
In Vivo Estimation of Ketogenesis Using Metabolic Flux Analysis—Technical Aspects and Model Interpretation

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
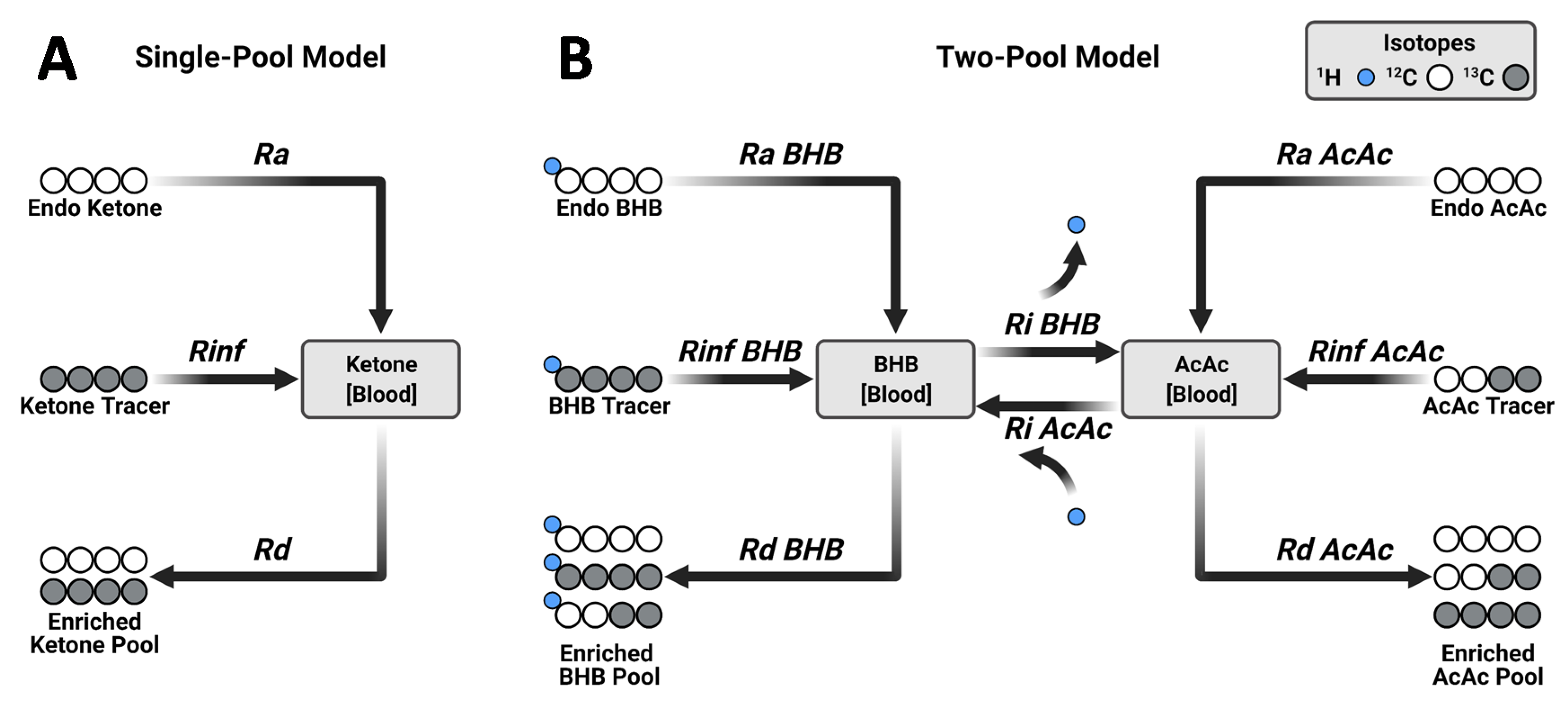
2.1. Tracer Approaches to Study Ketogenesis
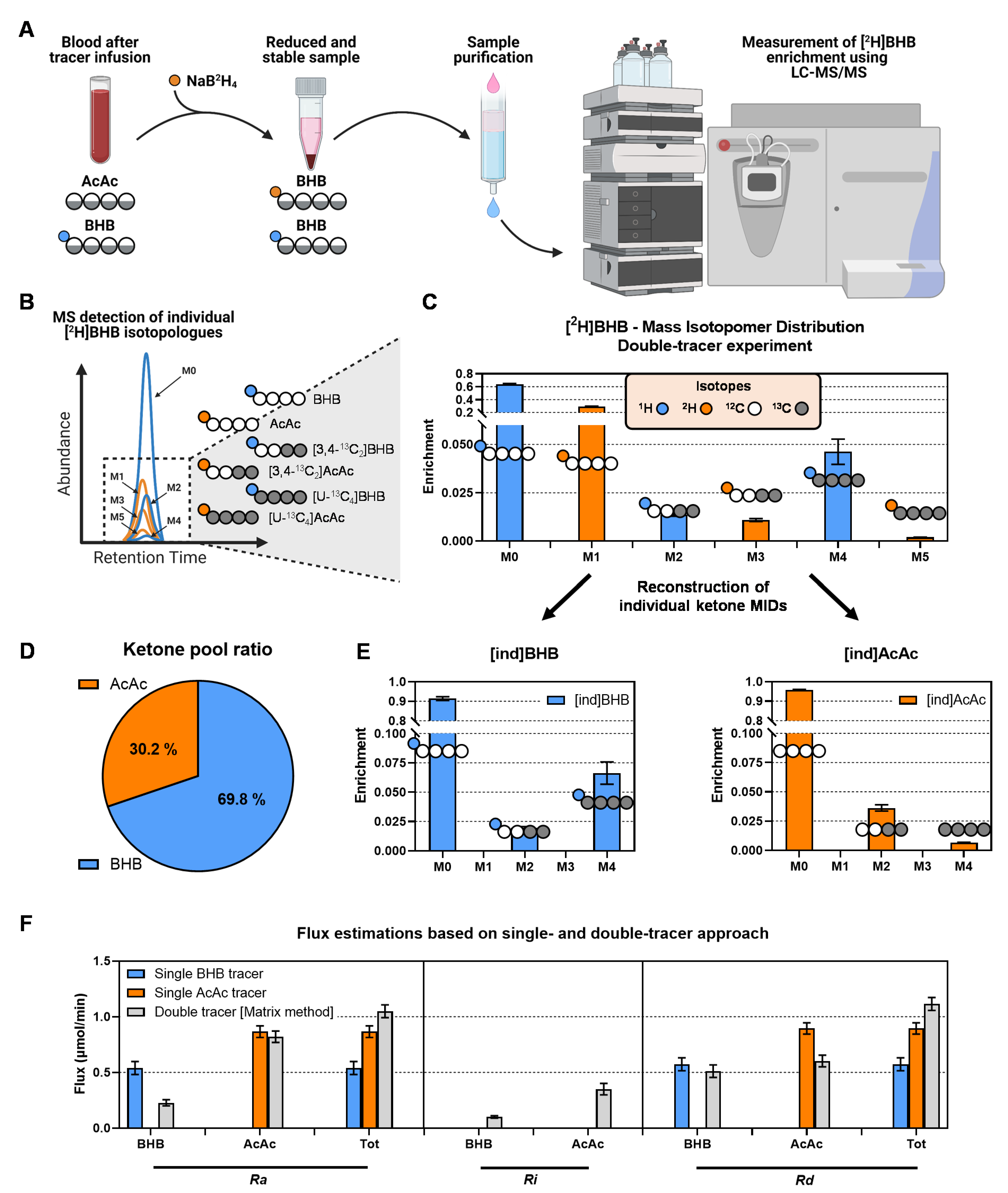
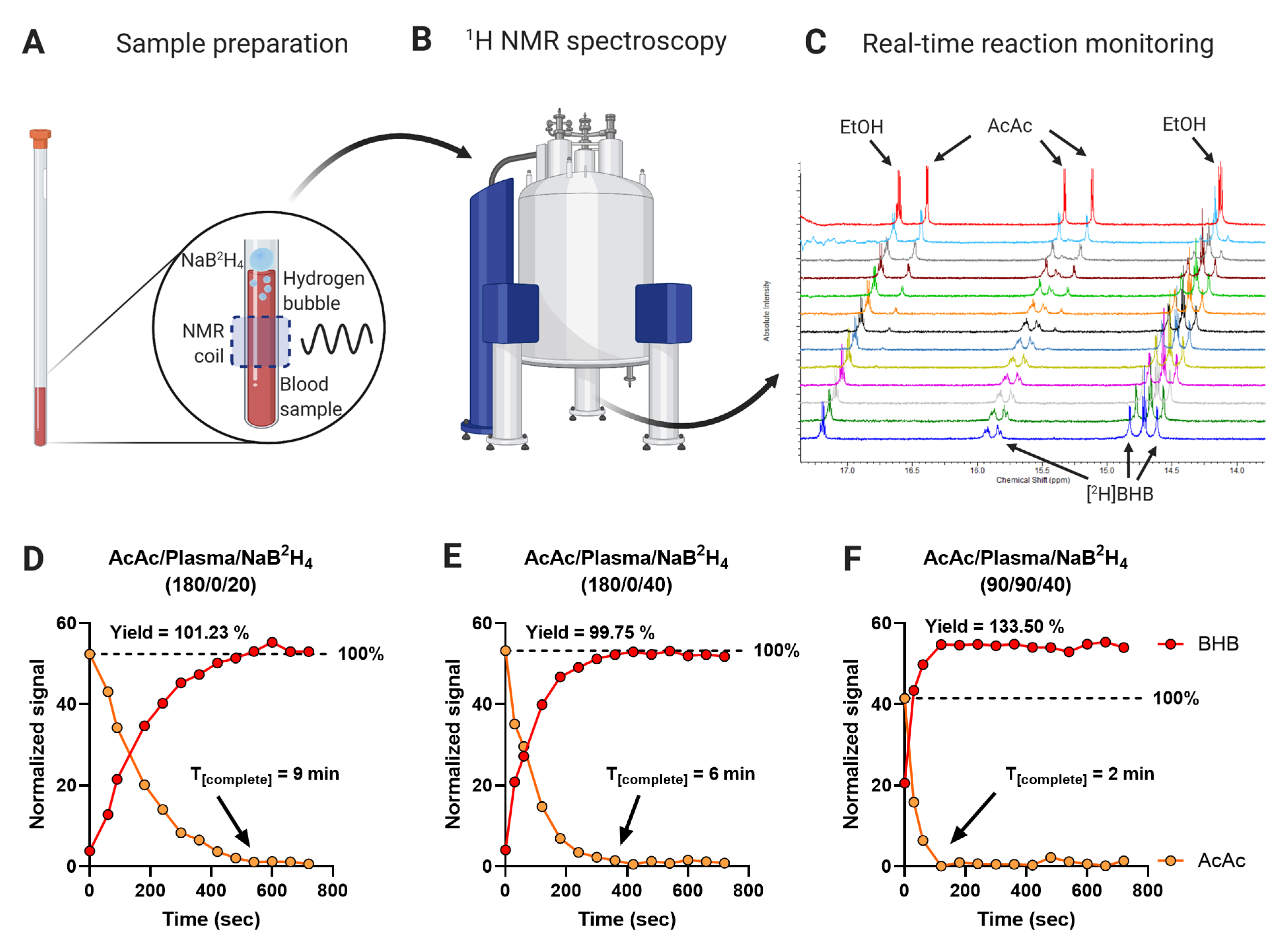
2.2. In Situ Reduction of AcAc to [2H]BHB Is Rapid and Complete
2.3. Stability of the AcAc Tracer
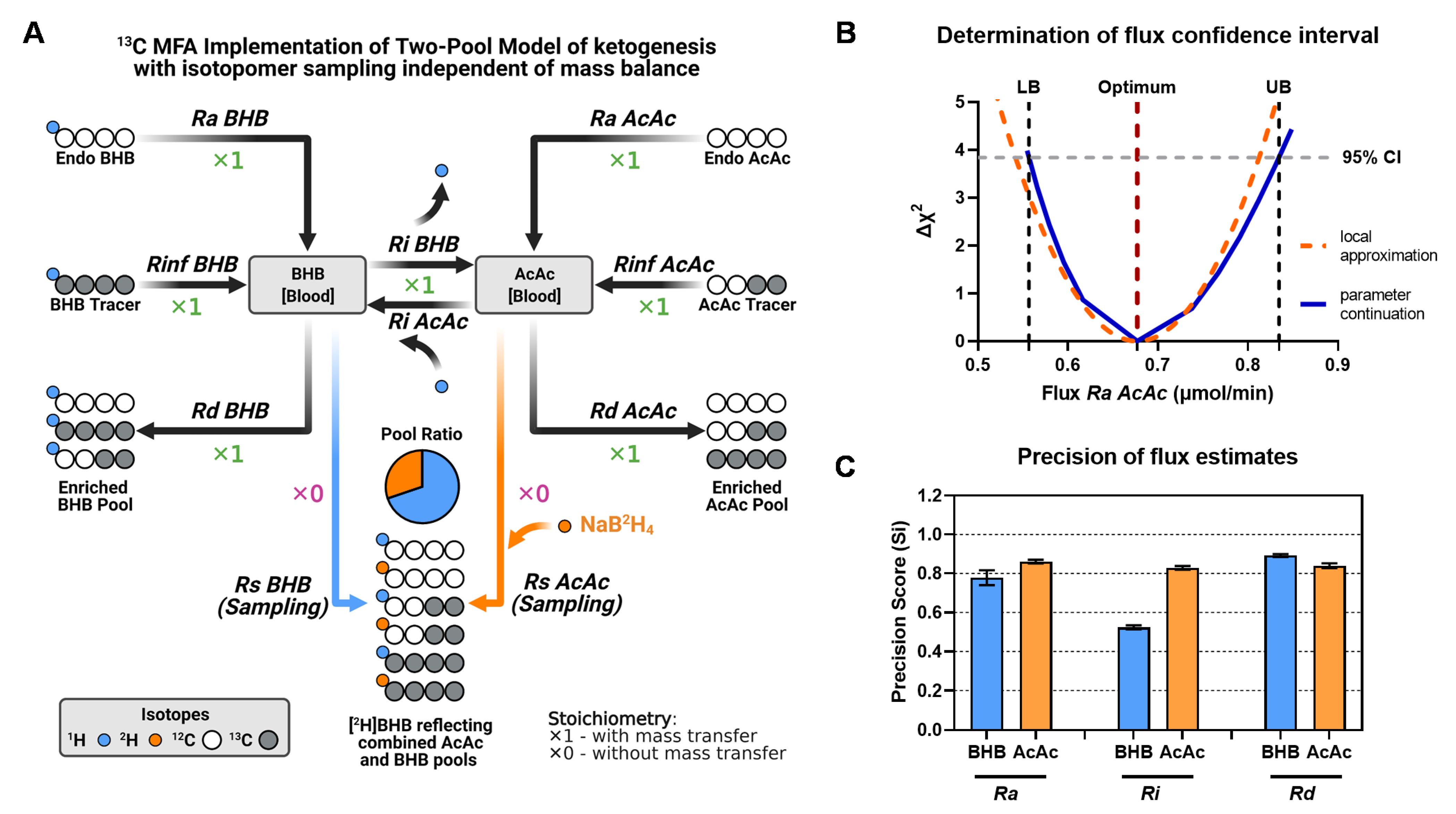
2.4. MFA Implementation of the Two-Pool Model of Ketogenesis
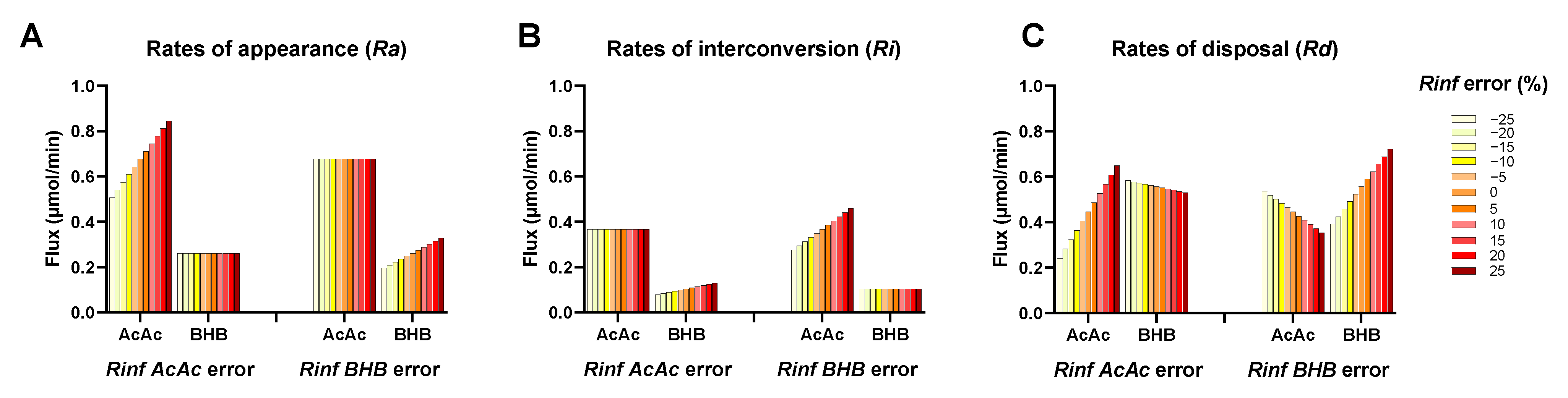
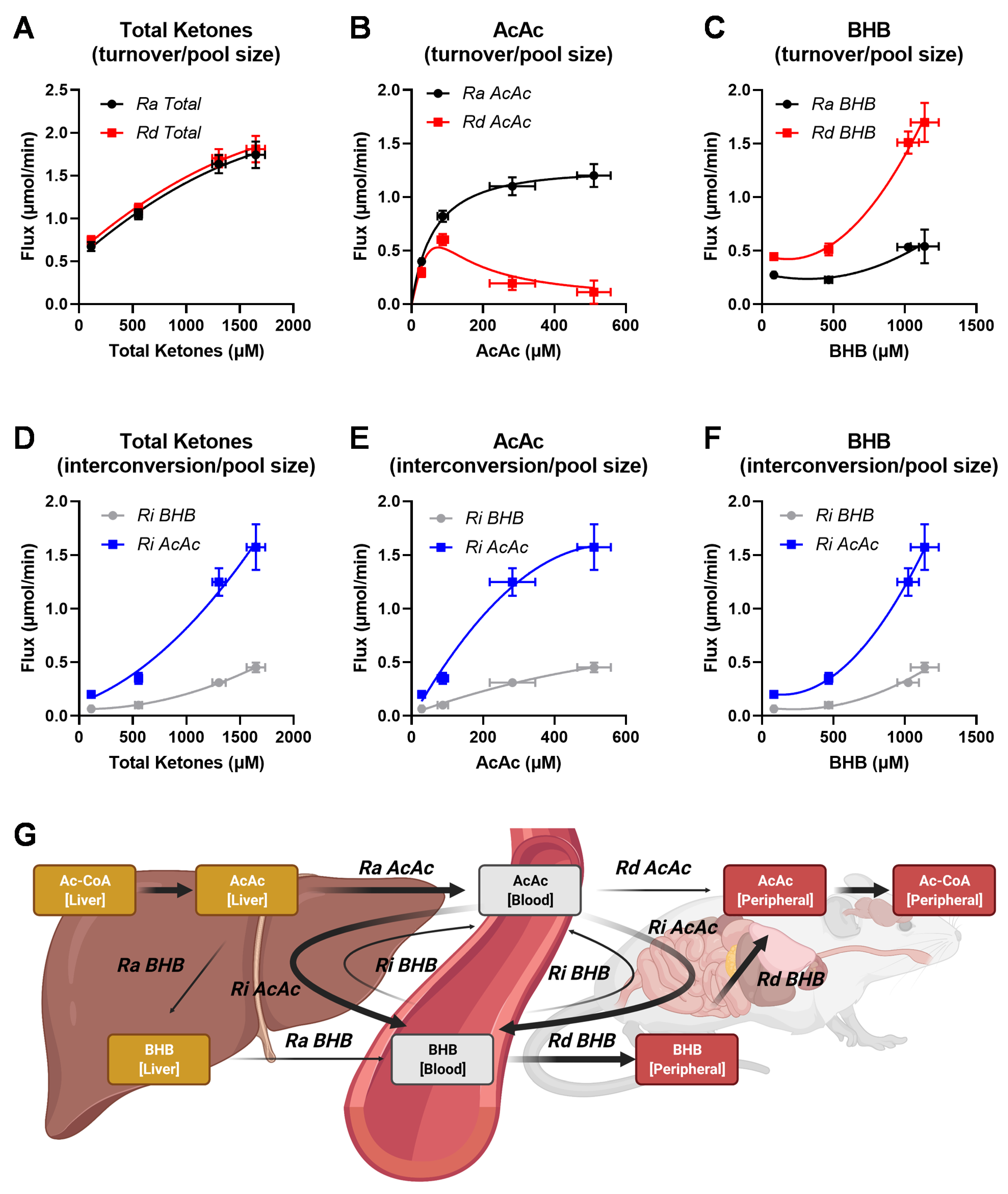
2.5. Ketone Fluxes Change Non-Linearly with the Progression of Ketosis
3. Discussion
3.1. Technical Aspects of In Vivo Estimation of Ketogenesis
3.2. Advantages of the MFA Implementation of a Two-Pool Model of Ketogenesis
3.3. Interpretation of Ketone Interconversions in Fasted Mice
4. Materials and Methods
4.1. Animals
4.2. Stable Isotope Tracer Preparation
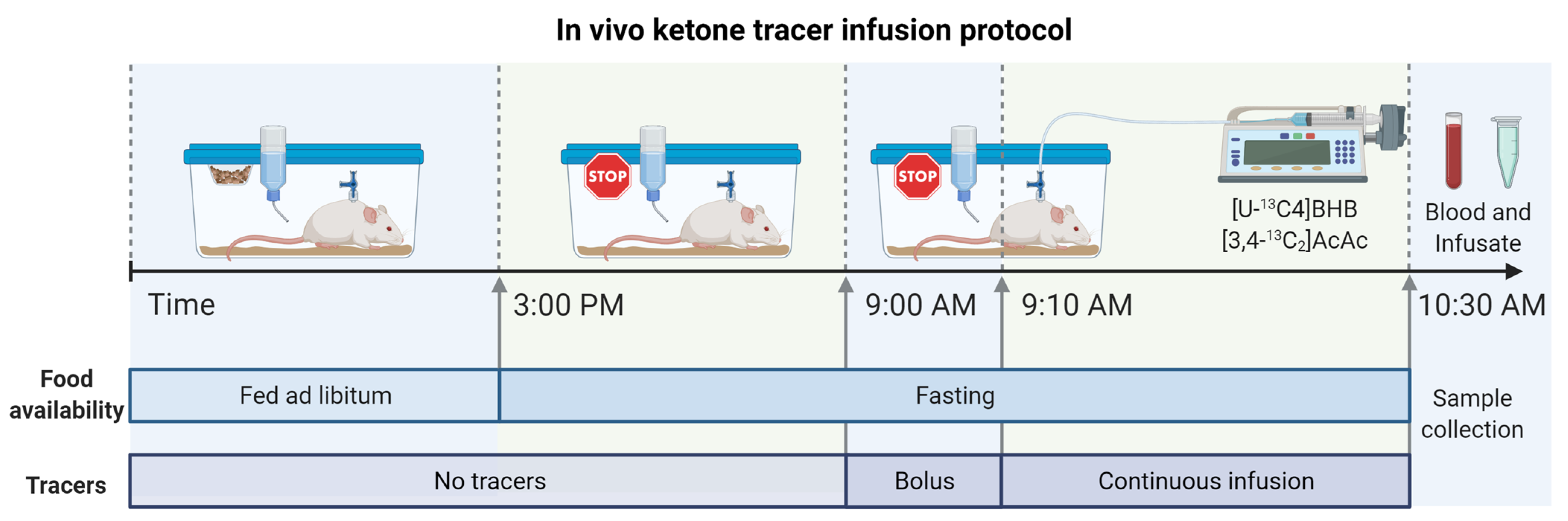
4.3. In Vivo Double-Tracer Infusions
4.4. Measurement of Ketone Body Concentration
4.5. In Situ Reduction of AcAc and Sample Purification
4.6. Measurement of Ketone Enrichment by LC-MS/MS
4.7. Real-Time Reaction Monitoring by 1H NMR Spectroscopy
4.8. Flux Calculations Using Analytical Equations (Matrix Method)
4.9. Metabolic Network and Flux Modeling Using MFA
4.10. Calculation of Precision Score
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, A.M.; Williamson, D.H. Physiological roles of ketone bodies as substrates and signals in mammalian tissues. Physiol. Rev. 1980, 60, 143–187. [Google Scholar] [CrossRef]
- Desrochers, S.; Dubreuil, P.; Brunet, J.; Jette, M.; David, F.; Landau, B.R.; Brunengraber, H. Metabolism of (R,S)-1,3-butanediol acetoacetate esters, potential parenteral and enteral nutrients in conscious pigs. Am. J. Physiol. 1995, 268, E660–E667. [Google Scholar] [CrossRef]
- Kim, I.Y.; Suh, S.H.; Lee, I.K.; Wolfe, R.R. Applications of stable, nonradioactive isotope tracers in in vivo human metabolic research. Exp. Mol. Med. 2016, 48, e203. [Google Scholar] [CrossRef] [Green Version]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarry, J.D. What if Minkowski had been ageusic? An alternative angle on diabetes. Science 1992, 258, 766–770. [Google Scholar] [CrossRef]
- Jefferies, C.A.; Nakhla, M.; Derraik, J.G.; Gunn, A.J.; Daneman, D.; Cutfield, W.S. Preventing Diabetic Ketoacidosis. Pediatric Clin. N. Am. 2015, 62, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Roman, M.A.; Burgess, S.C.; Browning, J.D. Metabolomics, stable isotopes, and A-β+ ketosis-prone diabetes. Diabetes 2013, 62, 682–684. [Google Scholar] [CrossRef] [Green Version]
- Satapati, S.; He, T.; Inagaki, T.; Potthoff, M.; Merritt, M.E.; Esser, V.; Mangelsdorf, D.J.; Kliewer, S.A.; Browning, J.D.; Burgess, S.C. Partial resistance to peroxisome proliferator-activated receptor-alpha agonists in ZDF rats is associated with defective hepatic mitochondrial metabolism. Diabetes 2008, 57, 2012–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.A.; Deja, S.; Satapati, S.; Fu, X.; Burgess, S.C.; Browning, J.D. Impaired ketogenesis and increased acetyl-CoA oxidation promote hyperglycemia in human fatty liver. JCI Insight 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- d’Avignon, D.A.; Puchalska, P.; Ercal, B.; Chang, Y.; Martin, S.E.; Graham, M.J.; Patti, G.J.; Han, X.; Crawford, P.A. Hepatic ketogenic insufficiency reprograms hepatic glycogen metabolism and the lipidome. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, K.; Nikkanen, J.; Man, K.; Leong, Y.A.; Sogawa, Y.; Maschek, J.A.; Van Ry, T.; Chagwedera, D.N.; Cox, J.E.; Chawla, A. Energetic Trade-Offs and Hypometabolic States Promote Disease Tolerance. Cell 2019, 177, 399–413.e312. [Google Scholar] [CrossRef] [Green Version]
- Puchalska, P.; Martin, S.E.; Huang, X.; Lengfeld, J.E.; Daniel, B.; Graham, M.J.; Han, X.; Nagy, L.; Patti, G.J.; Crawford, P.A. Hepatocyte-Macrophage Acetoacetate Shuttle Protects against Tissue Fibrosis. Cell Metab. 2019, 29, 383–398.e387. [Google Scholar] [CrossRef] [Green Version]
- Dong, T.A.; Sandesara, P.B.; Dhindsa, D.S.; Mehta, A.; Arneson, L.C.; Dollar, A.L.; Taub, P.R.; Sperling, L.S. Intermittent Fasting: A Heart Healthy Dietary Pattern? Am. J. Med. 2020, 133, 901–907. [Google Scholar] [CrossRef]
- O’Neill, B.; Raggi, P. The ketogenic diet: Pros and cons. Atherosclerosis 2020, 292, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Gershuni, V.M.; Yan, S.L.; Medici, V. Nutritional Ketosis for Weight Management and Reversal of Metabolic Syndrome. Curr. Nutr. Rep. 2018, 7, 97–106. [Google Scholar] [CrossRef]
- Browning, J.D.; Baker, J.A.; Rogers, T.; Davis, J.; Satapati, S.; Burgess, S.C. Short-term weight loss and hepatic triglyceride reduction: Evidence of a metabolic advantage with dietary carbohydrate restriction. Am. J. Clin. Nutr. 2011, 93, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Seaborg, K.; Wang, X.; Olson, C.; Felton, E.A. Pediatric to Adult Transitions of Ketogenic Dietary Therapy for Epilepsy. J. Child Neurol. 2020, 35, 896–900. [Google Scholar] [CrossRef]
- Kossoff, E.H. The ketogenic diet: An appropriate first-line therapy? Expert Rev. Neurother. 2010, 10, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Heath, D.F.; Barton, R.N. The design of experiments using isotopes for the determination of the rates of disposal of blood-borne substrates in vivo with special reference to glucose, ketone bodies, free fatty acids and proteins. Biochem. J. 1973, 136, 503–518. [Google Scholar] [CrossRef] [Green Version]
- McGarry, J.D.; Guest, M.J.; Foster, D.W. Ketone body metabolism in the ketosis of starvation and alloxan diabetes. J. Biol. Chem. 1970, 245, 4382–4390. [Google Scholar] [CrossRef]
- Barton, R.N. Isotopic study of ketone body kinetics: Invalidity of calculations based upon specific radioactivity of total ketone bodies. Metab. Clin. Exp. 1980, 29, 392–396. [Google Scholar] [CrossRef]
- Cobelli, C.; Nosadini, R.; Toffolo, G.; McCulloch, A.; Avogaro, A.; Tiengo, A.; Alberti, K.G. Model of the kinetics of ketone bodies in humans. Am. J. Physiol. 1982, 243, R7–R17. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.M.; Schwenk, W.F.; McClean, K.L.; Haymond, M.W. A dual-isotope technique for determination of in vivo ketone body kinetics. Am. J. Physiol. 1986, 251, E185–E191. [Google Scholar] [CrossRef]
- Bougneres, P.F.; Ferre, P. Study of ketone body kinetics in children by a combined perfusion of 13C and 2H3 tracers. Am. J. Physiol. 1987, 253, E496–E502. [Google Scholar] [CrossRef]
- Reed, W.D.; Baab, P.J.; Hawkins, R.L.; Ozand, P.T. A double-isotope method for the measurement of ketone-body turnover in the rat. Effect of L-alanine. Biochem. J. 1984, 219, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.M.; Pitt, J.J. Pitfalls in the use of 2-octynoic acid as an in vivo model of medium-chain acyl-coenzyme A dehydrogenase deficiency: Ketone turnover and metabolite studies in the rat. Metab. Clin. Exp. 1999, 48, 685–688. [Google Scholar] [CrossRef]
- Bailey, J.W.; Haymond, M.W.; Miles, J.M. Validation of two-pool model for in vivo ketone body kinetics. Am. J. Physiol. 1990, 258, E850–E855. [Google Scholar] [CrossRef]
- Satapati, S.; Sunny, N.E.; Kucejova, B.; Fu, X.; He, T.T.; Mendez-Lucas, A.; Shelton, J.M.; Perales, J.C.; Browning, J.D.; Burgess, S.C. Elevated TCA cycle function in the pathology of diet-induced hepatic insulin resistance and fatty liver. J. Lipid Res. 2012, 53, 1080–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunny, N.E.; Satapati, S.; Fu, X.; He, T.; Mehdibeigi, R.; Spring-Robinson, C.; Duarte, J.; Potthoff, M.J.; Browning, J.D.; Burgess, S.C. Progressive adaptation of hepatic ketogenesis in mice fed a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1226–E1235. [Google Scholar] [CrossRef] [Green Version]
- Balasse, E.O. Kinetics of ketone body metabolism in fasting humans. Metab. Clin. Exp. 1979, 28, 41–50. [Google Scholar] [CrossRef]
- Mamer, O.A. Measurement of urinary lactic, 3-hydroxybutyric, pyruvic and acetoacetic acids in a single analysis using selected ion monitoring and stable isotope labelling techniques. Biomed. Environ. Mass Spectrom. 1988, 15, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wolfenden, R.; Lewis, C.A., Jr.; Yuan, Y. Kinetic challenges facing oxalate, malonate, acetoacetate, and oxaloacetate decarboxylases. J. Am. Chem. Soc. 2011, 133, 5683–5685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniewicz, M.R.; Kelleher, J.K.; Stephanopoulos, G. Elementary metabolite units (EMU): A novel framework for modeling isotopic distributions. Metab. Eng. 2007, 9, 68–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.J.; Wang, Y.; Cline, G.W.; Rabin-Court, A.; Song, J.D.; Dufour, S.; Zhang, X.M.; Petersen, K.F.; Shulman, G.I. Leptin Mediates a Glucose-Fatty Acid Cycle to Maintain Glucose Homeostasis in Starvation. Cell 2018, 172, 234–248.e217. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Rabin-Court, A.; Song, J.D.; Cardone, R.L.; Wang, Y.; Kibbey, R.G.; Shulman, G.I. Dehydration and insulinopenia are necessary and sufficient for euglycemic ketoacidosis in SGLT2 inhibitor-treated rats. Nat. Commun. 2019, 10, 548. [Google Scholar] [CrossRef] [Green Version]
- Reichard, G.A., Jr.; Haff, A.C.; Skutches, C.L.; Paul, P.; Holroyde, C.P.; Owen, O.E. Plasma Acetone Metabolism in the Fasting Human. J. Clin. Investig. 1979, 63, 619–626. [Google Scholar] [CrossRef]
- Des Rosiers, C.; Montgomery, J.A.; Desrochers, S.; Garneau, M.; David, F.; Mamer, O.A.; Brunengraber, H. Interference of 3-hydroxyisobutyrate with measurements of ketone body concentration and isotopic enrichment by gas chromatography-mass spectrometry. Anal. Biochem. 1988, 173, 96–105. [Google Scholar] [CrossRef]
- Mamer, O.; Gravel, S.P.; Choinière, L.; Chénard, V.; St-Pierre, J.; Avizonis, D. The complete targeted profile of the organic acid intermediates of the citric acid cycle using a single stable isotope dilution analysis, sodium borodeuteride reduction and selected ion monitoring GC/MS. Metabolomics 2013, 9, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Hasenour, C.M.; Wall, M.L.; Ridley, D.E.; Hughey, C.C.; James, F.D.; Wasserman, D.H.; Young, J.D. Mass spectrometry-based microassay of (2)H and (13)C plasma glucose labeling to quantify liver metabolic fluxes in vivo. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E191–E203. [Google Scholar] [CrossRef] [Green Version]
- Hasenour, C.M.; Ridley, D.E.; James, F.D.; Hughey, C.C.; Donahue, E.P.; Viollet, B.; Foretz, M.; Young, J.D.; Wasserman, D.H. Liver AMP-Activated Protein Kinase Is Unnecessary for Gluconeogenesis but Protects Energy State during Nutrient Deprivation. PLoS ONE 2017, 12, e0170382. [Google Scholar] [CrossRef]
- Hughey, C.C.; James, F.D.; Bracy, D.P.; Donahue, E.P.; Young, J.D.; Viollet, B.; Foretz, M.; Wasserman, D.H. Loss of hepatic AMP-activated protein kinase impedes the rate of glycogenolysis but not gluconeogenic fluxes in exercising mice. J. Biol. Chem. 2017, 292, 20125–20140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deja, S.; Fu, X.; Fletcher, J.A.; Kucejova, B.; Browning, J.D.; Young, J.D.; Burgess, S.C. Simultaneous tracers and a unified model of positional and mass isotopomers for quantification of metabolic flux in liver. Metab Eng 2020, 59, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hasenour, C.M.; Rahim, M.; Young, J.D. In Vivo Estimates of Liver Metabolic Flux Assessed by (13)C-Propionate and (13)C-Lactate Are Impacted by Tracer Recycling and Equilibrium Assumptions. Cell Rep. 2020, 32, 107986. [Google Scholar] [CrossRef] [PubMed]
- Metallo, C.M.; Walther, J.L.; Stephanopoulos, G. Evaluation of 13C isotopic tracers for metabolic flux analysis in mammalian cells. J. Biotechnol. 2009, 144, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.H.; Lund, P.; Krebs, H.A. The redox state of free nicotinamide-adenine dinucleotide in the cytoplasm and mitochondria of rat liver. Biochem. J. 1967, 103, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Owen, O.E.; Reichard, G.A., Jr. Human forearm metabolism during progressive starvation. J. Clin. Investig. 1971, 50, 1536–1545. [Google Scholar] [CrossRef] [Green Version]
- Hagenfeldt, L.; Wahren, J. Human Forearm Muscle Metabolism during Exercise III Uptake, release and oxidation of β-hydroxybutyrate and observations on the β-hydroxybutyrate/acetoacetate ratio. Scand. J. Clin. Lab. Investig. 1968, 21, 314–320. [Google Scholar] [CrossRef]
- Hagenfeldt, L.; Wahren, J. Human Forearm Muscle Metabolism during Exercise VI. Substrate Utilization in Prolonged Fasting. Scand. J. Clin. Lab. Investig. 1971, 27, 299–306. [Google Scholar] [CrossRef]
- Aoki, T.T.; Toews, C.J.; Rossini, A.A.; Ruderman, N.B.; Cahill, G.F., Jr. Glucogenic substrate levels in fasting man. Adv. Enzym. Regul. 1975, 13, 329–336. [Google Scholar] [CrossRef]
- Winder, W.W.; Baldwin, K.M.; Holloszy, J.O. Enzymes involved in ketone utilization in different types of muscle: Adaptation to exercise. Eur. J. Biochem. 1974, 47, 461–467. [Google Scholar] [CrossRef]
- Williamson, D.H.; Bates, M.W.; Page, M.A.; Krebs, H.A. Activities of enzymes involved in acetoacetate utilization in adult mammalian tissues. Biochem. J. 1971, 121, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Fink, G.; Desrochers, S.; Des Rosiers, C.; Garneau, M.; David, F.; Daloze, T.; Landau, B.R.; Brunengraber, H. Pseudoketogenesis in the perfused rat heart. J. Biol. Chem. 1988, 263, 18036–18042. [Google Scholar] [CrossRef]
- Des Rosiers, C.; Montgomery, J.A.; Garneau, M.; David, F.; Mamer, O.A.; Daloze, P.; Toffolo, G.; Cobelli, C.; Landau, B.R.; Brunengraber, H. Pseudoketogenesis in hepatectomized dogs. Am. J. Physiol. 1990, 258, E519–E528. [Google Scholar] [CrossRef]
- Kucejova, B.; Duarte, J.; Satapati, S.; Fu, X.; Ilkayeva, O.; Newgard, C.B.; Brugarolas, J.; Burgess, S.C. Hepatic mTORC1 Opposes Impaired Insulin Action to Control Mitochondrial Metabolism in Obesity. Cell Rep. 2016, 16, 508–519. [Google Scholar] [CrossRef] [Green Version]
- Puchalska, P.; Nelson, A.B.; Stagg, D.B.; Crawford, P.A. Determination of ketone bodies in biological samples via rapid UPLC-MS/MS. Talanta 2021, 225, 122048. [Google Scholar] [CrossRef]
- Young, J.D. INCA: A computational platform for isotopically non-stationary metabolic flux analysis. Bioinformatics 2014, 30, 1333–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction | Atom Transitions |
|---|---|
| Rates of tracer infusion | |
| Rinf BHB | BHB.inf (AaBCD) -> BHB.blood (AaBCD) |
| Rinf AcAc | AcAc.inf (ABCD) -> AcAc.blood (ABCD) |
| Rates of appearance | |
| Ra BHB | BHB.source (AaBCD) -> BHB.blood (AaBCD) |
| Ra AcAc | AcAc.source (ABCD) -> AcAc.blood (ABCD) |
| Rates of interconversion | |
| Ri BHB | BHB.blood (AaBCD) -> AcAc.blood (ABCD) + H.bhb (a) |
| Ri AcAc | AcAc.blood (ABCD) + H.h2o (a) -> BHB.blood (AaBCD) |
| Rates of disposal | |
| Rd BHB | BHB.blood (AaBCD) -> BHB.tissue (AaBCD) |
| Rd AcAc | AcAc.blood (ABCD) -> AcAc.tissue (ABCD) |
| Sampling | |
| Rs BHB | 0*BHB.blood (AaBCD) -> BHB.s (AaBCD) |
| Rs AcAc | 0*AcAc.blood (ABCD) + D.h2o (a) -> BHB.s (AaBCD) |
| Sink | BHB.s (AaBCD) -> SINK |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deja, S.; Kucejova, B.; Fu, X.; Browning, J.D.; Young, J.D.; Burgess, S. In Vivo Estimation of Ketogenesis Using Metabolic Flux Analysis—Technical Aspects and Model Interpretation. Metabolites 2021, 11, 279. https://doi.org/10.3390/metabo11050279
Deja S, Kucejova B, Fu X, Browning JD, Young JD, Burgess S. In Vivo Estimation of Ketogenesis Using Metabolic Flux Analysis—Technical Aspects and Model Interpretation. Metabolites. 2021; 11(5):279. https://doi.org/10.3390/metabo11050279
Chicago/Turabian StyleDeja, Stanislaw, Blanka Kucejova, Xiaorong Fu, Jeffrey D. Browning, Jamey D. Young, and Shawn Burgess. 2021. "In Vivo Estimation of Ketogenesis Using Metabolic Flux Analysis—Technical Aspects and Model Interpretation" Metabolites 11, no. 5: 279. https://doi.org/10.3390/metabo11050279
APA StyleDeja, S., Kucejova, B., Fu, X., Browning, J. D., Young, J. D., & Burgess, S. (2021). In Vivo Estimation of Ketogenesis Using Metabolic Flux Analysis—Technical Aspects and Model Interpretation. Metabolites, 11(5), 279. https://doi.org/10.3390/metabo11050279