
Changes of Hematological and Hemorheological Parameters in Rabbits with Hypercholesterolemia


 and
and
Abstract
:1. Introduction
2. Results
2.1. Bodyweight Changes
2.2. Hematological Parameters
2.3. Hemorheological Parameters
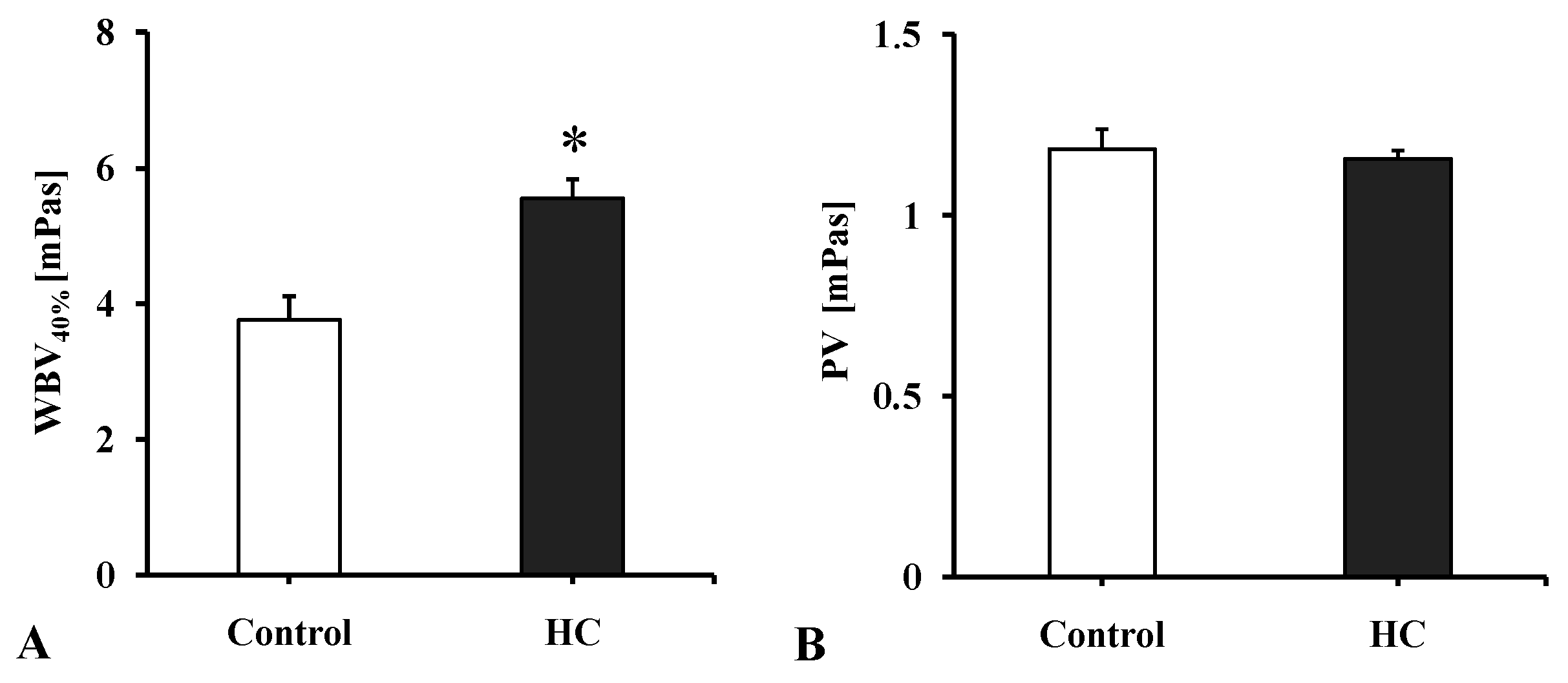
2.3.1. Whole Blood and Plasma Viscosity
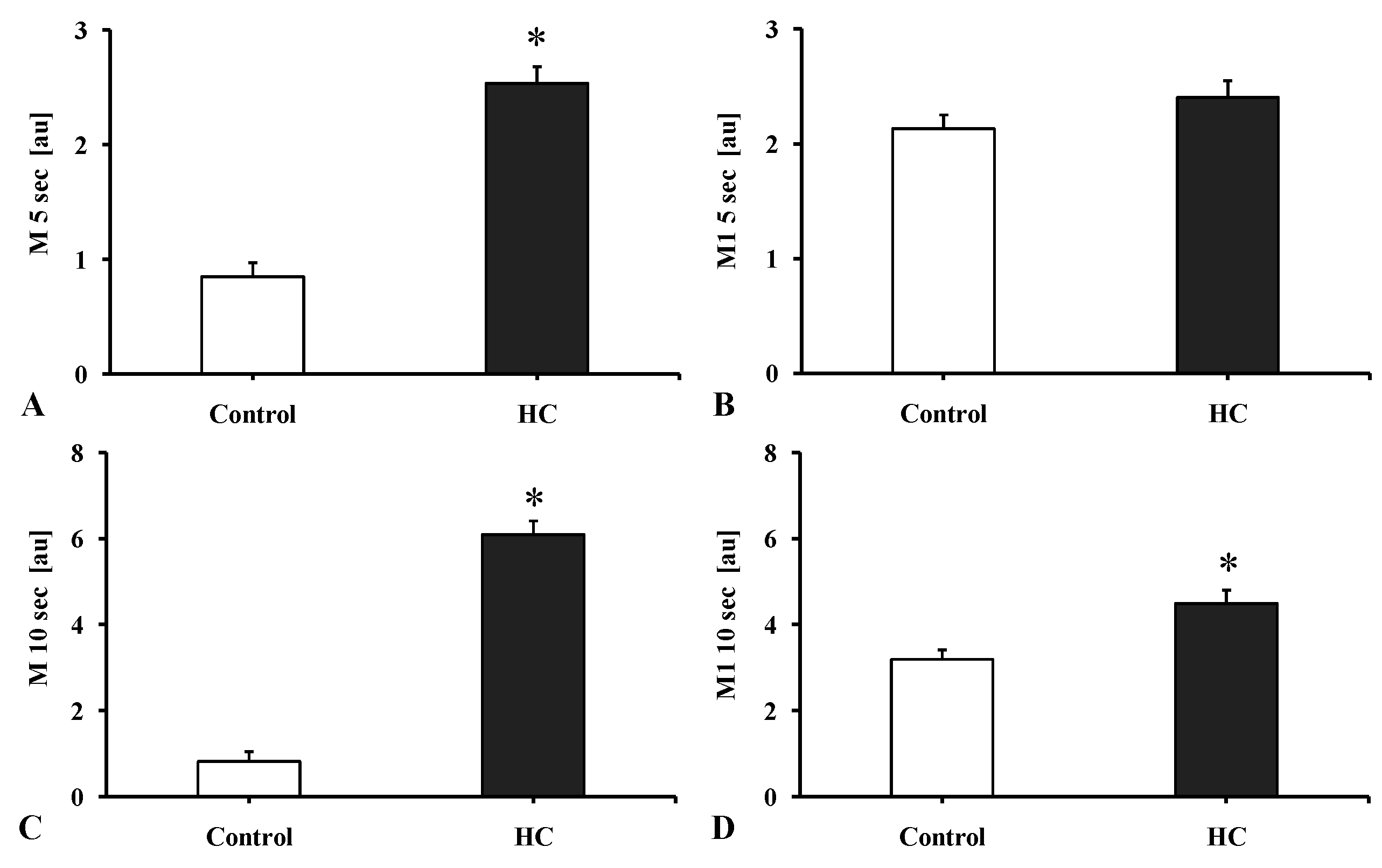
2.3.2. Red Blood Cell Aggregation
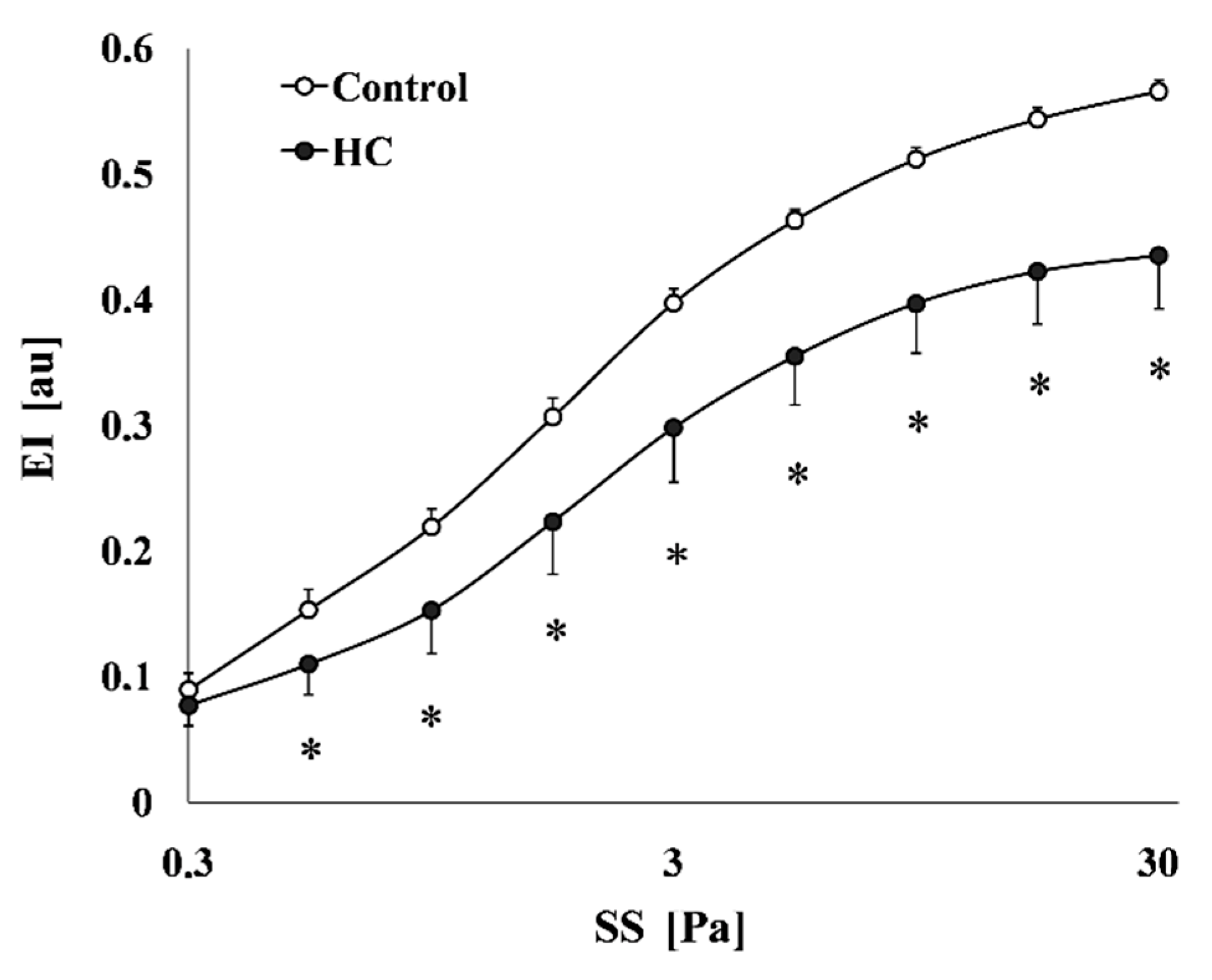
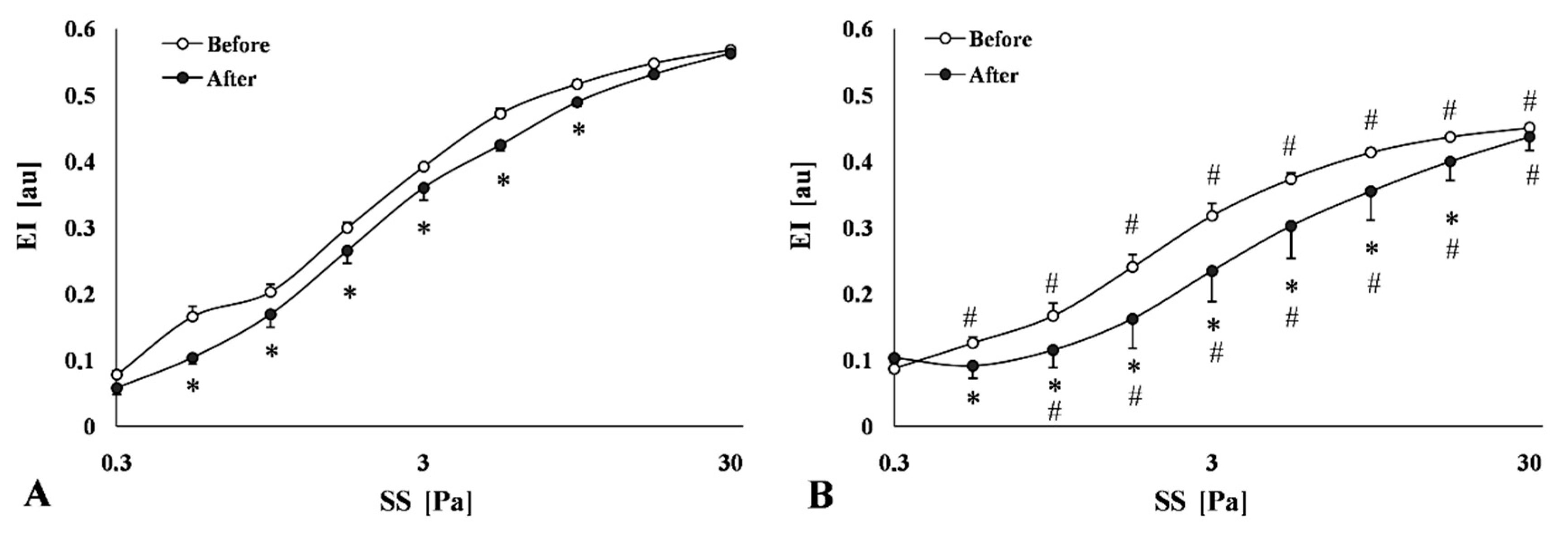
2.3.3. Red Blood Cell Deformability
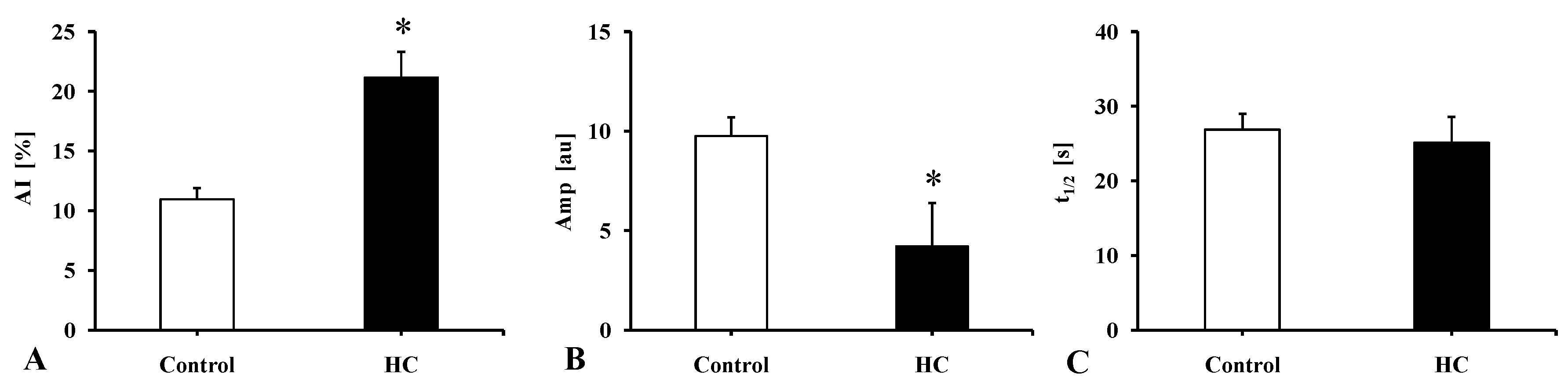
2.3.4. Red Blood Cell Membrane (Mechanical) Stability
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Collection of Blood Samples
4.3. Laboratory Methods
4.3.1. Hematological Parameters
4.3.2. Hemorheological Parameters
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rafieian-Kopaei, M.; Setorki, M.; Doudi, M.; Baradaran, A.; Nasri, H. Atherosclerosis: Process, indicators, risk factors and new hopes. Int. J. Prev. Med. 2014, 5, 927–946. [Google Scholar]
- Baker, I.A.; Pickering, J.; Elwood, P.C.; Bayer, A.; Ebrahim, S. Fibrinogen, viscosity and white blood cell count predict myocardial, but not cerebral infarction: Evidence from the Caerphilly and Speedwell cohort. Thromb. Haemost. 2002, 87, 421–425. [Google Scholar]
- Monsanto, H.A.; Renta-Muñoz, A.; Dones, W.; Comulada, A.; Cidre, C.; Orengo, J.C. The Puerto Rico Cardiovascular Risk-Estimation Study (PRCaRES): An exploratory assessment of new patients in physicians’ offices. P. R. Health Sci. J. 2014, 33, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Pizzi, C.; De Stavola, B.L.; Meade, T.W. Long-term association of routine blood count (Coulter) variables on fatal coronary heart disease: 30-year results from the first prospective Northwick Park Heart Study (NPHS-I). Int. J. Epidemiol. 2009, 39, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzoulaki, I.; Murray, G.D.; Lee, A.J.; Rumley, A.; Lowe, G.D.; Fowkes, F.G. Relative value of inflammatory, hemostatic, and rheological factors for incident myocardial infarction and stroke: The Edinburgh Artery Study. Circulation 2007, 115, 2119–2127. [Google Scholar] [CrossRef] [Green Version]
- Baskurt, O.K.; Meiselman, H.J. Blood rheology and hemodynamics. Semin. Thromb. Hemost. 2003, 29, 435–450. [Google Scholar] [PubMed] [Green Version]
- Koscielny, J.; Jung, E.M.; Mrowietz, C.; Kiesewetter, H.; Latza, R. Blood fluidity, fibrinogen, and cardiovascular risk factors of occlusive arterial disease: Results of the Aachen study. Clin. Hemorheol. Microcirc. 2004, 31, 185–195. [Google Scholar] [PubMed]
- Kolodgie, F.D.; Katocs, A.S., Jr.; Largis, E.E.; Wrenn, S.M.; Cornhill, J.F.; Herderick, E.E.; Lee, S.J.; Virmani, R. Hypercholesterolemia in the rabbit induced by feeding graded amounts of low-level cholesterol. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 1454–1464. [Google Scholar] [CrossRef]
- Yan, R.T.; Fernandes, V.; Yan, A.T.; Cushman, M.; Redheuil, A.; Tracy, R.; Vogel-Claussen, J.; Bahrami, H.; Nasir, K.; Bluemke, D.A.; et al. Fibrinogen and left ventricular myocardial systolic function: The Multi-Ethnic Study of Atherosclerosis (MESA). Am. Heart J. 2010, 160, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Dornas, W.C.; Oliveira, T.T.; Augusto, L.E.; Nagem, T.J. Experimental atherosclerosis in rabbits. Arq. Bras. Cardiol. 2010, 95, 272–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Kitajima, S.; Watanabe, T.; Xu, J.; Zhang, J.; Liu, E.; Chen, Y.E. Rabbit models for the study of human atherosclerosis: From pathophysiological mechanisms to translational medicine. Pharmacol. Ther. 2014, 146, 104–119. [Google Scholar] [CrossRef] [Green Version]
- Lozano, W.M.; Arias-Mutis, O.J.; Calvo, C.J.; Chorro, F.J.; Zarzoso, M. Diet-induced rabbit models for the study of metabolic syndrome. Animals 2019, 9, 463. [Google Scholar] [CrossRef] [Green Version]
- Milani-Nejad, N.; Janssen, P.M. Small and large animal models in cardiac contraction research: Advantages and disadvantages. Pharmacol. Ther. 2014, 141, 235–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição, G.; Heinonen, I.; Lourenço, A.P.; Duncker, D.J.; Falcão-Pires, I. Animal models of heart failure with preserved ejection fraction. Neth. Heart J. 2016, 24, 275–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windberger, U.; Bartholovitsch, A.; Plasenzotti, R.; Korak, K.J.; Heinze, G. Whole blood viscosity, plasma viscosity and erythrocyte aggregation in nine mammalian species: Reference values and comparison of data. Exp. Physiol. 2003, 88, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.; Alexy, T.; Furka, A.; Baskurt, O.K.; Meiselman, H.J.; Furka, I.; Miko, I. Inter-species differences in hematocrit to blood viscosity ratio. Biorheology 2009, 46, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Abdelhalim, M.A.K.; Al-Ayed, S.M.; Moussa, S.A.A.; Al-Mohy, Y.H. The changes in serum and whole blood rheological properties of rabbits during the progression of atherosclerosis. Pak. J. Pharm. Sci. 2016, 29, 1053–1057. [Google Scholar]
- Lee, Y.T.; Laxton, V.; Lin, H.Y.; Chan, Y.W.F.; Fitzgerald-Smith, S.; To, T.L.O.; Yan, B.P.; Liu, T.; Tse, G. Animal models of atherosclerosis. Biomed. Rep. 2017, 6, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priksz, D.; Bombicz, M.; Varga, B.; Kurucz, A.; Gesztelyi, R.; Balla, J.; Toth, A.; Papp, Z.; Szilvassy, Z.; Juhasz, B. Upregulation of myocardial and vascular phosphodiesterase 9a in a model of atherosclerotic cardiovascular disease. Int. J. Mol. Sci. 2018, 19, 2882. [Google Scholar] [CrossRef] [Green Version]
- Kertesz, A.; Bombicz, M.; Priksz, D.; Balla, J.; Balla, G.; Gesztelyi, R.; Varga, B.; Haines, D.D.; Tosaki, A.; Juhasz, B. Adverse impact of diet-induced hypercholesterolemia on cardiovascular tissue homeostasis in a rabbit model: Time-dependent changes in cardiac parameters. Int. J. Mol. Sci. 2013, 14, 19086–19108. [Google Scholar] [CrossRef] [Green Version]
- Abdelhalim, M.A.K.; Moussa, S.A.A. Biochemical changes of hemoglobin and osmotic fragility of red blood cells in high fat diet rabbits. Pak. J. Biol. Sci. 2010, 13, 73–77. [Google Scholar] [CrossRef]
- Króliczewska, B.; Miśta, D.; Zawadzki, W.; Wypchło, A.; Króliczewski, J. Effects of a skullcap root supplement on haematology, serum parameters and antioxidant enzymes in rabbits on a high-cholesterol diet. J. Anim. Physiol. Anim. Nutr. 2011, 95, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Karbiner, M.S.; Sierra, L.; Minahk, C.; Fonio, M.C.; Bruno, M.P.; Jerez, S. The role of oxidative stress in alterations of hematological parameters and inflammatory markers induced by early hypercholesterolemia. Life Sci. 2013, 93, 503–508. [Google Scholar] [CrossRef]
- Mozos, I. Mechanisms linking red blood cell disorders and cardiovascular diseases. Biomed. Res. Int. 2015, 2015, 682054. [Google Scholar] [CrossRef]
- Metivier, F.; Marchais, S.J.; Guerin, A.P.; Pannier, B.; London, G.M. Pathophysiology of anaemia: Focus on the heart and blood vessels. Nephrol. Dial. Transplant. 2000, 15 (Suppl. 3), 14–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felker, G.M.; Adams, K.F., Jr.; Gattis, W.A.; O’Connor, C.M. Anemia as a risk factor and therapeutic target in heart failure. J. Am. Coll. Cardiol. 2004, 44, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, T.; Luft, C.; Haidinger, D.; Poelz, W.; Haltmayer, M. Erythrocyte mean corpuscular volume associated with the anatomical distribution in peripheral arterial disease. VASA 2002, 31, 81–85. [Google Scholar] [CrossRef]
- Wang, N.; Tall, A.R. Cholesterol in platelet biogenesis and activation. Blood 2016, 127, 1949–1953. [Google Scholar] [CrossRef] [Green Version]
- Tomaiuolo, G. Biomechanical properties of red blood cells in health and disease towards microfluidics. Biomicrofluidics 2014, 8, 051501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Ley, K.F. Role of platelets in the development of atherosclerosis. Trends Cardiovasc. Med. 2004, 14, 18–22. [Google Scholar] [CrossRef]
- Barale, C.; Cavalot, F.; Frascaroli, C.; Bonomo, K.; Morotti, A.; Guerrasio, A.; Russo, I. Association between high on-aspirin platelet reactivity and reduced superoxide dismutase activity in patients affected by type 2 diabetes mellitus or primary hypercholesterolemia. Int. J. Mol. Sci. 2020, 21, 4983. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Hill, M.A.; Wu, J. Does C-reactive protein contribute to atherothrombosis via oxidant-mediated release of pro-thrombotic factors and activation of platelets? Front. Physiol. 2012, 3, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braune, S.; Küpper, J.H.; Jung, F. Effect of prostanoids on human platelet function: An overview. Int. J. Mol. Sci. 2020, 21, 9020. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Blocki, A.; Franke, R.P.; Jung, F. Vascular endothelial cell biology: An update. Int. J. Mol. Sci. 2019, 20, E4411. [Google Scholar] [CrossRef] [Green Version]
- Buchwalow, I.B.; Cacanyiova, S.; Neumann, J.; Samoilova, V.E.; Boecker, W.; Kristek, F. The role of arterial smooth muscle in vasorelaxation. Biochem. Biophys. Res. Commun. 2008, 377, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H. White blood cell and platelet counts could affect whole blood viscosity. J. Chin. Med. Assoc. 2004, 67, 394–397. [Google Scholar] [PubMed]
- Wiewiora, M.; Sosada, K.; Wylezol, M.; Slowinska, L.; Zurawinski, W. Red blood cell aggregation and deformability among patients qualified for bariatric surgery. Obes. Surg. 2007, 17, 365–371. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Sternitzky, R.; Pindur, G.; Rampling, M.; Franke, R.P.; Jung, F. Erythrocyte aggregation in relation to plasma proteins and lipids. J. Cell. Biotechnol. 2019, 5, 65–70. [Google Scholar] [CrossRef]
- Rampling, M.W. Hyperviscosity as a complication in a variety of disorders. Semin. Thromb. Hemost. 2003, 29, 459–465. [Google Scholar] [PubMed]
- Baskurt, O.K.; Yalcin, O.; Ozdem, S.; Armstrong, J.K.; Meiselman, H.J. Modulation of endothelial nitric oxide synthase expression by red blood cell aggregation. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H222–H229. [Google Scholar] [CrossRef] [Green Version]
- Baskurt, O.K.; Boynard, M.; Cokelet, G.C.; Connes, P.; Cooke, B.M.; Forconi, S.; Liao, F.; Hardeman, M.R.; Jung, F.; Meiselman, H.J.; et al. International Expert Panel for Standardization of Hemorheological Methods. New guidelines for hemorheological laboratory techniques. Clin. Hemorheol. Microcirc. 2009, 42, 75–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskurt, O.K.; Neu, B.; Meiselman, H.J. Determinants of red blood cell aggregation. In Red Blood Cell Aggregation; Baskurt, O.K., Neu, B., Meiselman, H.J., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 9–29. [Google Scholar]
- Geddes, J.B.; Carr, R.T.; Wu, F.; Lao, Y.; Maher, M. Blood flow in microvascular networks: A study in nonlinear biology. Chaos 2010, 20, 045123. [Google Scholar] [CrossRef] [Green Version]
- Lipowsky, H.H. Microvascular rheology and hemodynamics. Microcirculation 2005, 12, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Baskurt, O.K. Mechanisms of blood rheology alterations. In Handbook of Hemorheology and Hemodynamics; Baskurt, O.K., Hardeman, M.R., Rampling, M.W., Meiselman, H.J., Eds.; IOS Press: Amsterdam, The Netherlands, 2007; pp. 170–190. [Google Scholar]
- Muravyov, A.V.; Tikhomirova, I.A.; Maimistova, A.A.; Bulaeva, S.V.; Mikhailov, P.V.; Kislov, N.V. Red blood cell aggregation changes are depended on its initial value: Effect of long-term drug treatment and short-term cell incubation with drug. Clin. Hemorheol. Microcirc. 2011, 48, 231–240. [Google Scholar] [CrossRef]
- Munn, L.L.; Dupin, M.M. Blood cell interactions and segregation in flow. Ann. Biomed. Eng. 2008, 36, 534–544. [Google Scholar] [CrossRef]
- Nash, G.B.; Watts, T.; Thornton, C.; Barigou, M. Red cell aggregation as a factor influencing margination and adhesion of leukocytes and platelets. Clin. Hemorheol. Microcirc. 2008, 39, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, D.; Gompper, G. White blood cell margination in microcirculation. Soft Matter 2014, 10, 2961–2970. [Google Scholar] [CrossRef] [Green Version]
- Pretini, V.; Koenen, M.H.; Kaestner, L.; Fens, M.H.A.M.; Schiffelers, R.M.; Bartels, M.; Van Wijk, R. Red Blood Cells: Chasing Interactions. Front. Physiol. 2019, 10, 945. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.; Sogor, V.; Kiss, F.; Ulker, P. Interspecies diversity of erythrocyte mechanical stability at various combinations in magnitude and duration of shear stress, and osmolality. Clin. Hemorheol. Microcirc. 2016, 63, 381–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, T.M.; Braun, A.; Trigatti, B.L.; Brugnara, C.; Sakamoto, M.; Krieger, M.; Andrews, N.C. Failure of red blood cell maturation in mice with defects in the high-density lipoprotein receptor SR-BI. Blood 2002, 99, 1817–1824. [Google Scholar] [CrossRef]
- Meurs, I.; Hoekstra, M.; van Wanrooij, E.J.; Hildebrand, R.B.; Kuiper, J.; Kuipers, F.; Hardeman, M.R.; Van Berkel, T.J.; Van Eck, M. HDL cholesterol levels are an important factor for determining the lifespan of erythrocytes. Exp. Hematol. 2005, 33, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- van Zwieten, R.; Bochem, A.E.; Hilarius, P.M.; van Bruggen, R.; Bergkamp, F.; Hovingh, G.K.; Verhoeven, A.J. The cholesterol content of the erythrocyte membrane is an important determinant of phosphatidylserine exposure. Biochim. Biophys. Acta 2012, 1821, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Unruh, D.; Srinivasan, R.; Benson, T.; Haigh, S.; Coyle, D.; Batra, N.; Keil, R.; Sturm, R.; Blanco, V.; Palascak, M.; et al. Red blood cell dysfunction induced by high-fat diet: Potential implications for obesity-related atherosclerosis. Circulation 2015, 132, 1898–1908. [Google Scholar] [CrossRef] [Green Version]
- Koter, M.; Franiak, I.; Strychalska, K.; Broncel, M.; Chojnowska-Jezierska, J. Damage to the structure of erythrocyte plasma membranes in patients with type-2 hypercholesterolemia. Int. J. Biochem. Cell Biol. 2004, 36, 205–215. [Google Scholar] [CrossRef]
- Manno, S.; Takakuwa, Y.; Mohandas, N. Identification of a functional role for lipid asymmetry in biological membranes: Phosphatidylserine-skeletal protein interactions modulate membrane stability. Proc. Natl. Acad. Sci. USA 2002, 99, 1943–1948. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Kim, K.C.; Park, H.W.; Song, J.H.; Lee, C.H. Rheological properties of erythrocytes from male hypercholesterolemia. Microvasc. Res. 2004, 67, 133–138. [Google Scholar] [CrossRef] [PubMed]
- da Silva Garrote-Filho, M.; Bernardino-Neto, M.; Penha-Silva, N. Influence of erythrocyte membrane stability in atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 17. [Google Scholar] [CrossRef]
- Vayá, A.; Riveram, L.; de la Espriella, R.; Sanchez, F.; Suescun, M.; Hernandez, J.L.; Fácila, L. Red blood cell distribution width and erythrocyte deformability in patients with acute myocardial infarction. Clin. Hemorheol. Microcirc. 2015, 59, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Maj, D.; Bieniek, J.; Łapa, P.; Sternstein, I. The effect of crossing New Zealand White with Californian rabbits on growth and slaughter traits. Arch. Anim. Breed. 2009, 52, 205–211. [Google Scholar] [CrossRef]
- Matrai, A.; Whittington, R.B.; Ernst, E. A simple method of estimating whole blood viscosity at standardized hematocrit. Clin. Hemorheol. 1987, 7, 261–265. [Google Scholar] [CrossRef]
- Hardeman, M.; Goedhart, P.; Shin, S. Methods in hemorheology. In Handbook of Hemorheology and Hemodynamics; Baskurt, O.K., Hardeman, M.R., Rampling, M.W., Meiselman, H.J., Eds.; IOS Press: Amsterdam, The Netherlands, 2007; pp. 242–266. [Google Scholar]
- Baskurt, O.K.; Hardeman, M.R.; Uyuklu, M.; Ulker, P.; Cengiz, M.; Nemeth, N.; Shin, S.; Alexy, T.; Meiselman, H.J. Parameterization of red blood cell elongation index--shear stress curves obtained by ektacytometry. Scand. J. Clin. Lab. Investig. 2009, 69, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Sogor, V.; Tanczos, B.; Deak, A. Data interpretation of erythrocyte membrane mechanical stability test using the laser-assisted optical rotational cell analyzer. Ser. Biomech. 2016, 30, 27–34. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hematological Parameter | Control (n = 6) | HC (n = 6) | p Value vs. Control |
|---|---|---|---|
| White blood cell count [×109/L] | 8.375 ± 0.270 | 23.59 ± 4.762 | 0.0042 |
| Red blood cell count [×1012/L] | 6.863 ± 0.125 | 3.758 ± 0.245 | <0.0001 |
| Hemoglobin [g/dL] | 15.25 ± 0.272 | 8.392 ± 0.558 | <0.0001 |
| Hematocrit [%] | 46.37 ± 0.929 | 29.38 ± 1.729 | <0.0001 |
| Mean corpuscular volume [fL] | 67.58 ± 0.472 | 78.88 ± 2.426 | 0.0001 |
| Mean corpuscular hemoglobin [pg] | 22.22 ± 0.258 | 22.50 ± 0.757 | n.s. |
| Mean corpuscular hemoglobin concentration [g/L] | 32.90 ± 0.270 | 28.82 ± 1.270 | 0.0047 |
| Platelet count [×109/L] | 254.8 ± 27.54 | 481.5 ± 38.73 | <0.0001 |
| Parameter | Control | HC | p Value vs. Control |
|---|---|---|---|
| EI at 3Pa | 0.591 ± 0.009 | 0.448 ± 0.051 | <0.0001 |
| EImax | 0.397± 0.011 | 0.298 ± 0.043 | <0.0001 |
| SS1/2 [Pa] | 1.544 ± 0.172 | 2.125 ± 0.633 | 0.0272 |
| EImax/SS1/2 [Pa−1] | 0.387 ± 0.044 | 0.228 ± 0.069 | 0.0007 |
| Parameter | Test | Control | HC | p Value vs. Control, or vs. before (* Control, # HC) |
|---|---|---|---|---|
| EI at 3Pa | before (B) | 0.392 ± 0.002 | 0.318 ± 0.015 | 0.002 |
| after (A) | 0.360 ± 0.008 * | 0.235 ± 0.019 # | 0.0015; * 0.002; # 0.006 | |
| ratio (A/B) | 0.919 ± 0.019 | 0.734 ± 0.038 | <0.0001 | |
| EImax | before (B) | 0.592 ± 0.003 | 0.461 ± 0.017 | <0.0001 |
| after (A) | 0.595 ± 0.004 | 0.458 ± 0.011 | <0.0001 | |
| ratio (A/B) | 1.005 ± 0.007 | 0.999 ± 0.033 | ns | |
| SS½ [Pa] | before (B) | 1.690 ± 0.081 | 1.977 ± 0.227 | ns |
| after (A) | 1.960 ± 0.149 | 4.540 ± 0.875 # | 0.009; # 0.018 | |
| ratio (A/B) | 1.169 ± 0.089 | 2.228 ± 0.280 | 0.001 | |
| EImax/SS1/2 [Pa−1] | before (B) | 0.355 ± 0.021 | 0.249 ± 0.028 | 0.012 |
| after (A) | 0.311 ± 0.019 | 0.124 ± 0.025 # | 0.0318; # 0.002 | |
| ratio (A/B) | 0.883 ± 0.062 | 0.484 ± 0.062 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanczos, B.; Somogyi, V.; Bombicz, M.; Juhasz, B.; Nemeth, N.; Deak, A. Changes of Hematological and Hemorheological Parameters in Rabbits with Hypercholesterolemia. Metabolites 2021, 11, 249. https://doi.org/10.3390/metabo11040249
Tanczos B, Somogyi V, Bombicz M, Juhasz B, Nemeth N, Deak A. Changes of Hematological and Hemorheological Parameters in Rabbits with Hypercholesterolemia. Metabolites. 2021; 11(4):249. https://doi.org/10.3390/metabo11040249
Chicago/Turabian StyleTanczos, Bence, Viktoria Somogyi, Mariann Bombicz, Bela Juhasz, Norbert Nemeth, and Adam Deak. 2021. "Changes of Hematological and Hemorheological Parameters in Rabbits with Hypercholesterolemia" Metabolites 11, no. 4: 249. https://doi.org/10.3390/metabo11040249
APA StyleTanczos, B., Somogyi, V., Bombicz, M., Juhasz, B., Nemeth, N., & Deak, A. (2021). Changes of Hematological and Hemorheological Parameters in Rabbits with Hypercholesterolemia. Metabolites, 11(4), 249. https://doi.org/10.3390/metabo11040249