
Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Results and Discussion
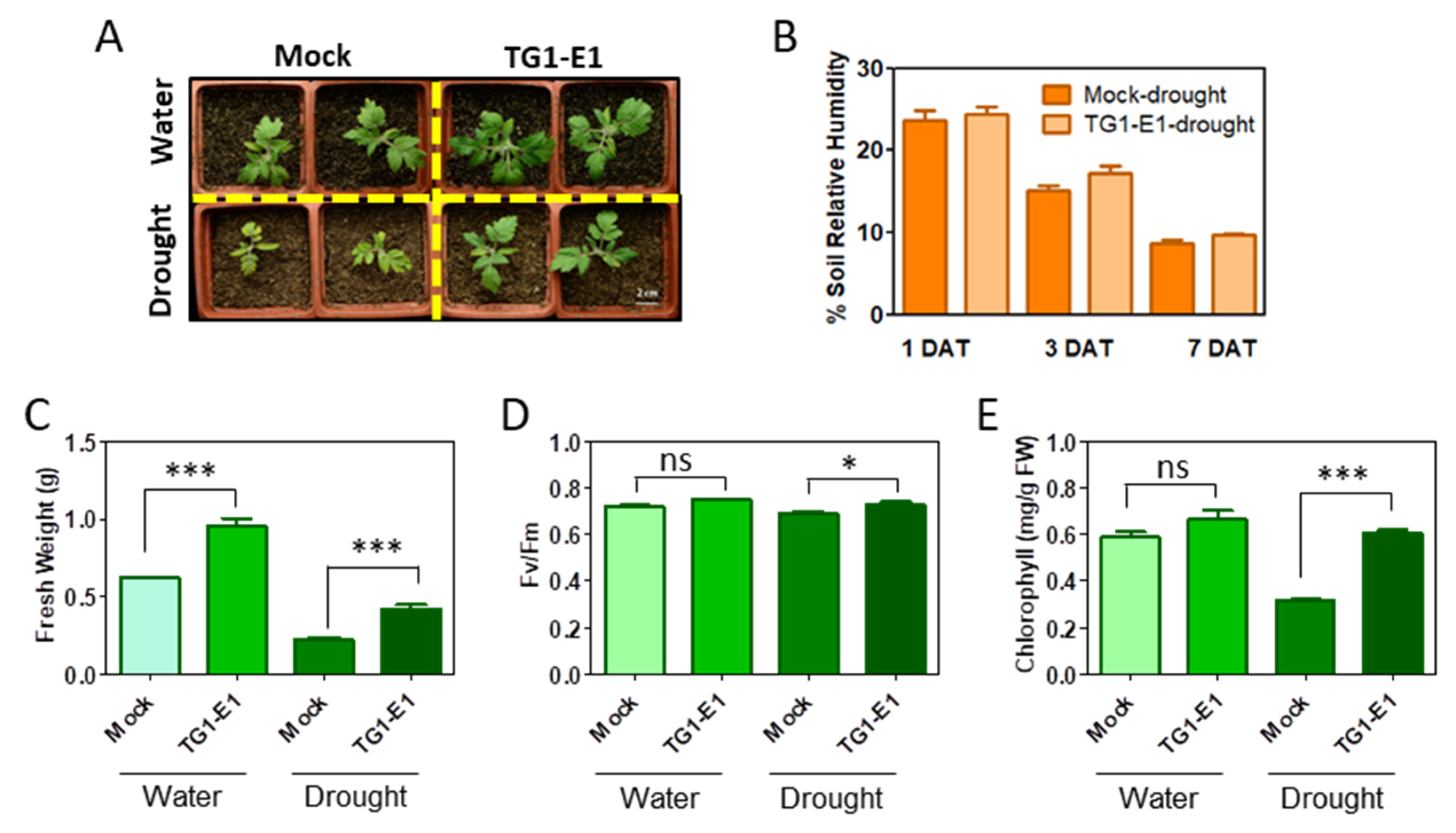
2.1. Bacillus Megaterium TG1-E1 Increases Drought Resistance in Tomato Seedlings
2.2. Bacillus Megaterium TG1-E1 Induces Transcriptomic Reprograming of in Tomato Plants under Drought Stress
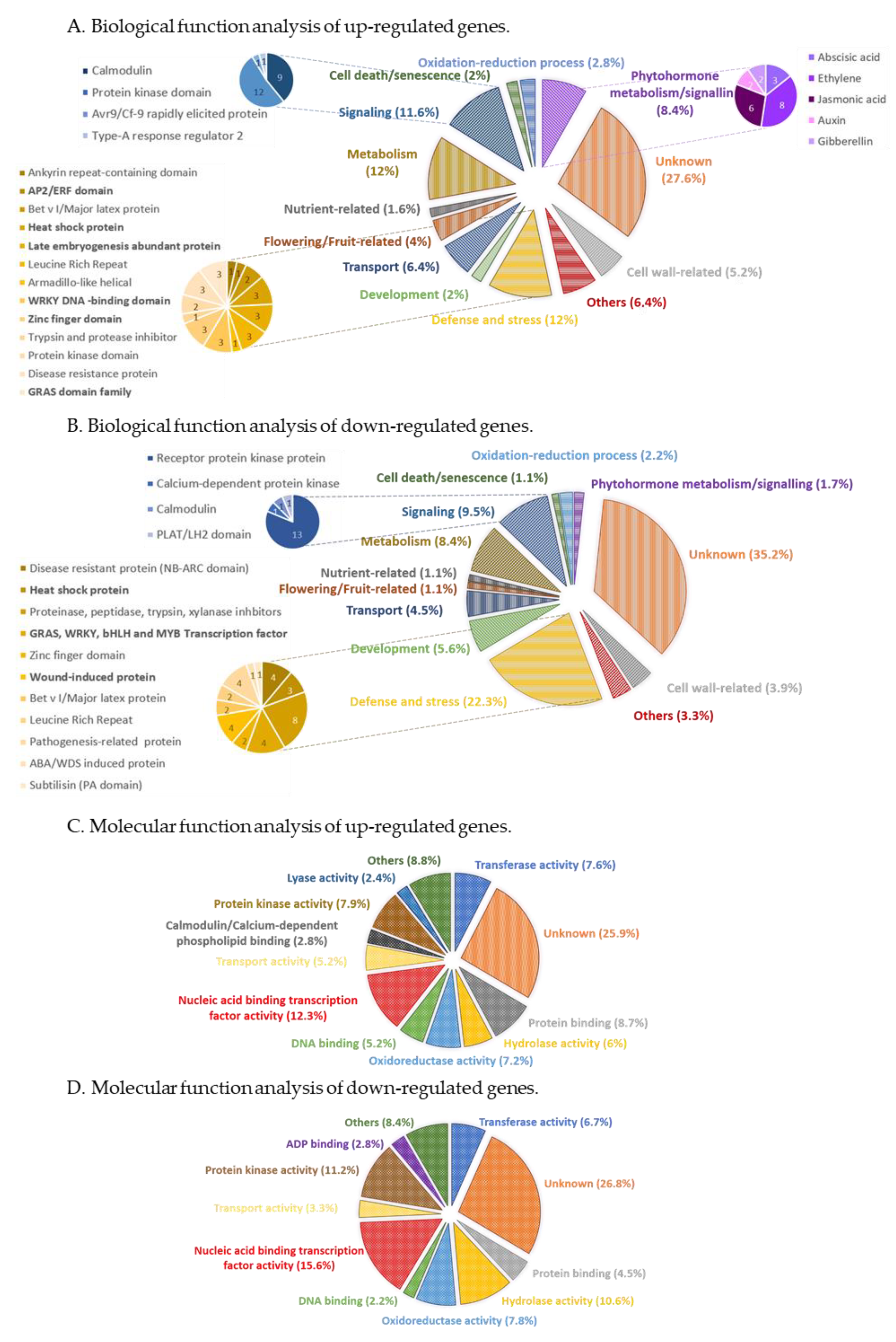
2.2.1. The Overall Functional Categorization of Differentially Expressed Genes (DEGs)
2.2.2. DEGs Implicating Regulation Mediated through Transcription Factors
2.2.3. DEGs Implicating Regulation of Drought-Responsive Signaling
2.2.4. DEGs Implicating Regulation of Cell Wall Biosynthesis and Organization
3. Hyperosmotic Stress Induces TG1-E1 Production of Extracellular Metabolites Including Potential Osmoprotectants
3.1. Sugars
3.2. Amino Acids
3.3. Precursors of the Osmoprotectant Glycine Betaine
3.4. Pinitol
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Bacterial Culture and Inoculation
4.3. Monitoring of Plant Growth
4.4. Bacteria Root Colonization
4.5. Quantification of Chlorophyll Contents and Photosynthesis Efficiency
4.6. Bacterial Extracellular Metabolite Collection and GC-MS Analysis
4.7. RNAseq Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Overview of plant growth-promoting bacteria. In Biochemical and Genetic Mechanisms Used by Plant Growth Promoting Bacteria; Patten, B.R.N., Holguin, G., Penrose, D.M., Eds.; Imperial College Press: London, UK, 1999; pp. 1–13. [Google Scholar]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.M.; Zhang, H. The effects of bacterial volatile emissions on plant abiotic stress resistance. Front. Plant. Sci. 2015, 6, 774. [Google Scholar] [CrossRef]
- Amellal, N.; Burtin, G.; Bartoli, F.; Heulin, T. Colonization of wheat rhizosphere by EPS producing Pantoea agglomerans and its effect on soil aggregation. Appl. Environ. Microbiol. 1998, 64, 3740–3747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Sun, Y.; Xie, X.; Kim, M.S.; Dowd, S.E.; Paré, P.W. A soil iron via deficiency-inducible mechanisms. Plant J. 2009, 58, 568–577. [Google Scholar] [CrossRef]
- Morcillo, R.J.L.; Singh, S.K.; He, D.; An, G.; Vílchez, J.I.; Tang, K.; Yuan, F.; Sun, Y.; Shao, C.; Zhang, S.; et al. Rhizobacterium-derived diacetyl modulates plant immunity in a phosphate-dependent manner. EMBO J. 2019, 39, e102602. [Google Scholar] [CrossRef]
- Vílchez, J.I.; Tang, Q.; Kaushal, R.; Wang, W.; Lv, S.; He, D.; Chu, Z.; Zhang, H.; Liu, R.; Zhang, H. Complete Genome Sequence of Bacillus megaterium Strain TG1-E1, a Plant Drought Resistance-Enhancing Bacterium. Microbiol. Resour. Announc. 2018, 7, e00842-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and resistance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genom. 2012, 287, 495–513. [Google Scholar] [CrossRef] [PubMed]
- Karkute, S.G.; Gujjar, R.S.; Rai, A.; Akhtar, M.; Singh, M.; Singh, B. Genome wide expression analysis of WRKY genes in tomato (Solanum lycopersicum) under drought stress. Plant Gene 2018, 13, 8–17. [Google Scholar] [CrossRef]
- Niu, Y.; Zhao, T.; Xu, X.; Li, J. Genome-wide identification and characterization of GRAS transcription factors in tomato (Solanum lycopersicum). PeerJ. 2017, 8, e3955. [Google Scholar] [CrossRef] [Green Version]
- Castelán-Muñoz, N.; Herrera, J.; Cajero-Sánchez, W.; Arrizubieta, M.; Trejo, C.; García-Ponce, B.; Sánchez, M.; Álvarez-Buylla, E.R.; Garay-Arroyo, A. MADS-Box Genes Are Key Components of Genetic Regulatory Networks Involved in Abiotic Stress and Plastic Developmental Responses in Plants. Front. Plant Sci. 2019, 10, 853. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, J.; Pan, C.; Guan, X.; Wang, Y.; Liu, S.; He, Y.; Chen, J.; Chen, L.; Lu, G. Genome-wide identification of MAPKK and MAPKKK gene families in tomato and transcriptional profiling analysis during development and stress response. PLoS ONE 2014, 18, e103032. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Wan, D.; Li, R.; Zou, B.; Zhang, X.; Cong, J.; Wang, R.; Xia, Y.; Li, G. Calmodulin-binding protein CBP60g is a positive regulator of both disease resistance and drought resistance in Arabidopsis. Plant Cell Rep. 2012, 31, 1269–1281. [Google Scholar] [CrossRef]
- Perez-Prat, E.; Narashimhan, M.L.; Binzel, M.L.; Botella, M.A.; Chen, Z.; Valpuesta, V.; Bressan, R.A.; Hasegawa, P.M. Induction of aputative Ca2+-ATPase mRNA in NaCl adapted cells. Plant Physiol. 1992, 100, 1471–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimmers, L.E.; Ewing, N.N.; Bennett, A.B. Higher plant Ca2+ATPase: Primary structure and regulation of mRNA abundance by salt. Proc. Natl. Acad. Sci. USA 1992, 89, 9205–9209. [Google Scholar] [CrossRef] [Green Version]
- Huda, K.M.; Banu, M.S.; Garg, B.; Tula, S.; Tuteja, R.; Tuteja, N. OsACA6, a P-type IIB Ca²⁺ ATPase promotes salinity and drought stress resistance in tobacco by ROS scavenging and enhancing the expression of stress-responsive genes. Plant J. 2013, 76, 997–1015. [Google Scholar] [CrossRef] [PubMed]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [Green Version]
- Rekha, K.; Baskar, B.; Srinath, S.; Usha, B. Plant-growth-promoting rhizobacteria Bacillus subtilis RR4 isolated from rice rhizosphere induces malic acid biosynthesis in rice roots. Can. J. Microbiol. 2018, 64, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 7, 771. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 16, 112. [Google Scholar] [CrossRef]
- Lakshmanan, V.; Castaneda, R.; Rudrappa, T.; Bais, H.P. Root transcriptome analysis of Arabidopsis thaliana exposed to beneficial Bacillus subtilis FB17 rhizobacteria revealed genes for bacterial recruitment and plant defense independent of malate efflux. Planta 2013, 238, 657–668. [Google Scholar] [CrossRef]
- Piro, G.; Leucci, M.R.; Waldron, K.; Dalessandro, G. Exposure to water stress causes changes in the biosynthesis of cell wall polysaccharides in roots of wheat cultivars varying in drought resistance. Plant Sci. 2003, 165, 559–569. [Google Scholar] [CrossRef]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Li, J.; Pan, Y.; Zhang, Y.; Ni, L.; Wang, Y.; Zhang, X. Genome-wide identification and expression analysis of the UGlcAE gene family in tomato. Int. J. Mol. Sci. 2018, 19, 1583. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, Y.; Kato, K.; Ogawa-Ohnishi, M.; Tsuruhama, K.; Kajiura, H.; Yagyu, K.; Takeda, A.; Takeda, Y.; Kunieda, T.; Hara-Nishimura, I.; et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nat. Plants 2018, 4, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Teng, Q.; Zhong, R.; Yuan, Y.; Haghighat, M.; Ye, Z.H. Three Arabidopsis DUF579 domain-containing GXM proteins are methyltransferases catalyzing 4-o-methylation of glucuronic acid on xylan. Plant Cell Physiol. 2012, 53, 1934–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, P.K.; Biswas, R.; Anjum, N.; Das, A.K.; Maiti, M.K. Rice matrix metalloproteinase OsMMP1 plays pleiotropic roles in plant development and symplastic-apoplastic transport by modulating cellulose and callose depositions. Sci. Rep. 2018, 8, 2783. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; García-Fontana, C.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant drought resistance enhancement by trehalose production of desiccation-tolerant microorganisms. Front. Microbiol. 2016, 7, 1577. [Google Scholar] [CrossRef] [Green Version]
- Iordachescu, M.; Imai, R. Trehalose biosynthesis in response to abiotic stresses. J. Integr. Plant Biol. 2008, 50, 1223–1229. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Effect of trehalose on protein structure. Protein Sci. 2009, 18, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, G.L.; Chi, Z.; Hu, Z.; Chi, Z.M. Genetics of trehalose biosynthesis in desert-derived Aureobasidium melanogenum and role of trehalose in the adaptation of the yeast to extreme environments. Curr. Genet. 2018, 64, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Chenu, C.; Roberson, E.B. Diffusion of glucose in microbial extracellular polysaccharide as affected by water potential. Soil Biol. Biochem. 1996, 28, 877–884. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z. The production of exopolysaccharide by Pseudomonas putida GAP-P45 under various abiotic stress conditions and its role in soil aggregation. Microbiology 2015, 84, 512–519. [Google Scholar] [CrossRef]
- Vardharajula, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- ElSayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: Protection against abiotic stress. Plant Biol. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef]
- Takahashi, T.; Kakehi, J.I. Polyamines: Ubiquitous polycations with unique roles in growth and stress responses. Ann. Bot. 2010, 105, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chan, Z. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J. Integr. Plant Biol. 2014, 56, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Sen, S.; Mohapatra, S. Modulation of proline metabolic gene expression in Arabidopsis thaliana under water-stressed conditions by a drought-mitigating Pseudomonas putida strain. Ann. Microbiol. 2017, 67, 655–668. [Google Scholar] [CrossRef]
- Sen, S.; Ghosh, D.; Mohapatra, S. Modulation of polyamine biosynthesis in Arabidopsis thaliana by a drought mitigating Pseudomonas putida strain. Plant Physiol. Biochem. 2018, 129, 180–188. [Google Scholar] [CrossRef]
- Rastogi, R.; Dulson, J.; Rothstein, S.J. Cloning of tomato (Lycopersicon esculenfum Mill.) Arginine decarboxylase gene and its expression during fruit ripening. Plant Physiol. 1993, 103, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Igarashi, Y.; Seki, M.; Sekiguchi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of Arabidopsis genes involved in biosynthesis of polyamines in abiotic stress responses and developmental stages. Plant. Cell Environ. 2003, 26, 1917–1926. [Google Scholar] [CrossRef]
- Kakkar, R.K.; Sawhney, V.K. Polyamine research in plants—A changing perspective. Physiol. Plant. 2002, 116, 281–288. [Google Scholar] [CrossRef]
- Sinha, R.; Rajam, M.V. RNAi silencing of three homologues of S-adenosylmethionine decarboxylase gene in tapetal tissue of tomato results in male sterility. Plant Mol. Biol. 2013, 82, 169–180. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought resistance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef]
- Kan, C.C.; Chung, T.Y.; Juo, Y.A.; Hsieh, M.H. Glutamine rapidly induces the expression of key transcription factor genes involved in nitrogen and stress responses in rice roots. BMC Genom. 2015, 16, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vílchez, J.I.; Niehaus, K.; Dowling, D.N.; González-López, J.; Manzanera, M. Protection of pepper plants from drought by Microbacterium sp. 3J1 by modulation of the plant’s glutamine and a-ketoglutarate content: A comparative metabolomics approach. Front. Microbiol. 2018, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. The glycerate and phosphorylated pathways of serine synthesis in plants: The branches of plant glycolysis linking carbon and nitrogen metabolism. Front. Plant Sci. 2018, 9, 318. [Google Scholar] [CrossRef] [Green Version]
- Rajaeian, S.; Ehsanpour, A.A.; Javadi, M.; Shojaee, B. Ethanolamine induced modification in glycine betaine and proline metabolism in Nicotiana rustica under salt stress. Biol. Plant. 2017, 61, 797–800. [Google Scholar] [CrossRef]
- Bhuiyan, N.H.; Hamada, A.; Yamada, N.; Rai, V.; Hibino, T.; Takabe, T. Regulation of betaine synthesis by precursor supply and choline monooxygenase expression in Amaranthus tricolor. J. Exp. Bot. 2007, 58, 4203–4212. [Google Scholar] [CrossRef]
- Streeter, J.G.; Lohnes, D.G.; Fioritto, R.J. Patterns of pinitol accumulation in soybean plants and relationships to drought resistance. Plant Cell Environ. 2001, 24, 429–438. [Google Scholar] [CrossRef]
- Ahn, C.H.; Hossain, M.D.; Lee, E.; Kanth, B.K.; Park, P.B. Increased salt and drought resistance by D-pinitol production in transgenic Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2018, 504, 315–320. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Yang, Y.; Yi, D.; Zhang, H. Measurements of root colonized bacteria species. Bio-Protoc. 2021, 11, e3976. [Google Scholar]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Gene ID | Annotation | Type of Nucleic Acid Binding Transcription Factor Activity | LogFC |
|---|---|---|---|
| Solyc07g061760.2 | Ankyrin repeats | Ankyrin repeat-containing domain | 2.29 |
| Solyc08g014570.3 | PGG domain | Ankyrin repeat-containing domain | 1.06 |
| Solyc06g076050.3 | Ankyrin repeats | Ankyrin repeat-containing domain | 1.04 |
| Solyc05g052570.3 | Ankyrin repeats | Ankyrin repeat-containing domain | 1.11 |
| Solyc05g013540.1 | AP2 domain | AP2/ERF domain | 1.07 |
| Solyc02g090770.1 | AP2 domain | AP2/ERF domain | 2.79 |
| Solyc08g080290.3 | AP2 domain | AP2/ERF domain | 2.58 |
| Solyc08g007830.1 | AP2 domain | AP2/ERF domain | 4.48 |
| Solyc06g035700.1 | AP2 domain | AP2/ERF domain | 3.41 |
| Solyc08g007820.1 | AP2 domain | AP2/ERF domain | 3.02 |
| Solyc10g050970.1 | AP2 domain | AP2/ERF domain | 3.01 |
| Solyc01g108240.3 | AP2 domain | AP2/ERF domain | 4.22 |
| Solyc10g086530.1 | GRAS domain family | GRAS domain | 1.10 |
| Solyc11g012510.2 | GRAS domain family | GRAS domain | 1.28 |
| Solyc12g005340.2 | GRAS domain family | GRAS domain | 1.77 |
| Solyc08g080540.3 | HSF-type DNA-binding | HSF-type DNA binding | 1.64 |
| Solyc02g089200.3 | SRF-type transcription factor | MADS-box | 1.95 |
| Solyc05g015750.3 | SRF-type transcription factor | MADS-box | 1.67 |
| Solyc01g093960.3 | SRF-type transcription factor | MADS-box | 1.04 |
| Solyc06g059970.3 | SRF-type transcription factor | MADS-box | 1.03 |
| Solyc02g071730.3 | SRF-type transcription factor | MADS-box | 1.09 |
| Solyc08g067230.3 | K-box region | MADS-box/K-box | 2.25 |
| Solyc04g081000.3 | K-box region | MADS-box/K-box | 1.42 |
| Solyc05g015840.3 | SBP domain | SBP-box | 1.17 |
| Solyc10g011910.3 | WRKY DNA -binding domain | Zinc finger domain | 1.84 |
| Solyc03g121400.1 | Dof domain, zinc finger | Zinc finger domain | 1.10 |
| Solyc06g053640.1 | Ring finger domain | Zinc finger domain | 1.85 |
| Solyc10g009550.3 | WRKY DNA -binding domain | Zinc finger domain | 1.40 |
| Solyc08g008280.3 | WRKY DNA -binding domain | Zinc finger domain | 1.28 |
| Solyc04g015360.3 | GATA zinc finger | Zinc finger domain | 1.04 |
| Solyc06g075780.2 | C2H2-type zinc finger | Zinc finger domain | 1.49 |
| Solyc05g051200.1 | AP2 domain | AP2/ERF domain | −1.41 |
| Solyc04g007000.1 | AP2 domain | AP2/ERF domain | −1.80 |
| Solyc03g119390.3 | Helix-loop-helix DNA-binding domain | bHLH domain | −1.55 |
| Solyc03g034000.3 | Helix-loop-helix DNA-binding domain | bHLH domain | −1.53 |
| Solyc12g049320.2 | GRAS domain family | GRAS domain | −1.34 |
| Solyc02g077590.1 | Homeobox domain | HD-Zip domain | −1.34 |
| Solyc05g048830.3 | Myb-like DNA-binding domain | Myb domain | −1.05 |
| Solyc05g053330.3 | Myb-like DNA-binding domain | Myb domain | −1.12 |
| Solyc12g089190.1 | Myb-like DNA-binding domain | Myb domain | −1.27 |
| Solyc12g008800.2 | Myb-like DNA-binding domain | Myb domain | −1.42 |
| Solyc04g076220.3 | Domain of unknown function | PPC domain | −1.30 |
| Solyc05g006340.3 | WD domain, G-beta repeat | WD domain | −1.16 |
| Solyc11g006450.2 | Ring finger domain | Zinc finger domain | −1.22 |
| Solyc05g054650.1 | C2H2-type zinc finger | Zinc finger domain | −1.24 |
| Solyc05g051860.3 | zinc-finger of the FCS-type, C2-C2 | Zinc finger domain | −1.33 |
| Solyc05g015850.3 | WRKY DNA -binding domain | Zinc finger domain | −1.48 |
| Solyc06g008020.3 | HIT zinc finger | Zinc finger domain | −1.15 |
| Solyc00g136260.1 | Ring finger domain | Zinc finger domain | −1.97 |
| Solyc12g008830.2 | GATA zinc finger | Zinc finger domain | −1.44 |
| Solyc03g032060.1 | Ring finger domain | Zinc finger domain | −1.11 |
| Solyc12g006230.2 | Ring finger domain | Zinc finger domain | −1.77 |
| Solyc08g067360.3 | WRKY DNA -binding domain | Zinc finger domain | −1.39 |
| Solyc07g045180.3 | B-box zinc finger | Zinc finger domain | −1.23 |
| Solyc08g082680.3 | Ring finger domain | Zinc finger domain | −1.00 |
| Solyc08g067970.3 | zinc-finger of the FCS-type, C2-C2 | Zinc finger domain | −1.11 |
| Solyc06g061240.3 | PLATZ transcription factor | Zinc finger domain | −1.08 |
| Solyc08g065940.3 | Zinc finger C-x8-C-x5-C-x3-H type | Zinc finger domain | −1.08 |
| Gene ID | Annotation | Type of Signaling Protein | LogFC |
|---|---|---|---|
| Solyc11g072930.2 | Carbohydrate-binding protein of the ER | RPK | 1.74 |
| Solyc06g062450.3 | Carbohydrate-binding protein of the ER | RPK | 1.44 |
| Solyc02g094010.2 | Protein kinase domain | RPK | 1.58 |
| Solyc09g014590.3 | Leucine Rich Repeat | RPK | 3.12 |
| Solyc10g006690.3 | Protein tyrosine kinase | RPK | 1.01 |
| Solyc06g007190.3 | Protein phosphatase 2C | ILKAP | 2.04 |
| Solyc12g009550.1 | Leucine rich repeat | RPK | 1.07 |
| Solyc01g005730.3 | Leucine rich repeat | RPK | 1.14 |
| Solyc08g077630.3 | Protein kinase domain | RPK | 1.54 |
| Solyc01g098690.2 | Leucine rich repeat N-terminal domain | RPK | 1.23 |
| Solyc05g053010.1 | Protein kinase domain | RPK | 1.59 |
| Solyc07g064820.1 | Protein kinase domain | MAPK | 1.02 |
| Solyc11g020230.1 | Protein kinase domain | RPK | 1.05 |
| Solyc02g064980.1 | Protein kinase domain | MAPK | 3.15 |
| Solyc02g090970.1 | Protein kinase domain | MAPK | 3.05 |
| Solyc08g077560.3 | Protein tyrosine kinase | RPK | 1.42 |
| Solyc04g074270.3 | Leucine rich repeat | RPK | 1.08 |
| Solyc02g089900.1 | LysM domain | RPK | 1.07 |
| Solyc06g005170.3 | Protein kinase domain | MAPK | 1.65 |
| Solyc09g018280.1 | NAF domain | CDPK | 1.14 |
| Solyc02g092450.3 | E1-E2 ATPase | PM-CA-ATPase | 1.28 |
| Solyc02g064680.3 | E1-E2 ATPase | PM-CA-ATPase | 1.87 |
| Solyc01g099370.3 | C2 domain | CaLB | 1.61 |
| Solyc08g008370.3 | Development and cell death domain | CaLB | 1.66 |
| Solyc03g113980.3 | Calmodulin binding protein-like | CLM | 1.05 |
| Solyc06g053930.3 | EF-hand domain pair | CaM | 1.04 |
| Solyc03g097100.1 | EF-hand domain pair | CLM | 1.10 |
| Solyc11g071740.2 | EF-hand domain pair | CLM | 2.67 |
| Solyc03g118810.1 | EF-hand domain pair | CaM | 1.38 |
| Solyc03g119250.3 | Calmodulin binding protein-like | CLM | 1.99 |
| Solyc06g006020.2 | Leucine Rich Repeat | RPK | −1.31 |
| Solyc12g009510.1 | Leucine rich repeat | RPK | −1.13 |
| Solyc11g017280.2 | Leucine rich repeat | RPK | −1.07 |
| Solyc04g074020.2 | Leucine rich repeat N-terminal domain | RPK | −1.75 |
| Solyc04g009910.3 | Protein kinase domain | CDPK | −1.21 |
| Solyc12g009780.1 | Leucine Rich Repeat | RPK | −1.19 |
| Solyc04g074030.3 | Leucine rich repeat N-terminal domain | RPK | −1.91 |
| Solyc11g011180.2 | Leucine Rich repeat | RPK | −1.23 |
| Solyc05g012430.1 | Leucine rich repeat | RPK | −1.71 |
| Solyc09g090210.3 | Protein tyrosine kinase | RPK | −1.10 |
| Solyc07g006770.2 | TMEM154 protein family | RPK | −1.20 |
| Solyc10g076550.1 | Protein kinase domain | RPK | −1.85 |
| Solyc04g074050.3 | Protein kinase domain | RPK | −1.75 |
| Solyc01g067020.3 | Protein kinase domain | RPK | −1.16 |
| Solyc09g075920.1 | D-mannose binding lectin | RPK | −1.10 |
| Solyc08g016270.2 | Leucine rich repeat | RPK | −1.13 |
| Solyc04g074000.3 | Protein tyrosine kinase | RPK | −1.64 |
| Solyc08g077730.3 | MORN repeat | PIPK | −1.22 |
| Solyc04g009900.3 | Protein kinase domain | PPCK | −1.27 |
| Solyc12g088840.1 | EF-hand domain | CaM | −1.03 |
| Malic acid related DEGs | |||
| Solyc04g080990.2 | Voltage-dependent anion channel | 1.32 | |
| Solyc09g014610.3 | Voltage-dependent anion channel | 1.04 | |
| Solyc03g119640.3 | Aluminum activated malate transporter | 1.30 | |
| Gene ID | Annotation | LogFC |
|---|---|---|
| Solyc06g073750.3 | Glycosyl hydrolase | 1.99 |
| Solyc07g006850.2 | Glycosyl hydrolase | 1.04 |
| Solyc03g115380.2 | UDP-glucose/GDP-mannose dehydrogenase | 1.78 |
| Solyc02g094010.2 | Protein kinase domain | 1.58 |
| Solyc03g110890.1 | Polysaccharide biosynthesis related protein | 1.13 |
| Solyc09g075330.3 | Pectinesterase | 1.08 |
| Solyc09g011860.3 | GDP-fucose protein O-fucosyltransferase | 1.69 |
| Solyc12g010540.1 | GDP-mannose 4,6 dehydratase | 1.07 |
| Solyc04g005040.1 | Putative peptidoglycan binding domain | 1.37 |
| Solyc02g089640.3 | Glycosyltransferase-like | 1.03 |
| Solyc03g123620.4 | Plant invertase/pectin methylesterase inhibitor | 1.35 |
| Solyc03g097050.3 | RING/Ubox-like zinc-binding domain | 1.42 |
| Solyc06g074670.3 | NAD dependent epimerase/dehydratase | 1.31 |
| Solyc04g072280.3 | Multicopper oxidase | −2.95 |
| Solyc03g026360.1 | LysM domain | −1.23 |
| Solyc11g044910.2 | Glycosyl hydrolase domain | −1.66 |
| Compound | Mock | PEG30 | LogFC |
|---|---|---|---|
| Piruvate | 0.04 | 0.14 | 1.72 |
| beta-Aminoisobutiric acid | 3.99 | 11.99 | 1.59 |
| acid α-ketocaproate | 0.35 | 2.88 | 3.06 |
| Urea | 20.53 | 55.66 | 1.44 |
| Ethanolamine | 2.11 | 15.58 | 2.88 |
| Leucine | 2666.87 | 6457.33 | 1.28 |
| Succinate | 51.39 | 130.40 | 1.34 |
| Glycerate | 6.04 | 16.87 | 1.48 |
| Fumarate | 4.64 | 9.08 | 0.97 |
| Serine | 7.56 | 34.32 | 2.18 |
| Threonine | 8.33 | 50.54 | 2.60 |
| beta-Alanine | 1.19 | 17.05 | 3.84 |
| Glutamine | 1.51 | 6.59 | 2.13 |
| Malate | 2.20 | 5.55 | 1.33 |
| Cytosine | 0.05 | 0.17 | 1.72 |
| L-Aspartate | 48.05 | 160.37 | 1.74 |
| Ornithin-Citrullin-Arginine | 1.09 | 4.55 | 2.06 |
| Xylose MeOX1 | 0.12 | 1.55 | 3.65 |
| Ribose | 0.22 | 0.64 | 1.54 |
| cis-Aconitate | 0.18 | 0.56 | 1.65 |
| 1-Methyl-L-histidine | 0.11 | 0.49 | 2.19 |
| Glycerate-3-P | 0.40 | 0.97 | 1.28 |
| Citrullin-Ornithin-Arginine | 6.14 | 78.11 | 3.67 |
| Citrate | 2.13 | 6.83 | 1.68 |
| Isocitrate | 0.52 | 1.45 | 1.47 |
| Arginine-NH3 | 8.62 | 16.92 | 0.97 |
| Pinitol | 0.10 | 0.28 | 1.50 |
| Fructose MEOX1 | 0.09 | 0.30 | 1.78 |
| Fructose MEOX2 | 0.04 | 0.12 | 1.52 |
| Galactose MeOX1 | 0.11 | 0.38 | 1.84 |
| Glucose MEOX1 | 0.46 | 1.46 | 1.67 |
| Glucose MEOX2 | 0.04 | 0.17 | 1.99 |
| Glucuronic acid MEOX1 | 0.03 | 0.13 | 2.04 |
| Gluconate | 0.23 | 0.59 | 1.35 |
| myo-Inositol | 7.21 | 24.71 | 1.78 |
| Spermidine | 0.45 | 3.41 | 2.93 |
| L-Cystathionine | 0.71 | 3.80 | 2.42 |
| Tryptophan | 112.05 | 222.43 | 0.99 |
| Fructose-6-P | 0.04 | 0.09 | 1.18 |
| Gluconate-6-P | 0.03 | 0.13 | 2.17 |
| Sucrose | 0.59 | 2.00 | 1.75 |
| Trehalose | 0.33 | 0.66 | 0.98 |
| Lysine | 4737.93 | 12276.77 | 1.37 |
| Histidine | 193.44 | 393.00 | 1.02 |
| Maleic acid | 9.10 | 2.61 | −1.80 |
| Uracil | 51.66 | 13.95 | −1.89 |
| Thymine | 6.94 | 0.16 | −5.46 |
| 4-Aminobutyrate (GABA) | 20.49 | 3.28 | −2.64 |
| Gene ID | Annotation | LogFC |
|---|---|---|
| Solyc10g054440.2 | Pyridoxal-dependent decarboxylase | 1.46 |
| Solyc02g089610.2 | Adenosylmethionine decarboxylase | 1.39 |
| Solyc02g089615.1 | S-adenosyl-l-methionine decarboxylase leader peptide | 1.31 |
| Solyc08g007820.1 | Dehydration-responsive element-binding protein 1E-like | 3.02 |
| Solyc03g080090.3 | No apical meristem (NAM) protein | 1.08 |
| Solyc07g008300.2 | Ring hydroxylating alpha subunit | 2.28 |
| Solyc08g068600.3 | Pyridoxal-dependent decarboxylase conserved domain | 1.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcillo, R.J.L.; Vílchez, J.I.; Zhang, S.; Kaushal, R.; He, D.; Zi, H.; Liu, R.; Niehaus, K.; Handa, A.K.; Zhang, H. Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria. Metabolites 2021, 11, 369. https://doi.org/10.3390/metabo11060369
Morcillo RJL, Vílchez JI, Zhang S, Kaushal R, He D, Zi H, Liu R, Niehaus K, Handa AK, Zhang H. Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria. Metabolites. 2021; 11(6):369. https://doi.org/10.3390/metabo11060369
Chicago/Turabian StyleMorcillo, Rafael J. L., Juan I. Vílchez, Song Zhang, Richa Kaushal, Danxia He, Hailing Zi, Renyi Liu, Karsten Niehaus, Avtar K. Handa, and Huiming Zhang. 2021. "Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria" Metabolites 11, no. 6: 369. https://doi.org/10.3390/metabo11060369
APA StyleMorcillo, R. J. L., Vílchez, J. I., Zhang, S., Kaushal, R., He, D., Zi, H., Liu, R., Niehaus, K., Handa, A. K., & Zhang, H. (2021). Plant Transcriptome Reprograming and Bacterial Extracellular Metabolites Underlying Tomato Drought Resistance Triggered by a Beneficial Soil Bacteria. Metabolites, 11(6), 369. https://doi.org/10.3390/metabo11060369