2.1. Food Intake, Weight Loss, Body Composition, and Global Serum Metabolomics
A complete description of the food intake, BW, body composition, serum chemistry, and fecal microbiota data of cats is available in our previous publication [
28]. Briefly, food intake was significantly lower for wk 1–18 than during the baseline period. Food intake was not different from wk 8 to 18 (47.7 to 44.7 g/d [153.2 to 143.7 kcal ME/d]), but cats continued to lose weight. All cats lost weight and body fat as a result of caloric restriction. Mean BW (7.7 vs. 6.2 kg) and mean BCS (7.6 vs. 6.0) decreased significantly from wk 0 to wk 16. Mean fat mass was significantly lower at wk 8, 12, and 16 (2.4–1.8 kg) than at wk 0 (2.9 kg). Body fat percentage was also significantly lower at wk 8, 12, and 16 (36.8–30.7%) than at wk 0 (40.9%). Mean lean body mass was significantly lower at wk 12 and 16 (3.7 kg) than at wk 0 (3.9 kg). Mean bone mineral content was significantly lower at wk 12 and 16 (92.7 and 92.4 g, respectively) than at wk 0 (108.2 g). Most serum biochemical results remained within the respective reference ranges of the clinical laboratory throughout the study. The exception was the significantly higher creatinine concentrations (reference range, 0.4 to 1.6 mg/dL) from wk 1 (1.74 mg/dL) to 16 (1.91 mg/dL) than the concentration at wk 0 (1.59 mg/dL). Mean triglyceride concentrations were significantly lower at wk 1–16 than the concentration at wk 0 (56.0 mg/dL). Relative abundance of fecal Actinobacteria increased and Bacteroidetes decreased with weight loss. At the genus level,
Blautia,
Dorea,
Eubacterium,
Oscillospira,
Peptococcus, and
Ruminococcus increased with weight loss, while
Lactobacillus,
Butyricicoccus, and
Phascolarctobacterium decreased. Alpha diversity (species richness) and beta diversity were not affected.
A total of 535 named biochemicals were identified, with up to 269 metabolites being altered (
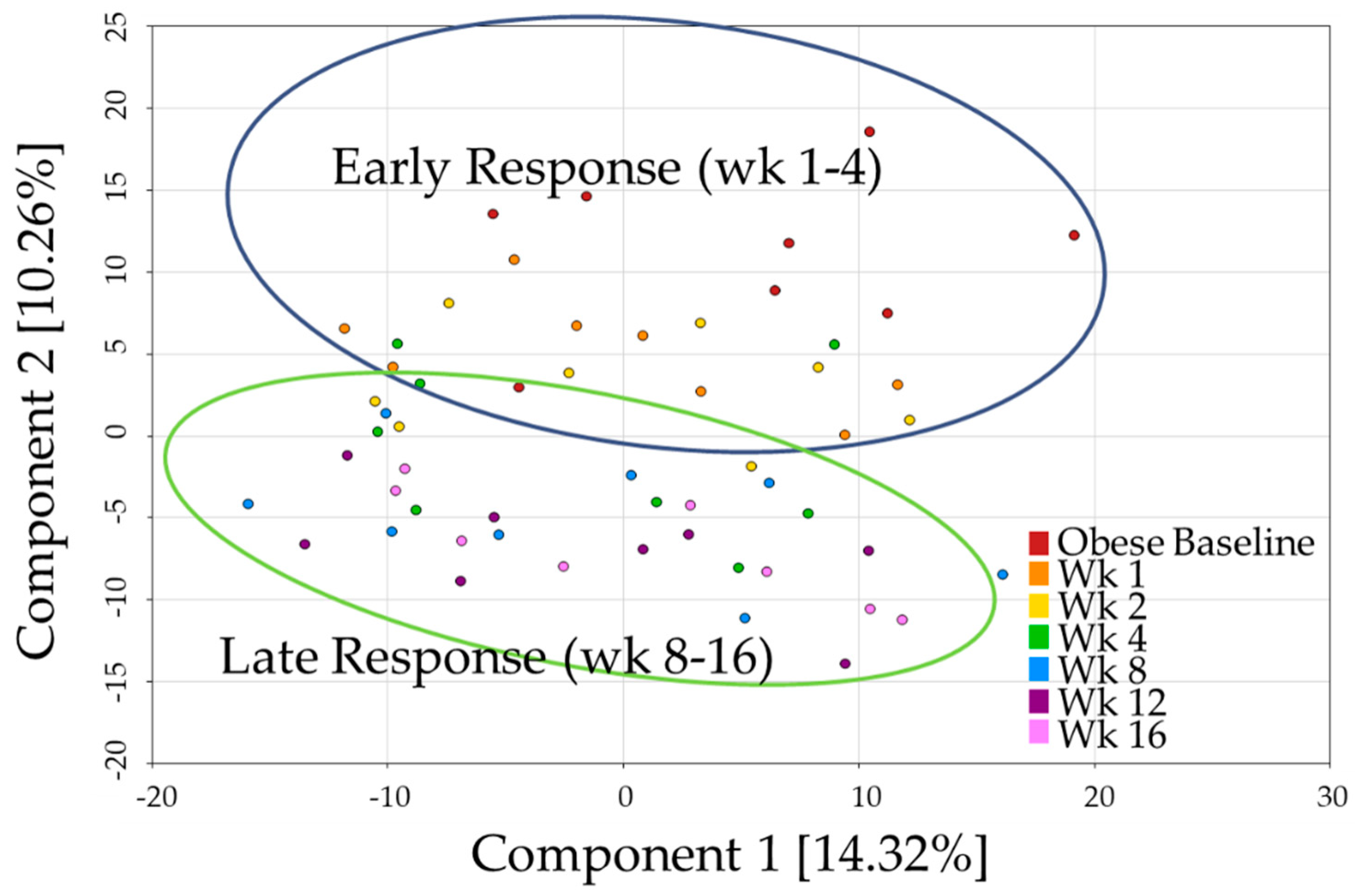
p- and q-values < 0.05) at any time point. Principal component analysis (PCA,
Figure 1) showed a continual shift in metabolite profile as weight loss progressed. Components one and two explained 14.3% and 10.3% of the variability, respectively. Although distinct clusters did not form, a biphasic relationship seemed to distinguish the early (wk 1–4) and late (wk 8–16) responses to weight loss. Broadly speaking, wk 1 and 2 appeared similar to baseline, wk 4 was variable, and wk 8, 12, and 16 were more differentiated from baseline. All of the metabolites discussed below were statistically significant with a combined
p- and q-value ≤ 0.05.
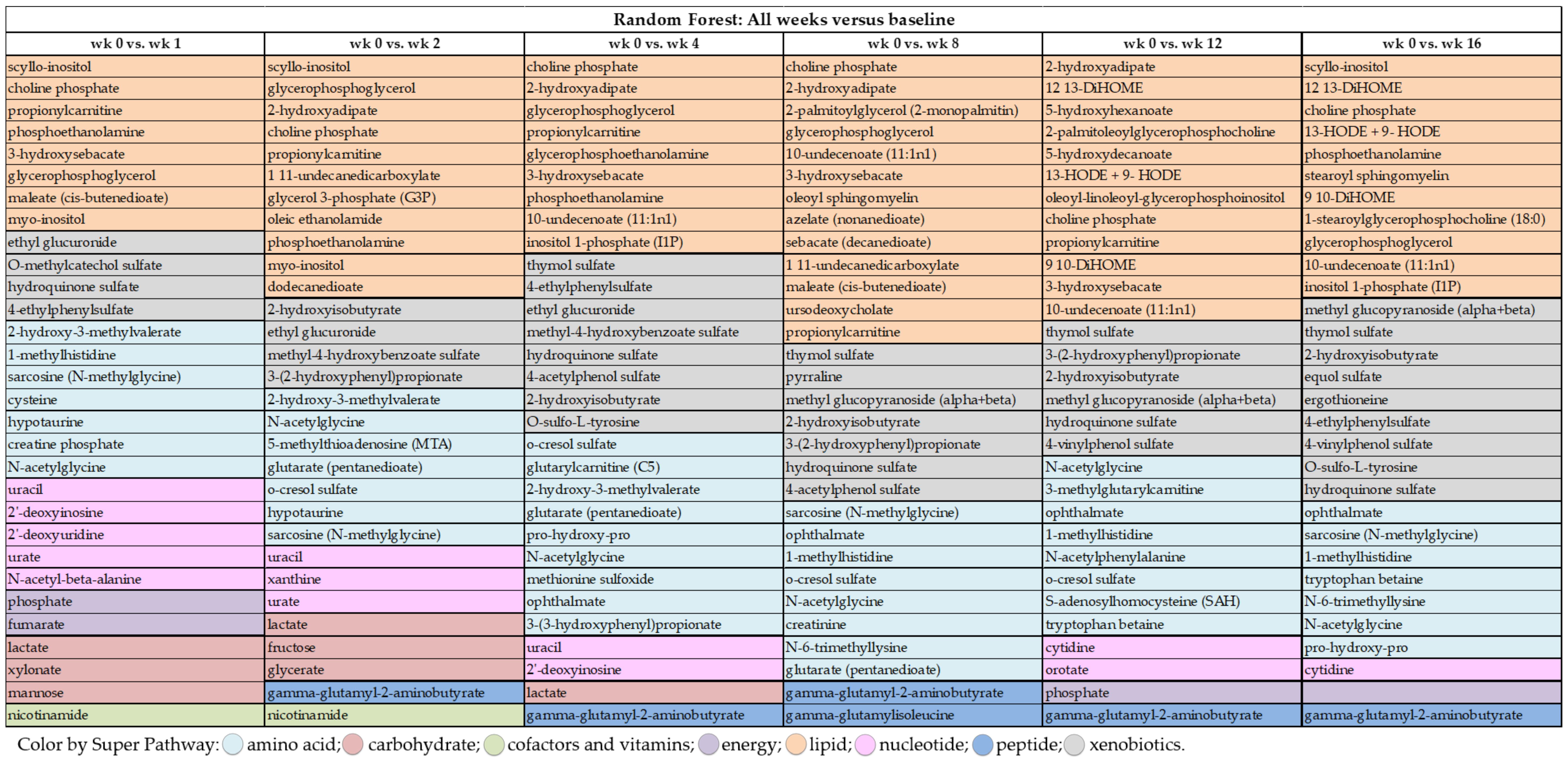
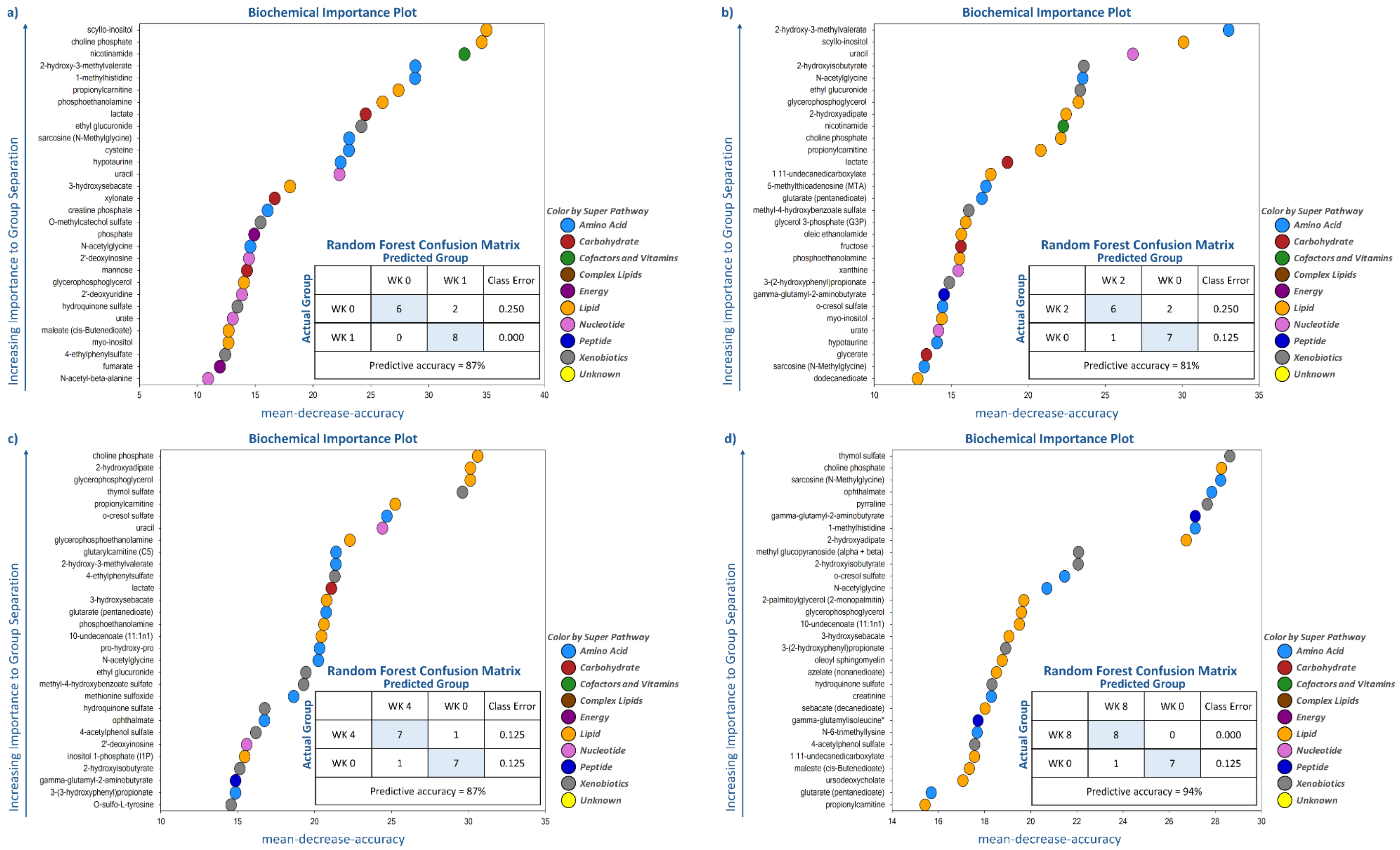
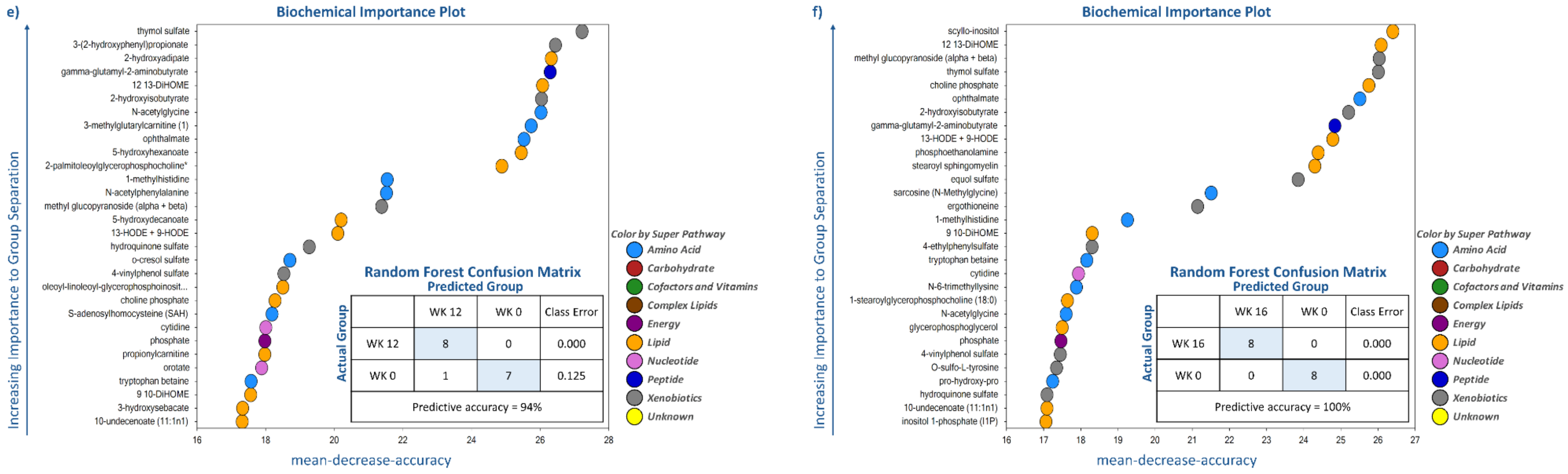
Random forest analysis was performed to identify and rank the top metabolites affected by weight loss by comparing metabolite profiles at wk 0 with the other time points (i.e., wk 1, 2, 4, 8, 12, and 16). As demonstrated by the large mean decrease accuracy (MDA) values in
Figure 2 and
Figure 3,
Supplementary Tables S1–S4, weight loss quickly and dramatically altered metabolite profiles. Even though predictive accuracy improved with greater weight loss over time, accuracy was between 81% and 87% during the initial four wk. Random forest analysis showed that lipid- and amino acid-based metabolites made up 15 to 21 of the top 30 metabolites identified at each time point. Moreover, while some metabolites such as N-acetylglycine (amino acid (AA) metabolism), sarcosine (AA metabolism), choline phosphate (lipid metabolism), and 2-hydroxyisobutyrate (
Figure 2 and
Figure 3) had consistently high MDA values through the entire study, others were indicative of initial (e.g., uracil, lactate, nicotinamide, myo-inositol) or long-term (e.g., thymol sulfate; 1-methylhistidine; 12, 13-dihydroxyoctadecanoic acid (DiHOME); 9, 10-DiHOME) weight loss.
Many of the metabolites with the highest MDA in early weight loss (wk 1–4 vs. wk 0) were related to lipid metabolism (e.g., scyllo-inositol, choline phosphate, propionylcarnitine, phosphoethanolamine,
Figure 2 and
Figure 3). Other metabolites with consistently high MDA values were nicotinamide (cofactors and vitamins), 2-hydroxy-3-methylvalerate (AA metabolism), ethyl glucuronide (xenobiotic metabolism), lactate (carbohydrate metabolism), and uracil (nucleotide metabolism) (
Figure 2 and
Figure 3). Late weight loss (wk 8–16 vs. wk 0) was characterized by altering metabolites associated with lipid (10-undecenoate [11:1n1]; 12, 13-DiHOME; 9, 10-DiHOME), AA (ophthalmate and 1-methylhistidine), and xenobiotic (thymol sulfate and 2-hydroxyisobutyrate) metabolism (
Figure 2 and
Figure 3). One metabolite associated with peptide metabolism, gamma-glutamyl-2-aminobutyrate, had a consistently high MDA from wk 8–16 vs. wk 0 (
Figure 2 and
Figure 3).
2.2. Metabolite Profiles Associated with Lipid Metabolism
Of the 269 metabolites altered with weight loss in this study, over half of them (i.e., 144) were related to lipid metabolism (
Table 1 and
Table 2,
Supplementary Table S1). Metabolites of inositol (myo-inositol, scyllo-inositol, and inositol 1-phosphate,
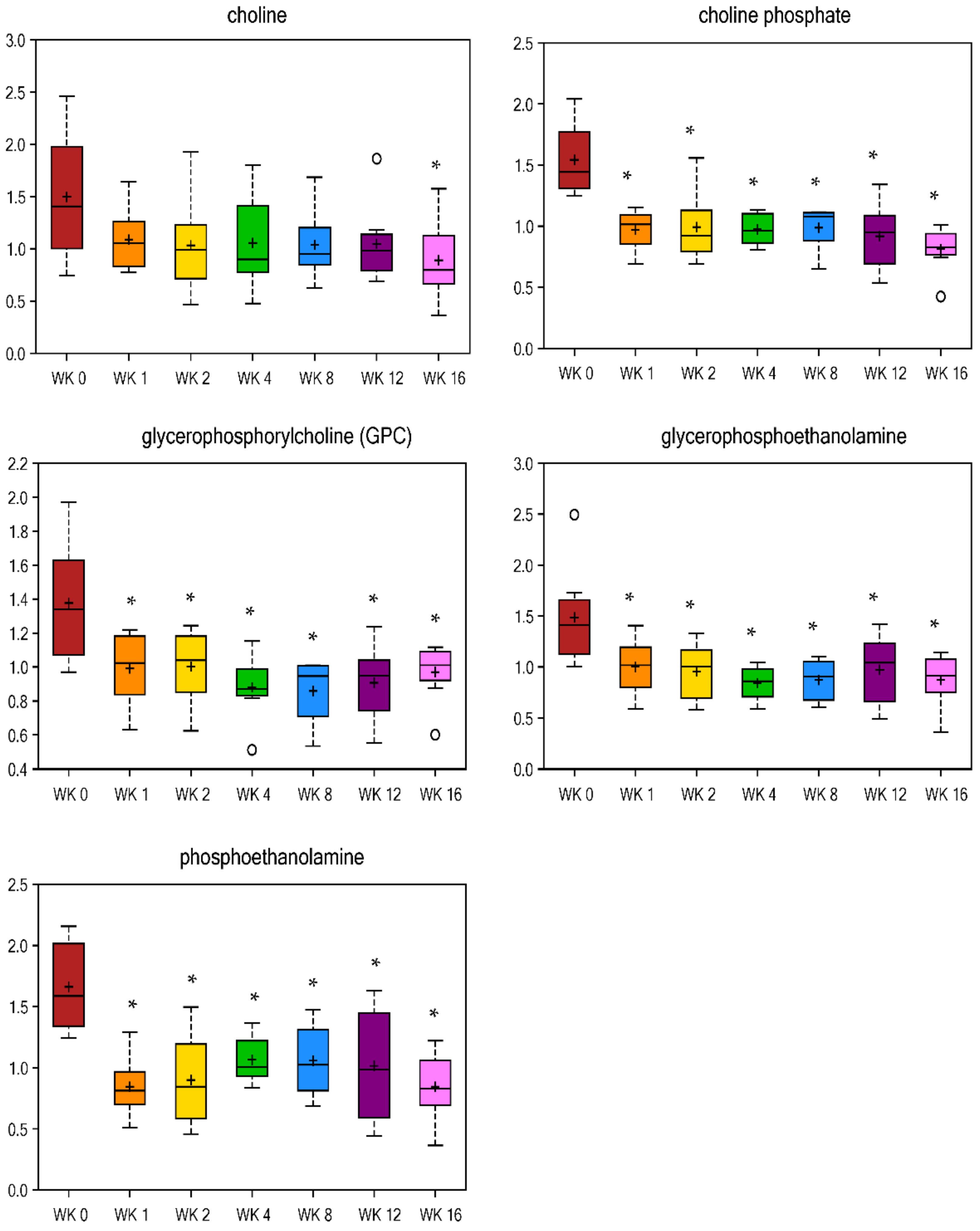
Table 2) and phospholipid metabolism (choline phosphate, phosphoethanolamine, and glycerophosphoethanolamine,
Figure 4) immediately decreased by wk 1 of weight loss and remained lower at all wk vs. wk 0. In humans, abnormalities in myo-inositol metabolism have been associated with insulin resistance and its depletion has been found in tissues affected by diabetic microvascular and neurological complications in animal models and humans [
29], but these metabolites have yet to be studied well in cats. Additionally, increases in myo-inositol may reduce insulin resistance [
30,
31,
32,
33,
34]. When metabolized in the liver, fatty acids (FA) may be oxidized within the mitochondria to produce acetyl-CoA (generate ATP or sterols). FA may also be converted to triacylglycerols and exported as VLDL or stored as lipid droplets. Finally, FA may be metabolized into various phospholipids or sphingolipids [
35]. The conversion of FA to sphingolipid or phospholipid metabolites has been linked to obesity, insulin resistance, type 2 DM, and cardiovascular disease [
35,
36,
37]. Choline plays a role in the cell membrane structure, methyl metabolism, and lipid metabolism. The majority (>95%) of choline is used to synthesize phosphatidylcholine (PC) [
38], which was shown to be greater in high-fat diet (HFD) fed obese mice [
39]. Monoacylglycerols, mainly 2-palmitoylglycerol (wk 1 fold change = 3.05,
Table 1), were increased by wk 1 of weight loss and remained higher throughout weight loss.
Long-chain FA and polyunsaturated FA (PUFA) were lower at wk 8, 12, and 16 vs. wk 0. The medium-chain FA 10-undeconoate (11: 1n1) was increased by wk 1 (fold change = 1.11) and remained higher at all wk vs. wk 0. Blood lipids may be derived from the diet, from adipose tissue, or the liver. These lipids are an important source of energy for the host and are stored primarily as triglycerides in adipose tissue [
39]. Obesity is generally associated with elevated plasma, serum, and liver concentrations of non-esterified FA (NEFA), especially saturated FA (SFA) [
40]. Furthermore, higher concentrations of stearic acid (
p = 0.035), total SFA (
p = 0.051), and palmitoleic acid (
p = 0.068) along with lower linoleic acid (
p = 0.084) concentrations have been reported in obese men [
41]. Another study reported that total SFA (palmitic acid [C16:0] and stearic acid [C18:0]) and monounsaturated FA were lower (
p < 0.005) after eight wk of energy restriction (–15% MER) in obese adults [
42]. Although cats are true carnivores and do not develop cardiovascular disease in the same way that humans do, blood lipid profiles are indicative of their metabolic status and are important in regard to obesity and type 2 DM. Not surprisingly, most long-chain FA in this study were lower at wk 8, 12, and 16 vs. wk 0. Interestingly, monoacylglycerols, which are intermediates of lipolysis, were increased with weight loss.
Markers of primary bile acid metabolism, cholate, and taurocholate (
Table 2), immediately and dramatically decreased with reduced food intake and weight loss, with a fold change of 0.03 and 0.14, respectively, at wk 1. Markers of secondary bile acid metabolism, deoxycholate, and ursodeoxycholate (
Table 2), also decreased with weight loss. Bile acids do not only function to digest lipids in the diet, but are signaling molecules that regulate metabolism and inflammation in obesity, type 2 DM, dyslipidemia, and nonalcoholic fatty liver disease [
43]. In humans, total bile acid concentrations are positively correlated with BMI [
44] and with type 2 DM [
45,
46]. The same relationship may be expected in cats, but has not yet been studied to our knowledge. Mevalonate (
Table 2), the product of rate-limiting enzyme HMG-CoA reductase (HMGR), was significantly lower after wk 8 vs. wk 0 (fold change = 0.65). Cholesterol, the major product of HMGR, remained unchanged. The cholesterol-derived primary and secondary bile acids were greatly decreased with weight loss. Cholate, in particular, was reduced by a fold change of 0.03 and 0.01 at wk 1 and wk 16 vs. wk 0, respectively. Taurocholate and deoxycholate had similar reductions. The bile acid precursor 7-Hoca was essentially unchanged, with an increase only at wk 8 vs. wk 0 (fold change = 1.1), suggesting a reduced need for emulsifying bile acids to aid in lipid digestion and absorption.
Markers of glycerolipid metabolism, glycerol 3-phosphate (G3P), and glycerophosphoglycerol (
Table 2) decreased by wk 1 vs. wk 0 (fold change = 0.59 and 0.72, respectively). Some lysolipids decreased with weight loss, with many changes occurring at wk 4 or later. Many dicarboxylate FA (i.e., 2-hydroxyadipate and 1,11-undecanedicarboxylate,
Table 2) decreased with weight loss. While markers of branched-chain amino acid (BCAA) metabolism (butyrylcarnitine and propionylcarnitine,
Table 2) decreased, those of acylglycine and acylcarnitine metabolism increased throughout weight loss (
Table 1). C3 acylcarnitine is a by-product of isoleucine and valine catabolism, while C5 acylcarnitines are intermediates of mitochondrial isoleucine and leucine catabolism. Both C3 and C5 acylcarnitines have been reported to increase with human obesity [
47]. Furthermore, propionylcarnitine, butyrylcarnitine, and hexanoylcarnitines have been identified as being greater in obese men [
41]. Butyrylcarnitine was lower from wk 2 to 16 vs. wk 0, and propionylcarnitine was lower at all wk vs. wk 0 in the current study.
Carnitine transports FA into the mitochondrion to produce energy via β-oxidation; therefore, carnitine is often used to promote weight loss [
48]. Furthermore, obese mice and humans have displayed a depletion of carnitine in liver tissue [
39,
41]. It may be that lower carnitine in an obese state leads to the insufficient β-oxidation of NEFA, resulting in NEFA being stored as triglycerides in adipose tissue and ultimately, an accumulation of fat [
17]. The majority of FA, including palmitate and stearate (
Table 2), were decreased with weight loss. The ketone bodies acetoacetate and 3-hydroxybutyrate (BHBA) increased throughout the weight-loss period (
Table 1). A study by Schmedes et al. reported an elevated (
p < 0.001) concentration of the ketone bodies, BHBA, and acetoacetate, and a lower (
p < 0.001) concentration of choline, glucose, tyrosine, and lactate in serum of overweight female subjects after a 6-wk very-low-calorie diet (average energy = 617 kcal/d) [
49]. Carbohydrate restriction leads to a reduction in insulin secretion. When circulating insulin concentrations are low, stored fat in adipose tissue undergoes lipolysis [
50,
51]. Once hormone-sensitive lipase is liberated, NEFA in the hepatic mitochondria are preferentially used for β-oxidation to produce acetyl-CoA and ketone bodies, rather than the fats being esterified into triglycerides [
52]. Decreased insulin release promotes a metabolic shift toward lipid oxidation and the utilization of FA and ketones for energy [
53]. This is reflected by an increase in ketone bodies and a decrease in triglycerides in fasting serum samples. Our results in cats agree with these previous reports, with acetoacetate and BHBA increasing up to 1.93 and 2.09 fold, respectively (
Table 1).
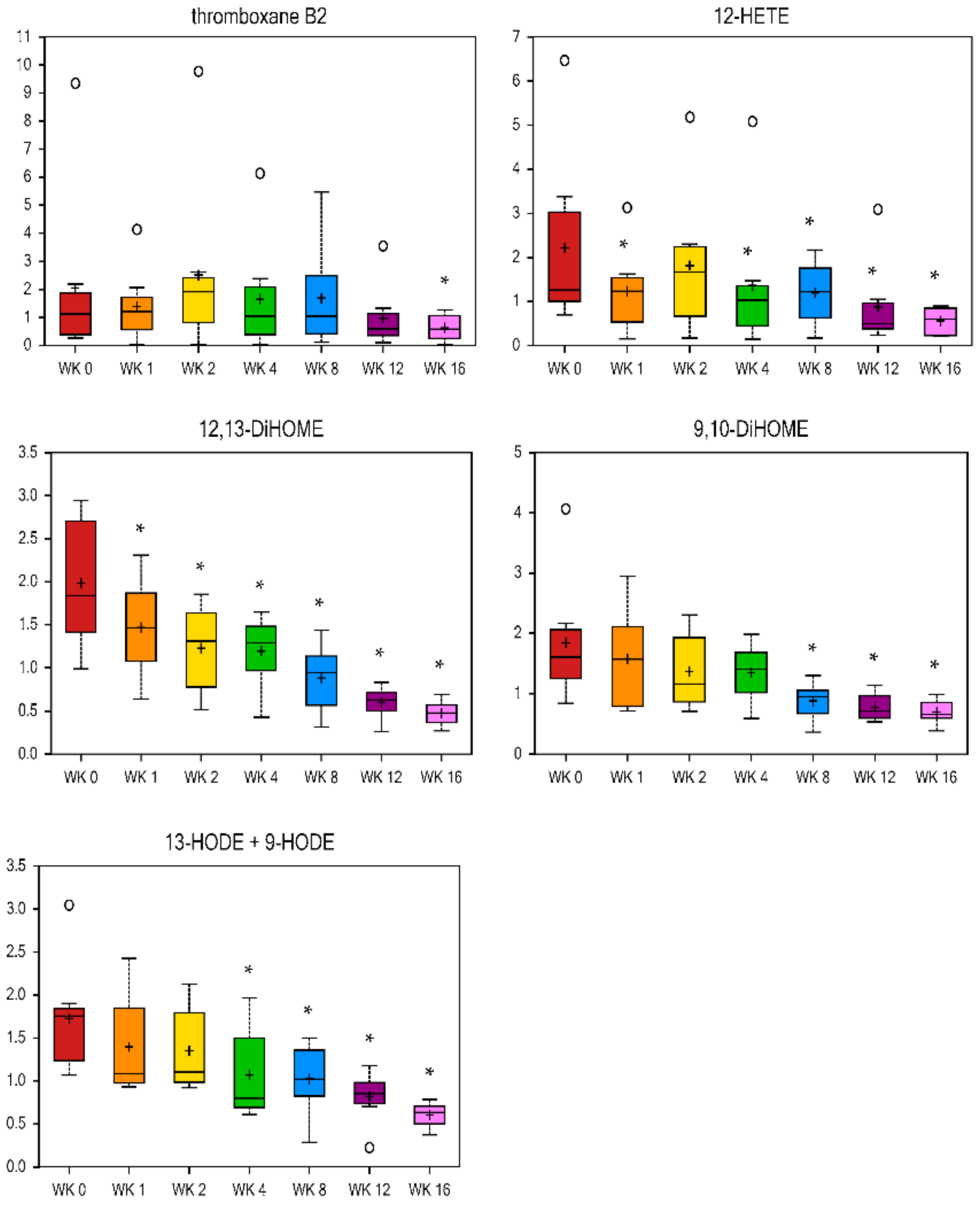
Eicosanoids are oxygenated bioactive metabolites derived from C-20 FA (including arachidonic acid), and include prostaglandins, thromboxanes, leukotrienes, and lipoxins [
54]. They are mediators of acute inflammation, fever, and diseases such as cancer, atherosclerosis, and thrombosis. Therefore, preventing eicosanoid synthesis and action, or modifying the type of eicosanoid to be synthesized is the aim of many drugs. Eicosanoid-related metabolites were decreased with weight loss in the current study. Thromboxane B2 was lower at wk 16 vs. wk 0 (fold change = 0.31), and 12-hydroxyeicosatetraenoic acid (HETE) was reduced more quickly, with a fold change of 0.56 and 0.25 by wk 1 and wk 16 vs. wk 0, respectively (
Figure 5). Triglyceride-rich lipoprotein (TGRL) lipolysis products cause inflammatory stimuli that possibly alter the endothelial barrier function and have pro-atherogenic and pro-inflammatory properties [
55]. Monohydroxy FA (i.e., 3-hydroxysebacate, 5-hydroxyhexanoate, 5-hydroxydecanoate, and 13-HODE + 9-HODE,
Figure 5,
Table 2) and dihydroxy FA (12, 13-DiHOME and 9, 10-DiHOME,
Figure 5) decreased with weight loss. Linoleic acid-derived 13-hydroxyl ocatadecadienoic acid (HODE) and 9-HODE are the major oxidized components of low-density lipoprotein (LDL) and very-low-density lipoprotein (VLDL), respectively [
56,
57]. Other products of linoleic acid oxidation include 12, 13-DiHOME, 9, 10-DiHOME, and epoxy octadecenoic acid (EpOME). Research by Wang et al. reported that significant amounts of these oxidized lipids are released during TGRL lipolysis [
55]. During weight loss in the current study, 12, 13-DiHOME was primarily reduced, with a fold change of 0.74 and 0.24 at wk 1 and wk 16 vs. wk 0, respectively. The metabolite 9, 10-DiHOME was also lower at wk 8, 12, and 16 vs. wk 0, while 13-HODE and 9-HODE were lower from wk 4 to 16 vs. wk 0 of weight loss (
Figure 5). These results in the study of cats agree with the human and rodent literature that obesity is a state of low-grade inflammation and that weight loss can reduce this state.
A correlation analysis identified 17 lipid metabolites from an untargeted analysis that were positively correlated with fasted serum triglyceride concentrations, and 4 lipid metabolites from an untargeted analysis were positively correlated with fasted serum cholesterol concentrations (
Supplementary Table S5). Another 33 lipid metabolites, including ketone bodies (3-hydroxybutyrate and acetoacetate), were positively correlated with body fat mass and fat percentage. Finally, 26 lipid metabolites were positively correlated with BW (
Supplementary Table S6).
2.3. Metabolite Profiles Associated with Amino Acid and Peptide Metabolism
Approximately 40% of the metabolites altered by reduced food intake and weight loss were related to AA metabolism (i.e., 100) and peptide metabolism (i.e., 14) (
Table 3 and
Table 4,
Supplementary Table S2). The sub-pathways of AA metabolism had variable results, containing metabolites that both increased and decreased with weight loss. Most metabolites of lysine metabolism (i.e., N-6-trimethyllysine, glutarylcarnitine, and 3-methylglutarylcarnitine,
Table 3) were higher at wk 1 or 2 vs. wk 0 and remained higher throughout the study. Conversely, glutarate was decreased by wk 1 vs. wk 0 (fold change = 0.75,
Table 4) and remained lower at all wk vs. wk 0. Metabolites of glycine, serine, and threonine metabolism had differing results, with N-acetylglycine being higher at wk 1 vs. wk 0 (fold change = 1.37,
Table 3), and sarcosine being lower (fold change = 0.62,
Table 4). Glutamate was lower at wk 1, 2, and 16 versus wk 0 (fold change = 0.71, 0.73, and 0.67, respectively,
Table 4), while glutamine was only higher at wk 12 vs. wk 0 (fold change = 1.19,
Table 3). In obese children as compared to normal-weight children, glutamine was also increased with weight loss [
58]. In humans, it was also reported that the association between obesity and the activation of the hexosamine pathway, which consumes glutamine upon the formation of glucosamine-6-phosphate from fructose 6-phosphate [
59,
60], and the glucosamine and hexosamines subsequently formed from it, are known to be associated with the development of insulin resistance [
60,
61]. There was an increase in 1-methylhistidine by wk 1 vs. wk 0 (fold change = 1.2,
Table 3), and it remained higher at all wk vs. baseline. Most metabolites of phenylalanine and tyrosine metabolism were decreased with weight loss, with N-acetylphenylalanine being lower at wk 1 vs. wk 0 (fold change = 0.87), o-cresol sulfate being lower at wk 2 vs. wk 0 (fold change = 0.49), and others being lower at wk 4 vs. wk 0 (
Table 4). Similarly, in humans, insulin resistance and an increased risk of developing type 2 DM are associated with elevated concentrations of aromatic AA (tyrosine and phenylalanine) and BCAA (isoleucine, leucine, and valine), with the aromatic AA and BCAA being reported to decrease after weight loss in obese individuals [
47,
62,
63,
64,
65].
Metabolites of BCAA (leucine, isoleucine, and valine) metabolism were inconsistently changed by weight loss, with 2-hydroxy-3-methylvalerate being the only metabolite to decrease by wk 1 vs. wk 0 (fold change = 0.57,
Table 4) and remain lower at all wk. Valine was not changed with weight loss in the current study, but leucine was lower at wk 12 vs. wk 0 (
Table 4). High fasted BCAA and aromatic AA concentrations have been documented in obese humans [
66] and are thought to contribute to obesity-related comorbidities such as insulin resistance and glucose intolerance [
47]. Reportedly, obese men had plasma valine and leucine concentrations that were 23% and 14%, respectively, higher than lean men [
41]. BCAA catabolism may be inhibited with obesity, as obese ob/ob mice and Zucker rats reportedly had depressed activities of BCAA aminotransferase and the branched-chain α-ketoacid dehydrogenase enzyme complex [
67]. Conversely, HFD-fed mice have been shown to have lower serum BCAA concentrations [
68,
69]. In approximately 1300 humans aged 40–79, higher BCAA concentrations were related to older age, male sex, metabolic syndrome, obesity, cardiovascular disease risk, dyslipidemia, hypertension, and uric acid [
70]. Medium- and long-chain acylcarnitines, by-products of the mitochondrial catabolism of BCAA, branched-chain keto acids, and BCAA distinguished obese people with insulin resistance from those without [
71]. In a study of nearly 900 hypertension patients, BCAA, tyrosine, and phenylalanine were associated with metabolic syndrome and impaired fasting glucose [
72]. Finally, elevations in BCAA and alanine were associated with insulin resistance, whereas higher concentrations of glutamine and glycine were associated with a lower likelihood of insulin resistance [
73].
Metabolites of methionine, cysteine, S-adenosylmethionine (SAM), and taurine metabolism mostly decreased with weight loss, and methionine sulfoxide, S-adenosylhomocysteine (SAH), taurine, hypotaurine, and N-acetyltaurine all decreased by wk 1 vs. wk 0 (fold change = 0.73, 0.63, 0.72, 0.31, and 0.86, respectively,
Table 4) and remained lower throughout weight loss. Methionine restriction in rodents reduced circulating lipids, increased metabolic flexibility, enhanced insulin sensitivity, and limited fat deposition by increasing the total daily energy expenditure [
74,
75,
76,
77,
78,
79,
80]. Taurine plays a role in the conjugation of cholesterol and bile acids and has been thought to play a role in obesity [
17]. In a previous study with Labrador Retriever dogs, a decrease in postprandial urinary taurine concentration indicated an alteration in lipid metabolism in overweight dogs, and that taurine may be a possible biomarker [
27]. In humans, plasma total cysteine has a positive relationship with BMI, with higher total cysteine being present in overweight individuals [
81,
82,
83,
84,
85]. Urea cycle metabolites such as urea and citrulline were lower at wk 8, 12, and 16 vs. wk 0 (
Table 4), while pro-hydroxy-pro was higher at wk 2, 4, 8, 12, and 16 vs. wk 0 (
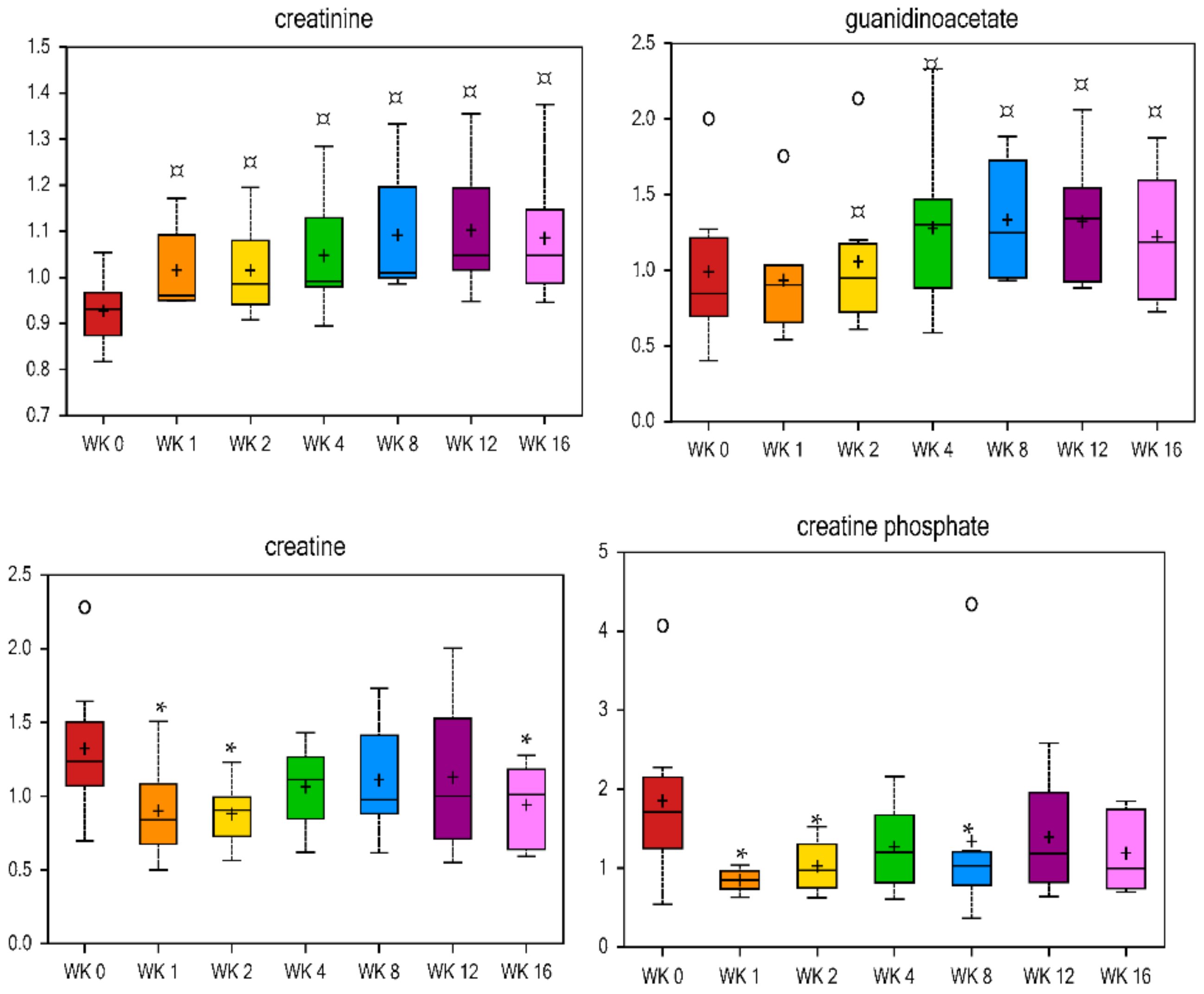
Table 3). Markers of creatine metabolism had opposing results, with both creatine and creatine phosphate being lower at wk 1 and 2 vs. baseline, and creatinine and its precursor guanidinoacetate being higher at wk 1–16 and 4–16 vs. baseline, respectively (
Figure 6).
Glutamine is the most abundant AA in plasma, and glycine is generated from serine, which is derived from pyruvate. Both of these AA, which are precursors of urea biosynthesis and glucose metabolism [
86], were previously reported to be lower (
p < 0.05) in obese individuals [
87,
88]. Protein restriction has been reported to increase glycine and serine concentrations [
89]. Serine, glycine, and threonine metabolic pathways were significantly higher in humans that had Roux-en-Y gastric bypass surgery and sustained weight loss (RYGB-SWL) compared to ones who regained BW (RYGB-WR); most of the statistically different metabolites between the RYGB-SWL and RYGB-WR groups were involved in AA metabolism, one-carbon metabolism, and nucleotide metabolism [
90]. Similarly, in the current study, metabolites of glycine, serine, and threonine were observed to be higher at all wk during weight loss vs. wk 0.
Arginine and glycine synthesize creatine, which is broken down in skeletal muscle to produce creatinine [
91]. Previous research has shown an increase of creatinine in the urine of obese humans (
p < 0.01) [
92] and in the serum of HFD-fed mice (
p < 0.05) [
69]. Ophthalmate, a metabolite of glutathione metabolism, was doubled at wk 4 (fold change = 2.02) in the current study and remained higher with weight loss (
Table 3). In general, most peptide-related metabolites were increased with weight loss. Particularly noteworthy are gamma-glutamylisoleucine, gamma-glutamyl-2-aminobutyrate, N-acetylcarnosine, and prolylglycine, which were higher at wk 1 vs. wk 0 (fold change = 1.23, 1.41, 1.24, and 1.27 respectively) and remained higher at all wk (
Table 3).
A correlation analysis identified five amino acid and one peptide metabolites from an untargeted analysis that were negatively correlated with fasted serum cholesterol concentrations; two amino acid and one peptide metabolites from an untargeted analysis that were negatively correlated with fasted serum triglyceride concentrations; nine amino acid and two peptide metabolites from an untargeted analysis that were positively correlated with fasted serum creatinine concentrations; and eleven amino acid and one peptide metabolites from an untargeted analysis that were positively correlated with fasted serum blood urea nitrogen concentrations (
Supplementary Table S5). Additionally, ten amino acid and one peptide metabolites were negatively correlated with body fat percentage, while six amino acid and one peptide metabolites were negatively correlated with BW (
Supplementary Table S6). The gut microbiome can influence the blood metabolites of the host, including indole-containing metabolites, phenyl derivatives, and flavones [
93]. It was previously reported that enteric bacteria (
Escherichia coli,
Clostridium sporogenes) can convert tryptophan to indoles, and
Bifidobacterium infantis can increase plasma levels of tryptophan [
94,
95,
96]. In the present study, body fat percentage was negatively correlated with two tryptophan-based metabolites, and positively correlated with tryptophan and one phenylalanine/tyrosine-based metabolite (
Supplementary Table S6). These results may be due to substrate availability and/or the alteration of gut bacteria. However, further studies are needed to evaluate the effects of dietary composition, food intake, and weight loss on the relationships among host metabolites and the gut microbiota. An improved understanding may consequently allow for improved dietary interventions for the treatment of obesity in cats.
2.5. Metabolite Profiles Associated with Nucleotide, Xenobiotic, and Cofactor and Vitamin Metabolism
The remaining metabolites altered by reduced food intake and weight loss were related to nucleotide (26 metabolites), xenobiotic (38 metabolites), and cofactor and vitamin (19 metabolites) metabolism (
Table 6 and
Table 7,
Supplementary Table S4). Markers of xanthine- or inosine-containing purine metabolism were decreased, with xanthine, 2′-deoxyinosine, and urate quickly decreasing by wk 1 vs. wk 0 (fold change = 0.48, 0.46, and 0.74, respectively,
Table 7). Some markers of pyrimidine metabolism such as uracil (
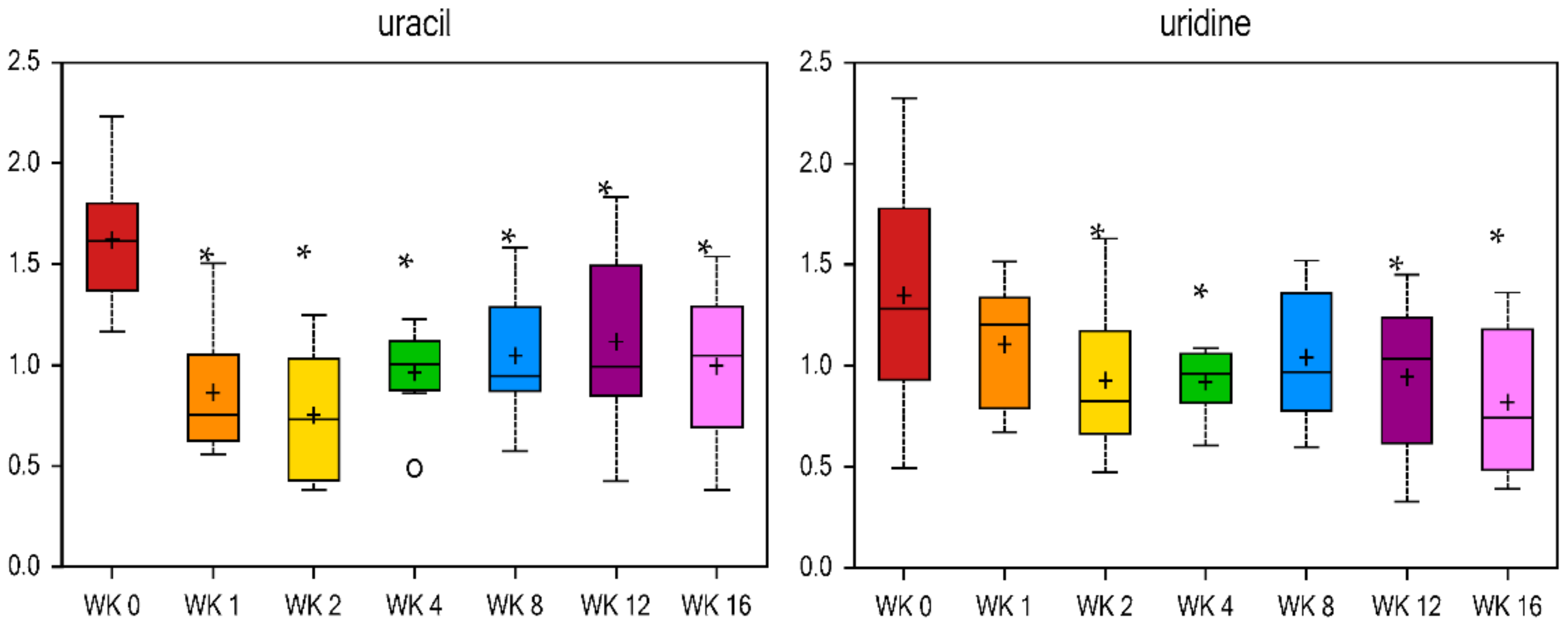
Figure 8), 2′-deoxyuridine, and cytidine were decreased (
Table 7), while others such as orotate were increased (
Table 6) with weight loss. It has been shown that uridine infusion induced insulin resistance in rats [
107] and has been correlated with insulin resistance in hypertensive patients [
108]. Results of the current study show lower uridine at wk 2, 4, 12, and 16 vs. wk 0 (fold change = 0.69, 0.68, 0.7, and 0.61, respectively). Uracil forms uridine when it is combined with a sugar ribose by a glycosidic linkage [
109]. Uracil was lower at all wk vs. wk 0, with a fold change of 0.53 at wk 1 vs. wk 0 (
Figure 8). Most xenobiotics decreased with weight loss. Markers of benzoate metabolism, such as 4-ethylphenylsulfate and 4-vinylphenol sulfate, were lower at wk 1 vs. wk 0 (fold change = 0.41 and 0.49) and continued to decrease with weight loss (
Table 7). Similar results were observed in xenobiotics related to food and plant components (i.e., ergothioneine and pyrraline), drugs (i.e., 4-acetylphenol sulfate and hydroquinone sulfate), and chemicals (i.e., O-sulfo-
l-tyrosine and ethyl glucuronide), with all being decreased with weight loss (
Table 7). Conversely, the chemical-related xenobiotic 2-hydroxyisobutyrate increased with weight loss (fold change = 1.87 at wk 16 vs. wk 0,
Table 6).
Correlation analysis identified five xenobiotic metabolites from an untargeted analysis that were negatively correlated with fasted serum cholesterol concentrations, two xenobiotic and one nucleotide metabolites from an untargeted analysis that were positively correlated with fasted serum triglyceride concentrations, four xenobiotic and one nucleotide metabolites from an untargeted analysis that were positively correlated with fasted serum creatinine concentrations, and fifteen xenobiotic and two nucleotide metabolites from an untargeted analysis that were positively correlated with fasted serum blood urea nitrogen concentrations (
Supplementary Table S5). Likewise, three xenobiotic and one nucleotide metabolites were negatively correlated with body fat percentage, and six xenobiotic and one nucleotide metabolites were negatively correlated with BW (
Supplementary Table S6).
In summary, this study used untargeted metabolomic analyses to identify hundreds of serum metabolites affected by weight loss in cats. Most of the metabolites identified from this study have not been reported previously. Because the cat is a strict carnivore, its metabolism is quite different from that of humans and rodent models. Therefore, these data are expected to serve as a foundation for future studies focused on feline obesity, metabolism, and health. A random forest analysis comparing metabolite profiles across time points demonstrated that lipid- and amino acid-based metabolites made up one-half to two-thirds of the top 30 metabolites most influential in regard to predictive accuracy. Our analyses also show that while some metabolites such as N-acetylglycine (amino acid metabolism), sarcosine (amino acid metabolism), and choline phosphate (lipid metabolism) were highly predictive of weight loss throughout the entire study, other metabolites were indicative of initial (e.g., uracil; lactate; nicotinamide; myo-inositol) or long-term (thymol sulfate; 1-methylhistidine; 12, 13-DiHOME; 9, 10-DiHOME) weight loss. The majority of metabolites associated with lipid metabolism decreased with weight loss, which was likely due to a reduction in food intake (diet acid-hydrolyzed fat = 8.9%, and caloric restriction 33–40% of baseline intake, wk 5–10, respectively). However, ketone bodies and small lipid particles (monoacylglycerol, FA, and medium-chain FA) were increased, indicating that obese cats undergoing weight loss use lipolysis and FA oxidation to produce energy. The majority of metabolites associated with carbohydrate metabolism were decreased with weight loss, which was thought to be due to lower intake. Metabolites associated with protein metabolism had a variable result with weight loss, which may indicate that cats are in a state of constant flux in regard to muscle mass loss and synthesis during energy deficit. Finally, metabolic mediators of inflammation, oxidative stress markers, xenobiotics, and biomarkers of insulin resistance were decreased with weight loss. This dataset provides an improved understanding of feline metabolism, with a specific focus on how it is impacted by reduced food intake and weight loss. Not only did this study identify biomarkers of weight loss in general, but many that are indicative of the early or late phases of weight loss, which may serve as a foundation for future research using targeted analysis. Such research may lead to the development of metabolite signature panels with an application in veterinary practice, whereby biomarkers may help diagnose disease and aid in the lifestyle, nutritional, and pharmaceutical management of obese cats. Targeted metabolite panels may also be used to develop and test therapeutic diets intended to improve metabolism and reduce clinical signs of obese cats.



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}