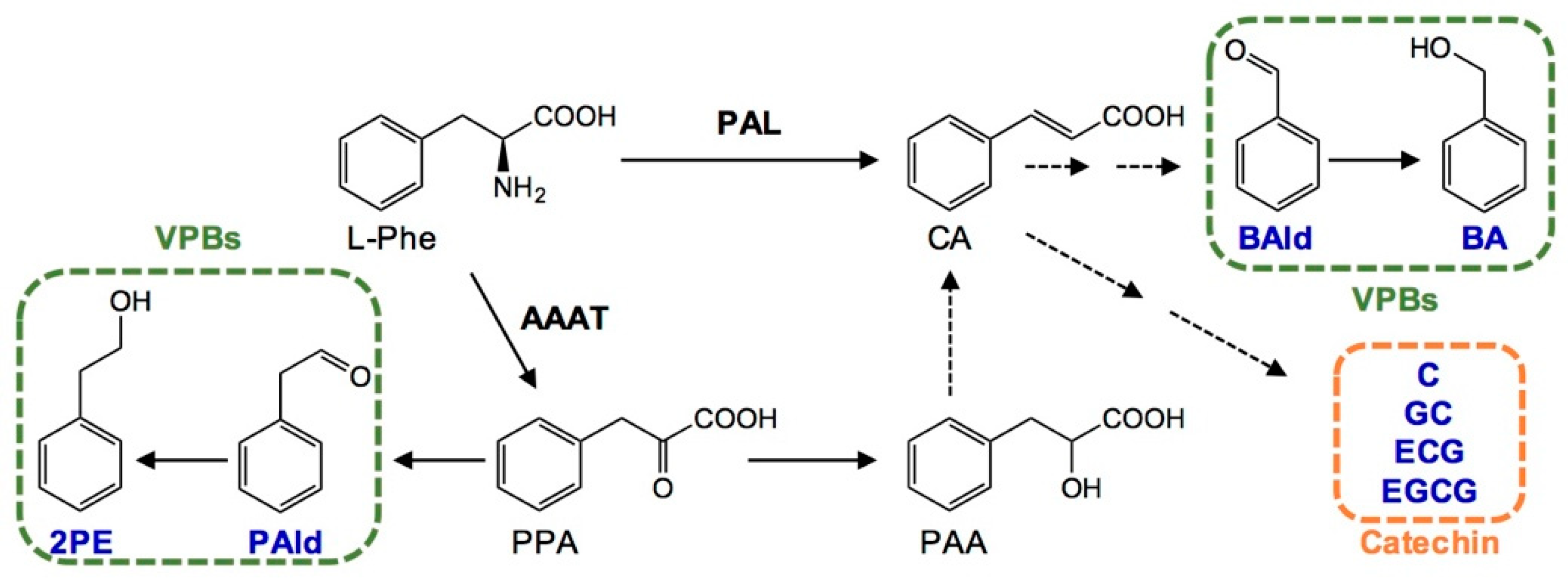
3.3. Effect of Long-Term Nitrogen Application in Decreasing Metabolites Derived from l-Phenylpropanoid Metabolism
3.4. Long-Term Nitrogen Application Might Affect the Formation of Floral Aroma Compounds in Tea Leaves during the Withering Process
4. Materials and Methods
4.1. Experimental Design and Different Fertilization Treatments
4.2. Tea Leaves Sampling and Analysis
4.3. Determination of Chlorophyll and Carotenoids
4.4. Analyses of Free Amino Acids in Tea Samples
4.5. Extraction and Analyses of Caffeine, Catechins, and Total Polyphenols in Tea Leaves
4.6. Extraction and Analysis of Aroma Compounds in Tea Leaves
4.7. Transcript Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EGCG | Epigallocatechin gallate |
| ECG | Epicatechin gallate |
| GC | Gallocatechin |
| C | Catechin |
| EC | Epicatechin |
| EGC | Epigallocatechin |
| CG | Catechin gallate |
| TC | Total catechin |
| PAL | Phenylalanine lyase |
| AAAT | Aromatic amino acid aminotransferase |
| NES | (E)-Nerolidol synthase |
| PCR | Polymerase chain reaction |
| EF1-α | Encoding elongation factor 1-α |
| HEMA | Glutamyl-tRNA reductase |
| POR | Protochlorophyllide oxidoreductase |
| PAO | Pheophorbide a oxygenase |
| TS | L-Theanine synthase |
| GS | L-Glutamine synthase |
| PDX | Pyridoxal 5′-phosphate synthase subunit |
| MYC | Myelocytomatosis proteins |
| TSA | Tryptophan synthase α-subunit |
| TSB | Tryptophan synthase β-subunit |
| LOX | Lipoxygenase |
| L-Phe | L-Phenylalanine |
| BAld | Benzaldehyde |
| BA | Benzyl alcohol |
| CA | trans-Cinnamic acid |
| PPA | Phenylpyruvic acid |
| PAA | Phenyllactic acid |
| PAld | Phenylacetaldehyde |
| 2-PE | 2-Phenylethanol |
| VPBs | Volatile phenylpropanoids/benzenoids |
| GC–MS | Gas chromatography–mass spectrometry |
| UPLC–QTOF–MS | Ultra performance liquid chromatography/quadrupole time-of-flight mass spectrometry |
References
- Tang, G.Y.; Meng, X.; Gan, R.Y.; Zhao, C.N.; Liu, Q.; Feng, Y.B.; Li, S.; Wei, X.L.; Atanasov, A.G.; Corke, H.; et al. Health functions and related molecular mechanisms of tea components: An update review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Jin, J.J.; Sun, C.J.; Ye, D.P.; Liu, Y.F. Simultaneous determination of six main types of lipid-soluble pigments in green tea by visible and near-infrared spectroscopy. Food Chem. 2019, 270, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Yang, C.Y.; Lee, S.J.; Wu, C.C.; Tzen, J.T.C. Catechin content and the degree of its galloylation in oolong tea are inversely correlated with cultivation altitude. J. Food Drug. Anal. 2014, 22, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.Y.; Zhao, C.N.; Xu, X.Y.; Gan, R.Y.; Cao, S.Y.; Liu, Q.; Shang, A.; Mao, Q.Q.; Li, H.B. Phytochemical composition and antioxidant capacity of 30 Chinese teas. Antioxidants 2019, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.Y.; Li, J.X.; Chen, W.; Wang, W.W.; Qi, D.D.; Pang, S.; Miao, A.Q. Study of the aroma formation and transformation during the manufacturing process of oolong tea by solid-phase micro-extraction and gas chromatography-mass spectrometry combined with chemometrics. Food Res. Int. 2018, 108, 413–422. [Google Scholar] [CrossRef]
- Zeng, L.T.; Watanabe, N.; Yang, Z.Y. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea (Camellia sinensis) to safely and effectively improve tea aroma. Crit. Rev. Food Sci. Nutr. 2019, 59, 2321–2334. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, X.C.; Su, X.G.; Yang, Z.Y. Chinese oolong tea: An aromatic beverage produced under multiple stresses. Trends Food Sci. Technol. 2020, 106, 242–253. [Google Scholar] [CrossRef]
- Deng, H.L.; Chen, S.S.; Zhou, Z.W.; Li, X.L.; Chen, S.; Hu, J.; Lai, Z.X.; Sun, Y. Transcriptome analysis reveals the effect of short-term sunlight on aroma metabolism in postharvest leaves of oolong tea (Camellia sinensis). Food Res. Int. 2020, 137, 109347. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.Y.; Zhou, J.; He, W.; Zeng, J.M.; Jiang, Y.W.; Cheng, H. Catechin contents in tea (Camellia sinensis) as affected by cultivar and environment and their relation to chlorophyll contents. Food Chem. 2011, 125, 44–48. [Google Scholar] [CrossRef]
- Yu, X.M.; Xiao, J.J.; Chen, S.; Yu, Y.; Ma, J.Q.; Lin, Y.Z.; Li, R.Z.; Lin, J.; Fu, Z.J.; Zhou, Q.Q.; et al. Metabolite signatures of diverse Camellia sinensis tea populations. Nat. Commun. 2020, 11, 5586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Wang, L.J.; Wei, K.; Ruan, L.; Wu, L.Y.; He, M.D.; Tong, H.R.; Cheng, H. Differential regulatory mechanisms of secondary metabolites revealed at different leaf positions in two related tea cultivars. Sci. Hortic. 2020, 272. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of root zone pH and form and concentration of nitrogen on accumulation of quality-related components in green tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Yang, T.Y.; Li, H.P.; Tai, Y.L.; Dong, C.X.; Cheng, X.M.; Xia, E.; Chen, Z.P.; Li, F.; Wan, X.C.; Zhang, Z.L. Transcriptional regulation of amino acid metabolism in response to nitrogen deficiency and nitrogen forms in tea plant root (Camellia sinensis L.). Sci. Rep. 2020, 10, 6868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, K.; Liao, W.Y.; Yi, X.Y.; Niu, S.Y.; Ma, L.F.; Shi, Y.Z.; Zhang, Q.F.; Liu, M.Y.; Ruan, J.Y. Fertilization status and reduction potential in tea gardens of China. J. Plant Nutr. Sci. 2019, 25, 421–432. [Google Scholar] [CrossRef]
- Wu, Y.Z.; Li, Y.; Fu, X.Q.; Shen, J.L.; Chen, D.; Wang, Y.; Liu, X.L.; Xiao, R.L.; Wei, W.X.; Wu, J.S. Effect of controlled-release fertilizer on N2O emissions and tea yield from a tea field in subtropical central China. Environ. Sci. Pollut. Res. Int. 2018, 25, 25580–25590. [Google Scholar] [CrossRef]
- Xie, S.W.; Yang, F.; Feng, H.X.; Yu, Z.Z.; Liu, C.S.; Wei, C.Y.; Liang, T. Organic fertilizer reduced carbon and nitrogen in runoff and buffered soil acidification in tea plantations: Evidence in nutrient contents and isotope fractionations. Sci. Total Environ. 2020. [Google Scholar] [CrossRef]
- Wang, Z.T.; Geng, Y.B.; Liang, T. Optimization of reduced chemical fertilizer use in tea gardens based on the assessment of related environmental and economic benefits. Sci. Total Environ. 2020, 713, 136439. [Google Scholar] [CrossRef]
- Liu, M.Y.; Burgos, A.; Ma, L.F.; Zhang, Q.F.; Tang, D.D.; Ruan, J.Y. Lipidomics analysis unravels the effect of nitrogen fertilization on lipid metabolism in tea plant (Camellia sinensis L.). BMC Plant Biol. 2017, 17, 165. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Hu, J.H.; Shi, Y.Z.; Liu, M.Y.; Zhang, Q.F.; Ruan, J.Y. Effects of nitrogen supply on flavonol glycoside biosynthesis and accumulation in tea leaves (Camellia sinensis). Plant Physiol. Biochem. 2019, 138, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Haerdter, R.; Gerendas, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.W.; Li, H.; Liu, J.X.; Wang, Y.; Zhuang, J. Integrative transcriptome, proteome, and microRNA analysis reveals the effects of nitrogen sufficiency and deficiency conditions on theanine metabolism in the tea plant (Camellia sinensis). Hortic. Res. 2020, 7, 65. [Google Scholar] [CrossRef]
- Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Nitrogen absorption by field-grown tea plants (Camellia sinensis) in winter dormancy and utilization in spring shoots. Plant Soil 2019, 442, 127–140. [Google Scholar] [CrossRef]
- Begum, Y.; Mondal, S.K. Comprehensive study of the genes involved in chlorophyll synthesis and degradation pathways in some monocot and dicot plant species. J. Biomol. Struct. Dyn. 2020, 3, 1–28. [Google Scholar] [CrossRef]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef]
- Fu, X.M.; Cheng, S.H.; Liao, Y.Y.; Xu, X.L.; Wang, X.C.; Hao, X.Y.; Xu, P.; Dong, F.; Yang, Z. Characterization of l-theanine hydrolase in vitro and subcellular distribution of its specific product ethylamine in tea (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 10842–10851. [Google Scholar] [CrossRef]
- Wei, C.L.; Yang, H.; Wang, S.B.; Zhao, J.; Liu, C.; Gao, L.P.; Xia, E.H.; Lu, Y.; Tai, Y.L.; She, G.B.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.T.; Yu, X.L.; He, C.; Qiu, A.D.; Li, Y.C.; Shu, Q.N.; Chen, Y.Q.; Ni, D.J. Withering degree affects flavor and biological activity of black tea: A non-targeted metabolomics approach. LWT Food Sci. Technol. 2020, 130, 109535. [Google Scholar] [CrossRef]
- Wang, X.Q.; Zeng, L.T.; Liao, Y.Y.; Zhou, Y.; Xu, X.L.; Dong, F.; Yang, Z.Y. An alternative pathway for the formation of aromatic aroma compounds derived from L-phenylalanine via phenylpyruvic acid in tea (Camellia sinensis (L.) O. Kuntze) leaves. Food Chem. 2019, 270, 17–24. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Fu, X.M.; Mei, X.; Cheng, S.H.; Gui, J.D.; Dong, F.; Tang, J.C.; Ma, S.Z.; Yang, Z.Y. Does oolong tea (Camellia sinensis) made from a combination of leaf and stem smell more aromatic than leaf-only tea? Contribution of the stem to oolong tea aroma. Food Chem. 2017, 237, 488–498. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Gui, J.D.; Fu, X.M.; Mei, X.; Zhen, Y.P.; Ye, T.X.; Du, B.; Dong, F.; Watanabe, N.; et al. Formation of volatile tea constituent indole during the oolong tea manufacturing process. J. Agric. Food Chem. 2016, 64, 5011–5019. [Google Scholar] [CrossRef]
- Wang, K.B.; Ruan, J.Y. Analysis of chemical components in green tea in relation with perceived quality, a case study with Longjing teas. Int. J. Food Sci. Technol. 2009, 44, 2476–2484. [Google Scholar] [CrossRef]
- Chen, P.A.; Lin, S.Y.; Liu, C.F.; Su, Y.S.; Cheng, H.Y.; Shiau, J.H.; Chen, I.Z. Correlation between nitrogen application to tea flushes and quality of green and black teas. Sci. Hortic. 2015, 181, 102–107. [Google Scholar] [CrossRef]
- Han, W.Y.; Ma, L.F.; Shi, Y.Z.; Ruan, J.Y.; Kemmitt, S.J. Nitrogen release dynamics and transformation of slow release fertiliser products and their effects on tea yield and quality. J. Sci. Food Agric. 2008, 88, 839–846. [Google Scholar] [CrossRef]
- Qiao, C.L.; Xu, B.; Han, Y.T.; Wang, J.; Wang, X.; Liu, L.L.; Liu, W.X.; Wan, S.Q.; Tan, H.; Liu, Y.Z.; et al. Synthetic nitrogen fertilizers alter the soil chemistry, production and quality of tea. A meta-analysis. Agron. Sustain. Dev. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.J.; Liao, W.Y.; Ding, Y.; Wang, H.S.; Xia, X.J. Effects of nitrogen fertilization on yield and quality of tea. J. Plant Nutr. Fertil. Sci. 2011, 16, 1430–1436. [Google Scholar] [CrossRef]
- Wang, H.J.; Hua, J.J.; Jiang, Y.W.; Yang, Y.Q.; Wang, J.J.; Yuan, H.B. Influence of fixation methods on the chestnut-like aroma of green tea and dynamics of key aroma substances. Food Res. Int. 2020, 136, 109479. [Google Scholar] [CrossRef]
- Brouwer, B.; Gardestrom, P.; Keech, O. In response to partial plant shading, the lack of phytochrome A does not directly induce leaf senescence but alters the fine-tuning of chlorophyll biosynthesis. J. Exp. Bot. 2014, 65, 4037–4049. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.G.; Xu, H.; Zhang, J.Y.; Liang, G.W.; Liu, Y.T.; Guo, A.G. Effect of low temperature on chlorophyll biosynthesis in albinism line of wheat (Triticum aestivum) FA85. Physiol Plant. 2012, 145, 384–394. [Google Scholar] [CrossRef]
- Petroutsos, D.; Tokutsu, R.; Maruyama, S.; Flori, S.; Greiner, A.; Magneschi, L.; Cusant, L.; Kottke, T.; Mittag, M.; Hegemann, P.; et al. A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 2016, 537, 563–566. [Google Scholar] [CrossRef]
- Kuai, B.K.; Chen, J.Y.; Hortensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J. Exp. Bot. 2018, 69, 751–767. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.L.; Chen, C.S.; Yue, C.; Hao, X.Y.; Yang, Y.J.; Wang, X.C. Complementary transcriptomic and proteomic analyses of a chlorophyll-deficient tea plant cultivar reveal multiple metabolic pathway changes. J. Proteom. 2016, 130, 160–169. [Google Scholar] [CrossRef]
- Hao, X.Y.; Zhang, W.F.; Liu, Y.; Zhang, H.J.; Ren, H.Z.; Chen, Y.; Wang, L.; Zeng, J.M.; Yang, Y.J.; Wang, X.C. Pale green mutant analyses reveal the importance of CsGLKs in chloroplast developmental regulation and their effects on flavonoid biosynthesis in tea plant. Plant Physiol. Biochem. 2020, 146, 392–402. [Google Scholar] [CrossRef]
- Wen, B.B.; Li, C.; Fu, X.L.; Li, D.M.; Li, L.; Chen, X.D.; Wu, H.Y.; Cui, X.W.; Zhang, X.H.; Shen, H.Y.; et al. Effects of nitrate deficiency on nitrate assimilation and chlorophyll synthesis of detached apple leaves. Plant Physiol. Biochem. 2019, 142, 363–371. [Google Scholar] [CrossRef]
- Bao, A.; Zhao, Z.Q.; Ding, G.D.; Shi, L.; Xu, F.S.; Cai, H.M. The stable level of Glutamine synthetase 2 plays an important role in rice growth and in carbon-nitrogen metabolic balance. Int. J. Mol. Sci. 2015, 16, 12713–12736. [Google Scholar] [CrossRef] [Green Version]
- Tegeder, M.; Hammes, U.Z. The way out and in: Phloem loading and unloading of amino acids. Curr. Opin. Plant Biol. 2018, 43, 16–21. [Google Scholar] [CrossRef]
- Ratiu, I.A.; Al-Suod, H.; Ligor, M.; Monedeiro, F.; Buszewski, B. Effects of growth conditions and cultivability on the content of cyclitols in Medicago sativa. Int. J. Environ. Sci. Technol. 2021, 18, 33–48. [Google Scholar] [CrossRef]
- Ruan, L.; Wei, K.; Wang, L.Y.; Cheng, H.; Wu, L.Y.; Li, H.L. Characteristics of free amino acids (the quality chemical components of tea) under spatial heterogeneity of different nitrogen forms in tea (Camellia sinensis) plants. Molecules 2019, 24, 415. [Google Scholar] [CrossRef] [Green Version]
- Sawicka, B.; Pszczolkowski, P.; Krochmal-Marczak, B.; Barbas, P.; Ozdemir, F.A. The effects of variable nitrogen fertilization on amino acid content in sweet potato tubers (Ipomoea batatas L. [Lam.]) cultivated in central and eastern Europe. J. Sci. Food Agric. 2020, 100, 4132–4138. [Google Scholar] [CrossRef]
- Fu, X.M.; Liao, Y.Y.; Cheng, S.H.; Xu, X.L.; Grierson, D.; Yang, Z.Y. Nonaqueous fractionation and overexpression of fluorescent-tagged enzymes reveals the subcellular sites of l-theanine biosynthesis in tea. Plant Biotechnol. J. 2021, 19, 98–108. [Google Scholar] [CrossRef]
- Tang, D.; Liu, M.Y.; Zhang, Q.F.; Fan, K.; Ruan, J.Y. Isolation and characterization of chloroplastic glutamine synthetase gene (CsGS2) in tea plant Camellia sinensis. Plant Physiol. Biochem. 2020, 155, 321–329. [Google Scholar] [CrossRef]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.L.; Chen, L.B.; Wu, Q.; Liu, Y.L.; Yu, J.; Li, F.D.; Li, R.P.; et al. The Reference genome of tea plant and resequencing of 81 diverse accessions provide insights into its genome evolution and adaptation. Mol. Plant. 2020, 13, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Li, W.; Li, K.; Nan, H.; Shi, C.; Zhang, Y.; Dai, Z.Y.; Lin, Y.L.; Yang, X.L.; Tong, Y.; et al. The chromosome-level reference genome of tea tree unveils recent bursts of non-autonomous LTR retrotransposons in driving genome size evolution. Mol. Plant 2020, 13, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Feng, H.; Chang, Y.X.; Ma, C.L.; Wang, L.Y.; Hao, X.Y.; Li, A.L.; Cheng, H.; Wang, L.; Cui, P.; et al. Population sequencing enhances understanding of tea plant evolution. Nat. Commun. 2020, 11, 4447. [Google Scholar] [CrossRef]
- Liu, Z.W.; Wu, Z.J.; Li, H.; Wang, Y.X.; Zhuang, J. l-theanine content and related gene expression: Novel insights into theanine biosynthesis and hydrolysis among different tea plant (Camellia sinensis L.) tissues and cultivars. Front. Plant Sci. 2017, 8, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, L.T.; Wang, X.Q.; Tan, H.B.; Liao, Y.Y.; Xu, P.; Kang, M.; Dong, F.; Yang, Z.Y. Alternative pathway to the formation of trans-cinnamic acid derived from L-phenylalanine in tea (Camellia sinensis) plants and other plants. J. Agric. Food Chem. 2020, 68, 3415–3424. [Google Scholar] [CrossRef]
- Liao, Y.Y.; Yu, Z.M.; Liu, X.Y.; Zeng, L.T.; Cheng, S.H.; Li, J.L.; Tang, J.C.; Yang, Z.Y. Effect of major tea insect attack on formation of quality-related nonvolatile specialized metabolites in tea (Camellia sinensis) leaves. J. Agric. Food Chem. 2019, 67, 6716–6724. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Wang, W.Z.; Li, Y.Z.; Dai, X.L.; Ma, G.L.; Xing, D.W.; Zhu, M.Q.; Gao, L.P.; Xia, T. Six phenylalanine ammonia-lyases from Camellia sinensis: Evolution, expression, and kinetics. Plant Physiol. Biochem. 2017, 118, 413–421. [Google Scholar] [CrossRef]
- Ashihara, H.; Deng, W.W.; Mullen, W.; Crozier, A. Distribution and biosynthesis of flavan-3-ols in Camellia sinensis seedlings and expression of genes encoding biosynthetic enzymes. Phytochemistry 2010, 71, 559–566. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, F.; Wan, Q.; Ruan, J.Y. Transcriptome analysis using RNA-Seq revealed the effects of nitrogen form on major secondary metabolite biosynthesis in tea (Camellia sinensis) plants. Acta Physiol. Plant. 2018, 40. [Google Scholar] [CrossRef]
- Ye, Y.L.; Yan, J.N.; Cui, J.L.; Mao, S.H.; Li, M.F.; Liao, X.L.; Tong, H.R. Dynamic changes in amino acids, catechins, caffeine and gallic acid in green tea during withering. J. Food Compos. Anal. 2018, 66, 98–108. [Google Scholar] [CrossRef]
- Li, D.; Li, C.Y.; Hu, C.J.; Yang, Y.S.; Lin, C.; Zhao, D.; Li, Q.S.; Ye, J.H.; Zheng, X.Q.; Liang, Y.R.; et al. Study on the accumulation mechanism of amino acids during bruising and withering treatment of oolong tea. J. Agric. Food Chem. 2020, 68, 14071–14080. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.T.; Liu, X.Y.; Gui, J.D.; Mei, X.; Fu, X.M.; Dong, F.; Tang, J.C.; Zhang, L.T.; Yang, Z.Y. Formation of (E)-nerolidol in tea (Camellia sinensis) leaves exposed to multiple stresses during tea manufacturing. Food Chem. 2017, 231, 78–86. [Google Scholar] [CrossRef]
- Yuan, L.J.; Liu, J.B.; Xiao, X.G. Biooxidation of indole and characteristics of the responsible enzymes. Afr. J. Biotechnol. 2011, 10, 19855–19863. [Google Scholar] [CrossRef]
- Gui, J.; Fu, X.; Zhou, Y.; Katsuno, T.; Mei, X.; Deng, R.; Xu, X.; Zhang, L.; Dong, F.; Watanabe, N.; et al. Does enzymatic hydrolysis of glycosidically bound volatile compounds really contribute to the formation of volatile compounds during the oolong tea manufacturing process? J. Agric. Food Chem. 2015, 63, 6905–6914. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, S.; Yang, Z.Y.; Katsuno, T.; Tu, V.A.; Mase, N.; Nakamura, Y.; Watanabe, N. Discrimination of green, oolong, and black teas by GC-MS analysis of characteristic volatile flavor compounds. Am. J. Anal. Chem. 2014, 5, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.J.; Li, D.; Ma, Y.X.; Zhang, W.; Lin, C.; Zheng, X.Q.; Liang, Y.R.; Lu, J.L. Formation mechanism of the oolong tea characteristic aroma during bruising and withering treatment. Food Chem. 2018, 269, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Liu, X.Y.; Zhou, Y.; Wang, X.Q.; Zeng, L.T.; Fu, X.M.; Li, J.L.; Tang, J.C.; Dong, F.; Yang, Z.Y. Formation and emission of linalool in tea (Camellia sinensis) leaves infested by tea green leafhopper (Empoasca (Matsumurasca) onukii Matsuda). Food Chem. 2017, 237, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Zeng, L.T.; Liao, Y.Y.; Gu, D.C.; Tang, J.C.; Yang, Z.Y. Influence of chloroplast defects on formation of jasmonic acid and characteristic aroma compounds in tea (Camellia sinensis) leaves exposed to postharvest stresses. Int. J. Mol. Sci. 2019, 20, 1044. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Y.; Zhang, Y.Z.; Ye, H.Z. Effect of different plucking season on aroma constituents in deep-fermented Lingtoudancong oolong tea. J. Tea Sci. 2007, 27, 236–242. (in Chinese). [Google Scholar]
- Zhou, P.; Hu, O.; Fu, H.; Ouyang, L.Q.; Gong, X.D.; Meng, P.; Wang, Z.; Dai, M.; Guo, X.M.; Wang, Y. UPLC-Q-TOF/MS-based untargeted metabolomics coupled with chemometrics approach for Tieguanyin tea with seasonal and year variations. Food Chem. 2019, 283, 73–82. [Google Scholar] [CrossRef]
- Tanan, T.T.; Nascimento, M.N.D.; Leite, R.D.S.; Guimarães, D.S. Spectrophotometric determinations of chloroplastidic pigments in Physalis gngulata L. leaves using different methodologies. J. Agric. Sci. 2017, 9. [Google Scholar] [CrossRef]
- Liang, Y.; Urano, D.; Liao, K.L.; Hedrick, T.L.; Gao, Y.; Jones, A.M. A nondestructive method to estimate the chlorophyll content of Arabidopsis seedlings. Plant Methods 2017, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Fu, X.M.; Liao, Y.Y.; Xu, X.L.; Zeng, L.T.; Tang, J.C.; Li, J.L.; Lai, J.H.; Yang, Z.Y. Differential accumulation of specialized metabolite l-theanine in green and albino-induced yellow tea (Camellia sinensis) leaves. Food Chem. 2019, 276, 93–100. [Google Scholar] [CrossRef]
- Hao, X.Y.; Horvath, D.P.; Chao, W.S.; Yang, Y.J.; Wang, X.C.; Xiao, B. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plant (Camellia sinensis (L.) O. Kuntze). Int. J. Mol. Sci. 2014, 15, 22155–22172. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collected Season | Treatment | Budding Density | 100-Bud Weight (g) | Yield (kg/hm2) |
|---|---|---|---|---|
| Spring | N0 | 3681.81 ± 874.85 c | 27.07 ± 1.44 c | 1.11 ± 0.35 c |
| N1 | 5481.81 ± 1188.47 b | 42.93 ± 0.44 b | 2.60 ± 0.51 b | |
| N2 | 7015.90 ± 924.15 a | 63.36 ± 7.60 a | 4.90 ± 1.10 a | |
| N3 | 7895.45 ± 748.38 a | 67.30 ± 3.42 a | 5.88 ± 0.63 a |
| Collected Season | Treatment | Budding Density | 100-Bud Weight (g) | Yield (kg/hm2) |
|---|---|---|---|---|
| Autumn | N0 | 5031.81 ± 263.00 b | 64.38 ± 1.75 c | 3.60 ± 0.25 c |
| N1 | 5890.90 ± 714.83 a | 72.05 ± 4.10 b | 4.69 ± 0.23 b | |
| N2 | 6238.63 ± 652.41 a | 76.76 ± 5.70 b | 5.31 ± 0.68 ab | |
| N3 | 6320.45 ± 454.93 a | 85.83 ± 3.02 a | 6.03 ± 0.53 a |
| Free Amino Acid (μg/g FW) | N0 | N1 | N2 | N3 |
| l-Theanine | 6812.34 ± 3645.83 c | 11,190.05 ± 5058.10 bc | 15,328.78 ± 401.43 ab | 20,477.66 ± 2475.90 a |
| l-Glutamic acid | 603.84 ± 334.96 c | 1026.84 ± 341.09 c | 1570.96 ± 116.33 b | 2081.24 ± 290.85 a |
| l-Glutamine | 845.10 ± 578.61 d | 1729.21 ± 566.34 c | 2601.55 ± 265.81 b | 3961.21 ± 437.59 a |
| Serine | 220.68 ± 18.62 b | 248.12 ± 24.89 ab | 295.71 ± 45.67 a | 285.02 ± 26.04 a |
| Threonine | 92.54 ± 13.11 c | 120.64 ± 14.63 b | 113.51 ± 9.20 b | 141.04 ± 10.19 a |
| Alanine | 74.57 ± 6.87 c | 106.01 ± 12.66 b | 132.19 ± 7.55 a | 138.90 ± 9.90 a |
| Valine | 50.88 ± 13.25 a | 48.63 ± 29.19 a | 59.72 ± 32.89 a | 29.47 ± 3.09 b |
| Phosphoserine | 61.76 ± 1.99 a | 60.19 ± 8.89 a | 65.71 ± 4.74 a | 57.05 ± 6.22 a |
| Phosphorylethanolamine | 35.43 ± 4.61 b | 39.71 ± 1.91 ab | 44.50 ± 0.88 a | 41.17 ± 2.50 a |
| Asparagine | 20.18 ± 1.56 b | 49.87 ± 20.7 a | 42.27 ± 7.07 a | 47.51 ± 6.73 a |
| Aspartate | 9.60 ± 1.58 b | 11.87 ± 1.81 ab | 14.13 ± 1.60 a | 12.73 ± 1.02 a |
| Glycine | 11.41 ± 1.09 a | 12.84 ± 1.62 a | 12.76 ± 1.66 a | 12.98 ± 1.18 a |
| Tyrosine | 9.66 ± 3.80 b | 20.20 ± 7.35 a | 22.11 ± 8.58 a | 24.42 ± 7.39 a |
| Phenylalanine | 9.85 ± 1.15 b | 14.42 ± 2.99 ab | 17.27 ± 7.32 a | 12.88 ± 0.30 ab |
| Gamma-aminobutyric acid | 17.63 ± 1.10 b | 22.53 ± 3.85 ab | 29.28 ± 10.32 a | 24.62 ± 4.07 ab |
| Histidine | 14.91 ± 3.21 c | 24.57 ± 4.02 bc | 44.79 ± 19.70 a | 32.93 ± 2.05 ab |
| Cystine | 6.39 ± 0.57 c | 11.86 ± 7.32 a | 8.99 ± 2.56 a | 7.97 ± 1.34 b |
| Leucine | 5.37 ± 0.18 b | 8.12 ± 1.89 ab | 11.03 ± 4.81 a | 9.27 ± 2.04 ab |
| Isoleucine | 6.45 ± 1.30 c | 9.14 ± 1.87 bc | 14.43 ± 6.01 a | 12.29 ± 0.96 ab |
| Tryptophan | 27.80 ± 2.90 a | 32.49 ± 3.55 a | 36.62 ± 12.55 a | 27.69 ± 5.88 a |
| Lysine | 6.92 ± 1.18 c | 8.69 ± 1.33 bc | 13.44 ± 5.02 a | 12.33 ± 0.63 a |
| β-Aminoisobutyric acid | 2.20 ± 0.25 ab | 1.77 ± 0.19 b | 6.38 ± 7.36 a | 2.30 ± 0.09 ab |
| α-Aminobutyric acid | 1.25 ± 0.68 b | 2.39 ± 2.65 a | 1.55 ± 1.32 ab | 1.79 ± 1.00 ab |
| β-Alanine | 4.18 ± 0.87 b | 4.40 ± 0.77 ab | 7.01 ± 3.17 a | 4.37 ± 0.15 ab |
| Taurine | 3.44 ± 0.06 a | 3.45 ± 0.16 a | 3.44 ± 0.14 a | 3.50 ± 0.09 a |
| Total free amino acid content | 8949.54 ± 4581.88 c | 14,804.82 ± 5983.96 bc | 20,499.23 ± 724.62 b | 27,464.77 ± 3124.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Wang, F.; Wu, Z.; Jiang, F.; Yu, W.; Yang, J.; Chen, J.; Jian, G.; You, Z.; Zeng, L. Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China. Metabolites 2021, 11, 146. https://doi.org/10.3390/metabo11030146
Chen Y, Wang F, Wu Z, Jiang F, Yu W, Yang J, Chen J, Jian G, You Z, Zeng L. Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China. Metabolites. 2021; 11(3):146. https://doi.org/10.3390/metabo11030146
Chicago/Turabian StyleChen, Yuzhen, Feng Wang, Zhidan Wu, Fuying Jiang, Wenquan Yu, Jie Yang, Jiaming Chen, Guotai Jian, Zhiming You, and Lanting Zeng. 2021. "Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China" Metabolites 11, no. 3: 146. https://doi.org/10.3390/metabo11030146
APA StyleChen, Y., Wang, F., Wu, Z., Jiang, F., Yu, W., Yang, J., Chen, J., Jian, G., You, Z., & Zeng, L. (2021). Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China. Metabolites, 11(3), 146. https://doi.org/10.3390/metabo11030146