
Deuterium Magnetic Resonance Imaging and the Discrimination of Fetoplacental Metabolism in Normal and L-NAME-Induced Preeclamptic Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
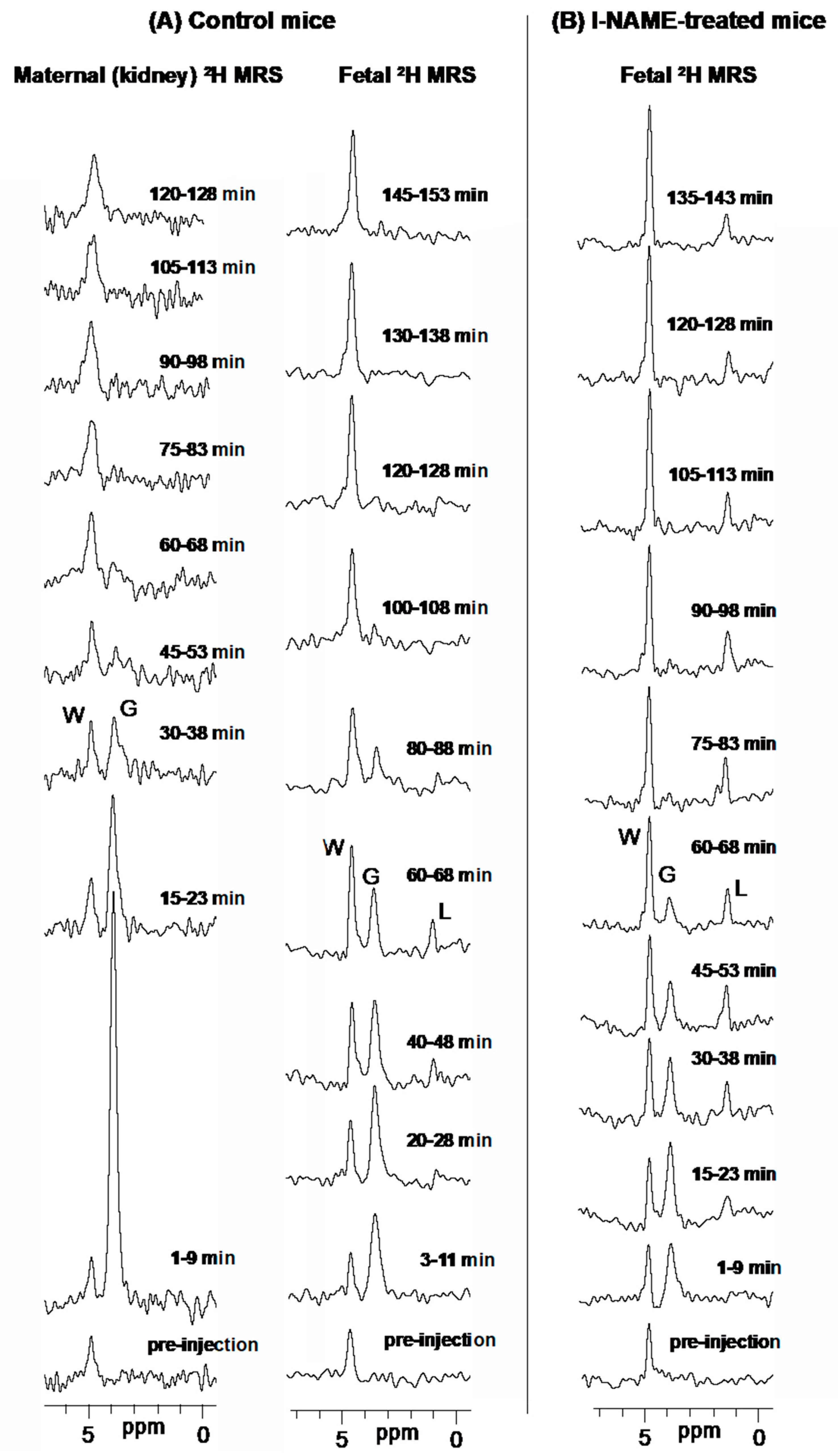
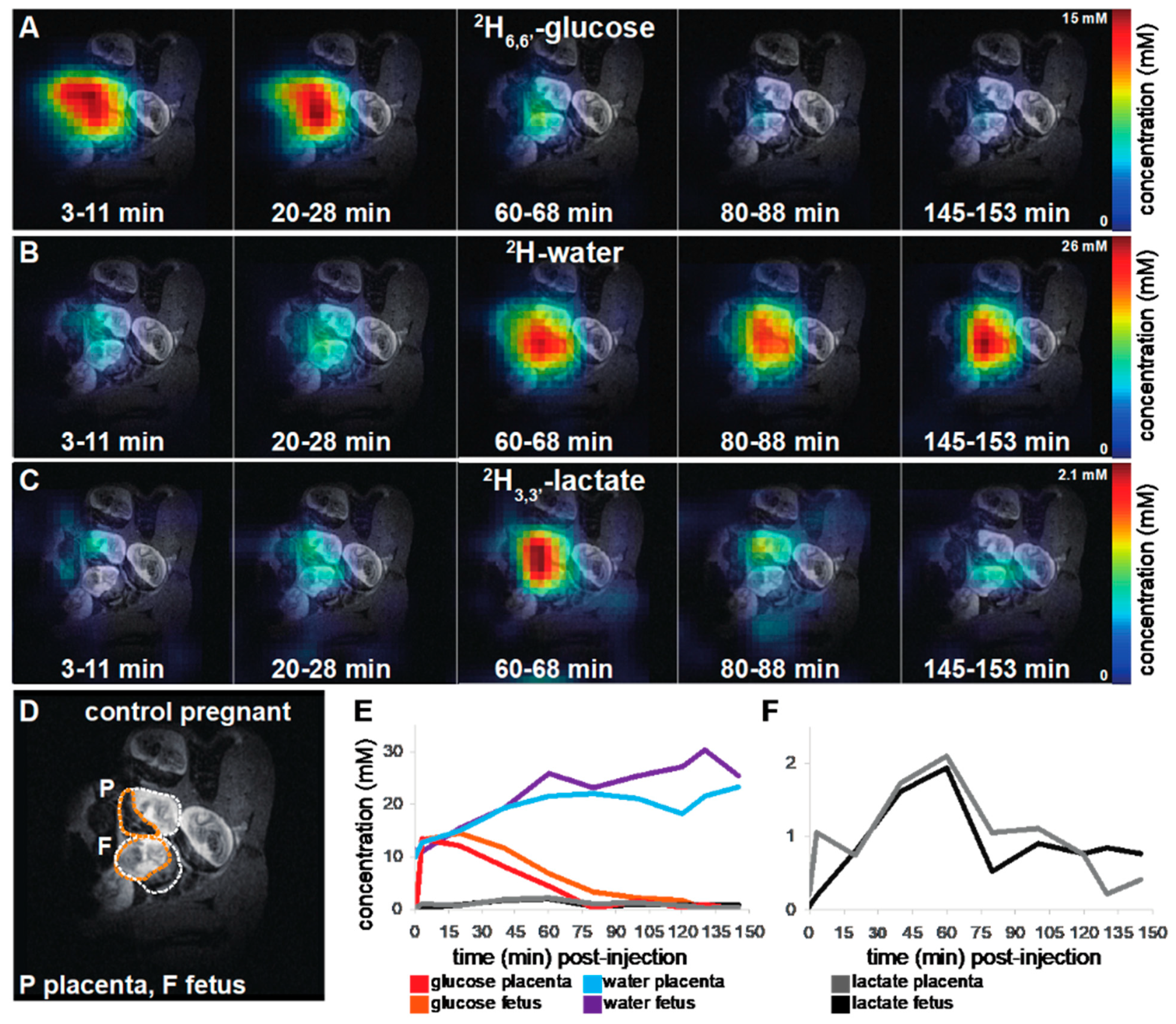
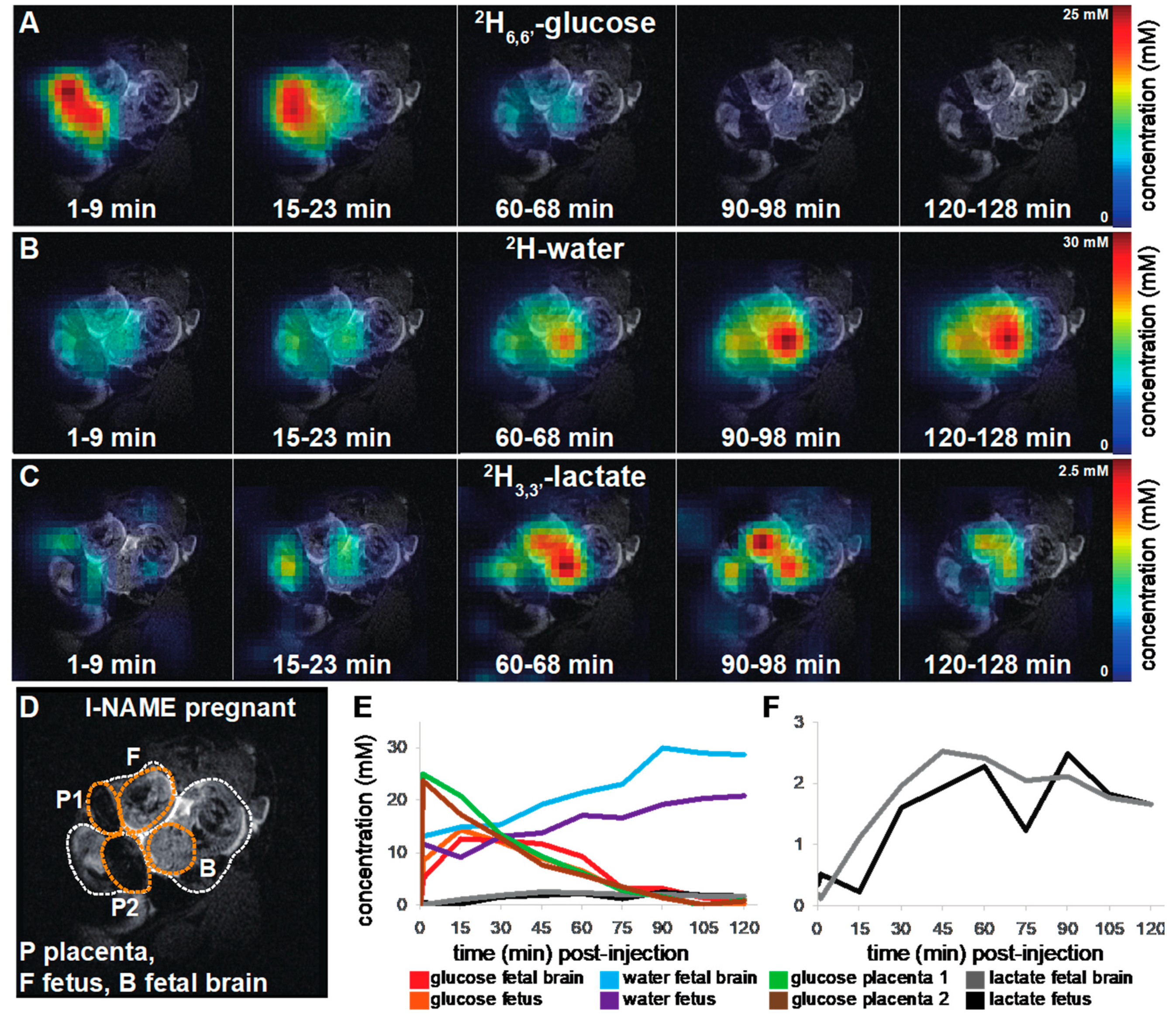
2.1. 2H magnetic Resonance on Pregnant Animals: Overall Features
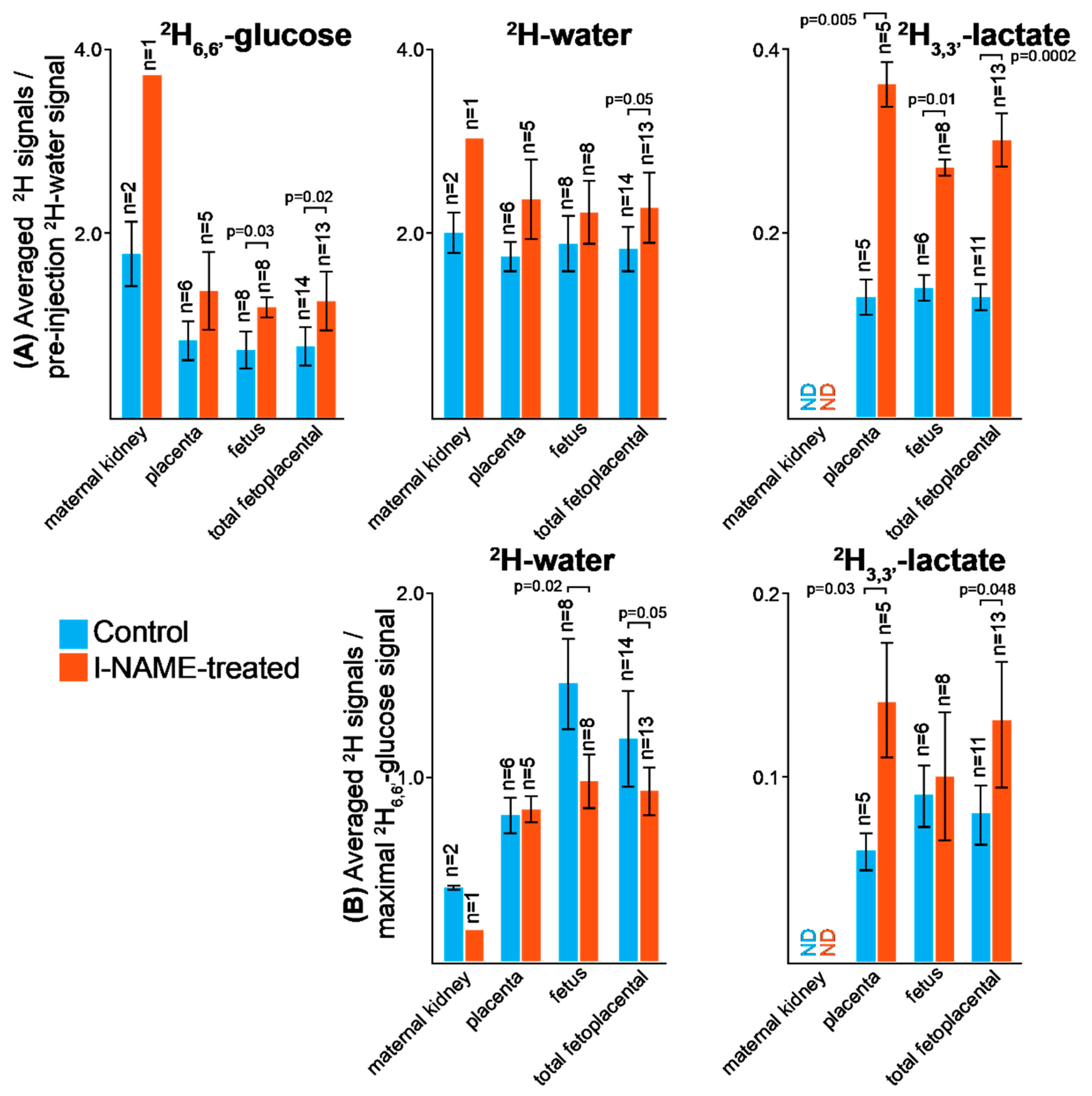
2.2. DMI Reveals Differences between Normal and Preeclamptic Pregnancies
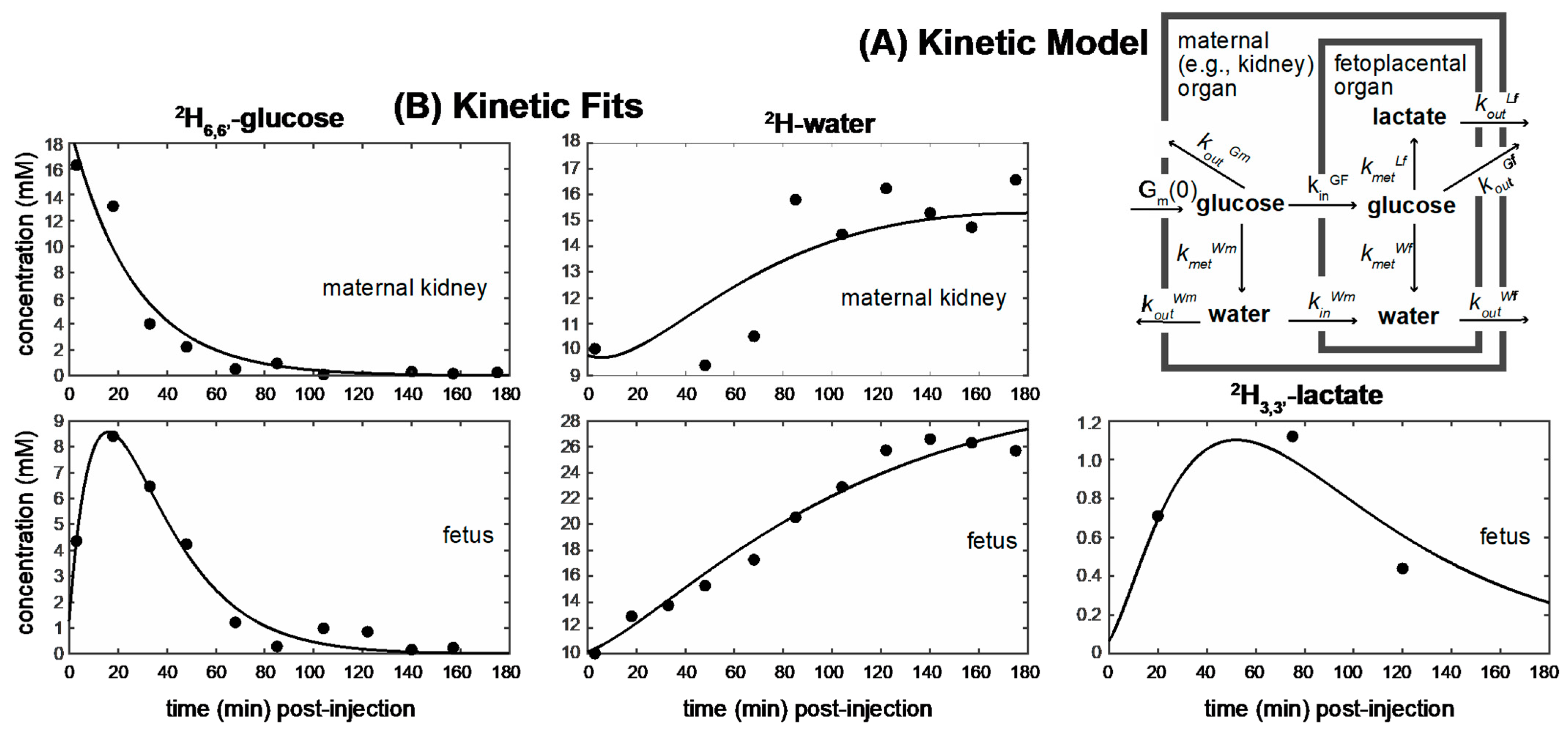
2.3. Kinetic Analyses of the DMI Data
2.4. Ancillary 2H2,2′-Glycine Experiments
3. Discussion
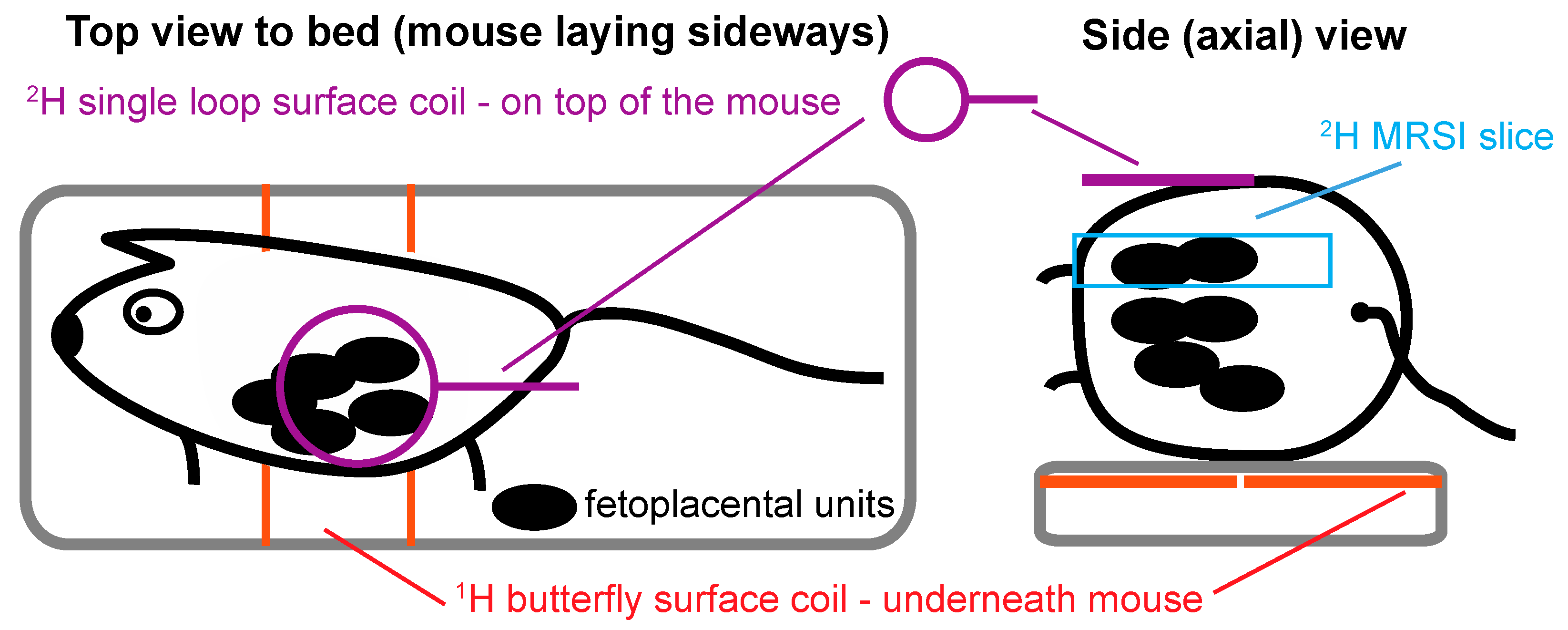
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simeoni, U.; Armengaud, J.-B.; Siddeek, B.; Tolsa, J.-F. Perinatal Origins of Adult Disease. Neonatology 2018, 113, 393–399. [Google Scholar] [CrossRef]
- Cuffe, J.S.; Xu, Z.C.; Perkins, A.V. Biomarkers of Oxidative Stress in Pregnancy Complications. Biomark. Med. 2017, 11, 295–306. [Google Scholar] [CrossRef]
- Abramowicz, J.; Sheiner, E. Ultrasound of the Placenta: A Systematic Approach. Part I: Imaging. Placenta 2008, 29, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, K.; Mohan, A.R. Imaging in pregnancy. Obstet. Gynaecol. 2019, 21, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Reddy, U.M.; Filly, R.A.; Copel, J.A. Prenatal imaging: Ultrasonography and magnetic resonance imaging. Obstet. Gynecol. 2008, 112, 145–157. [Google Scholar] [CrossRef]
- Wang, P.I.; Chong, S.T.; Kielar, A.Z.; Kelly, A.M.; Knoepp, U.D.; Mazza, M.B.; Goodsitt, M.M. Imaging of Pregnant and Lactating Patients: Part 1, Evidence-Based Review and Recommendations. Am. J. Roentgenol. 2012, 198, 778–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avni, R.; Golani, O.; Akselrod-Ballin, A.; Cohen, Y.; Biton, I.; Garbow, J.R.; Neeman, M. MR Imaging-derived Oxygen-Hemoglobin Dissociation Curves and Fetal-Placental Oxygen-Hemoglobin Affinities. Radiology 2016, 280, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avni, R.; Neeman, M.; Garbow, J. Functional MRI of the placenta—From rodents to humans. Placenta 2015, 36, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Bao, Q.; Liberman, G.; Solomon, E.; Frydman, L. High-resolution diffusion MRI studies of development in pregnant mice visualized by novel spatiotemporal encoding schemes. NMR Biomed. 2020, 33, e4208. [Google Scholar] [CrossRef]
- Bonel, H.M.; Stolz, B.; Diedrichsen, L.; Frei, K.; Saar, B.; Tutschek, B.; Raio, L.; Surbek, D.; Srivastav, S.; Nelle, M.; et al. Diffusion-weighted MR Imaging of the Placenta in Fetuses with Placental Insufficiency. Radiology 2010, 257, 810–819. [Google Scholar] [CrossRef] [Green Version]
- Gowland, P.A.; Francis, S.T.; Duncan, K.R.; Freeman, A.J.; Issa, B.; Moore, R.J.; Bowtell, R.; Baker, P.; Johnson, I.R.; Worthington, B.S. In vivo perfusion measurements in the human placenta using echo planar imaging at 0.5 T. Magn. Reson. Med. 1998, 40, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Huen, I.; Morris, D.M.; Wright, C.; Parker, G.; Sibley, C.P.; Johnstone, E.D.; Naish, J.H. R 1 and R 2 * changes in the human placenta in response to maternal oxygen challenge. Magn. Reson. Med. 2013, 70, 1427–1433. [Google Scholar] [CrossRef]
- Morita, S.; Ueno, E.; Fujimura, M.; Muraoka, M.; Takagi, K.; Fujibayashi, M. Feasibility of diffusion-weighted MRI for defining placental invasion. J. Magn. Reson. Imaging 2009, 30, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.F.; Confort-Gouny, S.; Le Fur, Y.; Viout, P.; Bennathan, M.; Chapon, F.; Fogliarini, C.; Cozzone, P.; Girard, N. Diffusion-weighted imaging in normal fetal brain maturation. Eur. Radiol. 2007, 17, 2422–2429. [Google Scholar] [CrossRef] [Green Version]
- Sohlberg, S.; Mulic-Lutvica, A.; Lindgren, P.; Ortiz-Nieto, F.; Wikström, A.-K.; Wikström, J. Placental perfusion in normal pregnancy and early and late preeclampsia: A magnetic resonance imaging study. Placenta 2014, 35, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.; Avni, R.; Hadas, R.; Raz, T.; Garbow, J.R.; Bendel, P.; Frydman, L.; Neeman, M. Major mouse placental compartments revealed by diffusion-weighted MRI, contrast-enhanced MRI, and fluorescence imaging. Proc. Natl. Acad. Sci. USA 2014, 111, 10353–10358. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, A.; Peters, D.; Simonsen, C.; Pedersen, M.; Stausbøl-Grøn, B.; Christiansen, O.B.; Lingman, G.; Uldbjerg, N. Changes in human fetal oxygenation during maternal hyperoxia as estimated by BOLD MRI. Prenat. Diagn. 2012, 33, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Taillieu, F.; Salomon, L.J.; Siauve, N.; Clément, O.; Faye, N.; Balvay, D.; Vayssettes, C.; Frija, G.; Ville, Y.; Cuenod, C.A. Placental perfusion and permeability: Simultaneous assessment with dual-echo contrast-enhanced MR imaging in mice. Radiology 2006, 241, 737–745. [Google Scholar] [CrossRef]
- Wu, D.; Lei, J.; Rosenzweig, J.M.; Burd, I.; Zhang, J. In utero localized diffusion MRI of the embryonic mouse brain microstructure and injury. J. Magn. Reson. Imaging 2014, 42, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Denison, F.C.; Semple, S.I.; Stock, S.J.; Walker, J.; Marshall, I.; Norman, J.E. Novel use of proton magnetic resonance spectroscopy (1HMRS) to non-invasively assess placental metabolism. PLoS ONE 2012, 7, e42926. [Google Scholar] [CrossRef] [Green Version]
- Sohlberg, S.; Wikström, A.K.; Olovsson, M.; Lindgren, P.; Axelsson, O.; Mulic-Lutvica, A.; Weis, J.; Wikström, J. In vivo 31P-MR spectroscopy in normal pregnancy, early and late preeclampsia: A study of placental metabolism. Placenta 2014, 35, 318–323. [Google Scholar] [CrossRef]
- Levine, D.; Barnes, P.D.; Edelman, R.R. Obstetric MR Imaging. Radiology 1999, 211, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Zheleznyak, A.; Garbow, J.R.; Neeman, M.; Lapi, S.E. Preclinical Positron Emission Tomographic Imaging of Acute Hyperoxia Therapy of Chronic Hypoxia during Pregnancy. Mol. Imaging 2015, 14, 366–372. [Google Scholar] [CrossRef]
- Takalkar, A.M.; Khandelwal, A.; Lokitz, S.; Lilien, D.L.; Stabin, M.G. 18F-FDG PET in Pregnancy and Fetal Radiation Dose Estimates. J. Nucl. Med. 2011, 52, 1035–1040. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, J.; Semelka, R.C.; Nunes, R.H.; AlObaidy, M.; Castillo, M. Gadolinium-Based Contrast Agent Accumulation and Toxicity: An Update. Am. J. Neuroradiol. 2015, 37, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Yadav, B.K.; Neelavalli, J.; Krishnamurthy, U.; Szalai, G.; Shen, Y.; Nayak, N.R.; Chaiworapongsa, T.; Hernandez-Andrade, E.; Than, N.G.; Haacke, E.M.; et al. A longitudinal study of placental perfusion using dynamic contrast enhanced magnetic resonance imaging in murine pregnancy. Placenta 2016, 43, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Ardenkjaer-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of > 10,000 times in liquid-state NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158–10163. [Google Scholar] [CrossRef] [Green Version]
- Brindle, K.M. Imaging Metabolism with Hyperpolarized13C-Labeled Cell Substrates. J. Am. Chem. Soc. 2015, 137, 6418–6427. [Google Scholar] [CrossRef] [PubMed]
- Golman, K.; Ardenkjaer-Larsen, J.H.; Petersson, J.S.; Mansson, S.; Leunbach, I. Molecular imaging with endogenous substances. Proc. Natl. Acad. Sci. USA 2003, 100, 10435–10439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golman, K.; Thaning, M. Real-time metabolic imaging. Proc. Natl. Acad. Sci. USA 2006, 103, 11270–11275. [Google Scholar] [CrossRef] [Green Version]
- Markovic, S.; Fages, A.; Roussel, T.; Hadas, R.; Brandis, A.; Neeman, M.; Frydman, L. Placental physiology monitored by hyperpolarized dynamic 13C magnetic resonance. Proc. Natl. Acad. Sci. USA 2018, 115, e2429–e2436. [Google Scholar] [CrossRef] [Green Version]
- Friesen-Waldner, L.J.; Sinclair, K.J.; Wade, T.P.; Michael, B.; Chen, A.P.; De Vrijer, B.; Regnault, T.; McKenzie, C.A. Hyperpolarized [1-13C]pyruvate MRI for noninvasive examination of placental metabolism and nutrient transport: A feasibility study in pregnant guinea pigs. J. Magn. Reson. Imaging 2015, 43, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, E.; Lauridsen, H.; Nielsen, P.M.; Qi, H.; Nørlinger, T.; Andersen, M.D.; Uldbjerg, N.; Laustsen, C.; Sandager, P.; Pedersen, M. The chinchilla as a novel animal model of pregnancy. R. Soc. Open Sci. 2017, 4, 161098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, K.E.; Mariager, C.Ø.; Duvald, C.S.; Hansen, E.S.S.; Bertelsen, L.B.; Pedersen, M.; Pedersen, L.H.; Uldbjerg, N.; Laustsen, C. Ex Vivo Human Placenta Perfusion, Metabolic and Functional Imaging for Obstetric Research—A Feasibility Study. Tomography 2019, 5, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.W.Y.; McMahon, M.T.; Kato, Y.; Liu, G.; Bulte, J.W.M.; Bhujwalla, Z.M.; Artemov, D.; van Zijl, P.C.M. NaturalD-glucose as a biodegradable MRI contrast agent for detecting cancer. Magn. Reson. Med. 2012, 68, 1764–1773. [Google Scholar] [CrossRef] [Green Version]
- Walker-Samuel, S.; Ramasawmy, R.; Torrealdea, F.; Rega, M.; Rajkumar, V.; Johnson, S.P.; Richardson, S.; Gonçalves, M.; Parkes, H.G.; Arstad, E.; et al. In vivo imaging of glucose uptake and metabolism in tumors. Nat. Med. 2013, 19, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Zaiss, M.; Bachert, P. Chemical exchange saturation transfer (CEST) and MR Z-spectroscopy in vivo: A review of theoretical approaches and methods. Phys. Med. Biol. 2013, 58, R221–R269. [Google Scholar] [CrossRef]
- Wu, D.; Xu, J.; Lei, J.; Mclane, M.; van Zijl, P.C.; Burd, I. Dynamic glucose enhanced MRI of the placenta in a mouse model of intrauterine inflammation. Placenta 2018, 69, 86–91. [Google Scholar] [CrossRef]
- Lu, M.; Zhu, X.H.; Zhang, Y.; Mateescu, G.; Chen, W. Quantitative assessment of brain glucose metabolic rates using in vivo deuterium magnetic resonance spectroscopy. J. Cereb. Blood Flow Metab. 2017, 37, 3518–3530. [Google Scholar] [CrossRef] [PubMed]
- De Feyter, H.M.; Behar, K.L.; Corbin, Z.A.; Fulbright, R.K.; Brown, P.B.; McIntyre, S.; Nixon, T.W.; Rothman, D.L.; De Graaf, R.A. Deuterium metabolic imaging (DMI) for MRI-based 3D mapping of metabolism in vivo. Sci. Adv. 2018, 4, eaat7314. [Google Scholar] [CrossRef] [Green Version]
- Kreis, F.; Wright, A.J.; Hesse, F.; Fala, M.; Hu, D.-E.; Brindle, K.M. Measuring Tumor Glycolytic Flux in Vivo by Using Fast Deuterium MRI. Radiology 2020, 294, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhu, X.H.; Zhu, W.; Lee, B.Y.; Wiesner, H.M.; Zhang, Y.; Wang, T.; Chen, W. Dynamic Deuterium MRS Imaging for Studying Rat Heart Energy Metabolism In Vivo—Initial Experience. Proc. Intl. Soc. Mag. Reson. Med. 2019, 27. [Google Scholar]
- Riis-Vestergaard, M.J.; Laustsen, C.; Mariager, C.Ø.; Schulte, R.F.; Pedersen, S.B.; Richelsen, B. Glucose metabolism in brown adipose tissue determined by deuterium metabolic imaging in rats. Int. J. Obes. 2020, 44, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- De Feyter, H.M.; Thomas, M.A.; Behar, K.L.; de Graaf, R.A. NMR visibility of deuterium-labeled liver glycogen in vivo. Magn. Reson. Med. 2021, 86, 62–68. [Google Scholar] [CrossRef]
- Hesse, F.; Somai, V.; Kreis, F.; Bulat, F.; Wright, A.J.; Brindle, K.M. Monitoring tumor cell death in murine tumor models using deuterium magnetic resonance spectroscopy and spectroscopic imaging. Proc. Natl. Acad. Sci. USA 2021, 118, e2014631118. [Google Scholar] [CrossRef] [PubMed]
- Straathof, M.; Meerwaldt, A.E.; De Feyter, H.M.; de Graaf, R.A.; Dijkhuizen, R.M. Deuterium Metabolic Imaging of the Healthy and Diseased Brain. Neuroscience 2021. [Google Scholar] [CrossRef]
- Von Morze, C.; Engelbach, J.A.; Blazey, T.; Quirk, J.D.; Reed, G.D.; Ippolito, J.E.; Garbow, J.R. Comparison of hyperpolarized (13) C and non-hyperpolarized deuterium MRI approaches for imaging cerebral glucose metabolism at 4.7 T. Magn. Reson. Med. 2021, 85, 1795–1804. [Google Scholar] [CrossRef] [PubMed]
- Markovic, S.; Roussel, T.; Preise, D.; Sasson, K.; Agemy, L.; Scherz, A.; Frydman, L. Deuterium MRSI characterizations of glucose metabolism in orthotopic pancreatic cancer mouse models. NMR Biomed. 2021. [Google Scholar] [CrossRef]
- Mahar, R.; Donabedian, P.L.; Merritt, M.E. HDO production from [2H7]glucose Quantitatively Identifies Warburg Metabolism. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- De Graaf, R.A.; Hendriks, A.D.; Klomp, D.W.; Kumaragamage, C.; Welting, D.; Arteaga de Castro, C.S.; Brown, P.B.; McIntyre, S.; Nixon, T.W.; Prompers, J.J.; et al. On the magnetic field dependence of deuterium metabolic imaging. NMR Biomed. 2020, 33, e4235. [Google Scholar] [CrossRef]
- Vangrieken, P.; Al-Nasiry, S.; Bast, A.; Leermakers, P.A.; Tulen, C.B.M.; Schiffers, P.M.H.; van Schooten, F.J.; Remels, A.H.V. Placental Mitochondrial Abnormalities in Preeclampsia. Reprod. Sci. 2021, 1–14. [Google Scholar] [CrossRef]
- De Souza, C.O.; Peraçoli, M.T.S.; Weel, I.C.; Bannwart, C.F.; Romão, M.; Nakaira-Takahagi, É.; de Medeiros, L.T.L.; da Silva, M.G.; Peraçoli, J.C. Hepatoprotective and anti-inflammatory effects of silibinin on experimental preeclampsia induced by L-NAME in rats. Life Sci. 2012, 91, 159–165. [Google Scholar] [CrossRef]
- Fantel, A.G.; Person, R.E. Involvement of mitochondria and other free radical sources in normal and abnormal fetal development. Ann. N. Y. Acad. Sci. 2002, 959, 424–433. [Google Scholar] [CrossRef]
- Bao, Q.; Hadas, R.; Markovic, S.; Neeman, M.; Frydman, L. Diffusion and perfusion MRI of normal, preeclamptic and growth-restricted mice models reveal clear fetoplacental differences. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Prior, R.L. Glucose and lactate metabolism in vivo in ovine fetus. Am. J. Physiol. Metab. 1980, 239, e208–e214. [Google Scholar] [CrossRef] [PubMed]
- Sparks, J.W.; Hay, W.W.; Bonds, D.; Meschia, G.; Battaglia, F.C. Simultaneous measurements of lactate turnover rate and umbilical lactate uptake in the fetal lamb. J. Clin. Investig. 1982, 70, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yoseph, O.; Kingsley, P.B.; Ross, B.D. Metabolic loss of deuterium from isotopically labeled glucose. Magn. Reson. Med. 1994, 32, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Marconi, A.M.; Cetin, I.; Ferrazzi, E.; Ferrari, M.M.; Pardi, G.; Battaglia, F.C. Lactate Metabolism in Normal and Growth-Retarded Human Fetuses. Pediatr. Res. 1990, 28, 652–656. [Google Scholar] [CrossRef] [Green Version]
- Père, M.-C. Materno-foetal exchanges and utilisation of nutrients by the foetus: Comparison between species. Reprod. Nutr. Dev. 2003, 43, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, H.; Zhu, S.; Tsoi, S. Hypoxia and Lactate Production in Trophoblast Cells. Placenta 2007, 28, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Jaiswar, S.P.; Gupta, A.; Sachan, R.; Natu, S.N.; Shaili, M. Lactic Dehydrogenase: A Biochemical Marker for Preeclampsia–Eclampsia. J. Obstet. Gynecol. India 2011, 61, 645–648. [Google Scholar] [CrossRef] [Green Version]
- Economides, D.; Proudler, A.; Nicolaides, K. Plasma insulin in appropriate- and small-for-gestational-age fetuses. Am. J. Obstet. Gynecol. 1989, 160, 1091–1094. [Google Scholar] [CrossRef]
- Marconi, A.M.; Paolini, C.; Buscaglia, M.; Zerbe, G.; Battaglia, F.C.; Pardi, G. The Impact of Gestational Age and Fetal Growth on the Maternal-Fetal Glucose Concentration Difference. Obstet. Gynecol. 1996, 87, 937–942. [Google Scholar] [CrossRef]
- Soothill, P.W.; Nicolaides, K.H.; Campbell, S. Prenatal asphyxia, hyperlacticaemia, hypoglycaemia, and erythroblastosis in growth retarded fetuses. BMJ 1987, 294, 1051–1053. [Google Scholar] [CrossRef] [Green Version]
- Challis, E.D.; Pfarrer, C.D.; Ritchie, J.W.K.; Koren, G.; Adamson, S.L. Glucose Metabolism Is Elevated and Vascular Resistance and Maternofetal Transfer Is Normal in Perfused Placental Cotyledons from Severely Growth-Restricted Fetuses. Pediatr. Res. 2000, 47, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K. Hypoxia in the pathogenesis of preeclampsia. Hypertens. Res. Pregnancy 2017, 5, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Verhaeghe, J.; Pintiaux, A.; Van Herck, E.; Hennen, G.; Foidart, J.M.; Igout, A. Placental GH, IGF-I, IGF-binding protein-1, and leptin during a glucose challenge test in pregnant women: Relation with maternal body weight, glucose tolerance, and birth weight. J. Clin. Endocrinol. Metab. 2002, 87, 2875–2882. [Google Scholar] [CrossRef]
- Soonthornpun, S.; Aksonteing, J.; Thamprasit, A. A comparison between a 75-g and 100-g oral glucose tolerance test in pregnant women. Int. J. Gynecol. Obstet. 2003, 81, 169–173. [Google Scholar] [CrossRef]
- Harris, T.; Degani, H.; Frydman, L. Hyperpolarized 13 C NMR studies of glucose metabolism in living breast cancer cell cultures. NMR Biomed. 2013, 26, 1831–1843. [Google Scholar] [CrossRef]
- Rodrigues, T.B.; Serrao, E.M.; Kennedy, B.W.; Hu, D.-E.; Kettunen, M.I.; Brindle, K.M. Magnetic resonance imaging of tumor glycolysis using hyperpolarized 13C-labeled glucose. Nat. Med. 2014, 20, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Peters, D.C.; Markovic, S.; Bao, Q.; Preise, D.; Sasson, K.; Agemy, L.; Scherz, A.; Frydman, L. Improving deuterium metabolic imaging (DMI) signal to noise ratio by spectroscopic multi-echo bSSFP: A pancreatic cancer investigation. Magn. Reson. Med. 2021. [Google Scholar] [CrossRef]
- Hu, C.; Peters, D.C. SUPER: A blockwise curve-fitting method for accelerating MR parametric mapping with fast reconstruction. Magn. Reson. Med. 2019, 81, 3515–3529. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markovic, S.; Roussel, T.; Neeman, M.; Frydman, L. Deuterium Magnetic Resonance Imaging and the Discrimination of Fetoplacental Metabolism in Normal and L-NAME-Induced Preeclamptic Mice. Metabolites 2021, 11, 376. https://doi.org/10.3390/metabo11060376
Markovic S, Roussel T, Neeman M, Frydman L. Deuterium Magnetic Resonance Imaging and the Discrimination of Fetoplacental Metabolism in Normal and L-NAME-Induced Preeclamptic Mice. Metabolites. 2021; 11(6):376. https://doi.org/10.3390/metabo11060376
Chicago/Turabian StyleMarkovic, Stefan, Tangi Roussel, Michal Neeman, and Lucio Frydman. 2021. "Deuterium Magnetic Resonance Imaging and the Discrimination of Fetoplacental Metabolism in Normal and L-NAME-Induced Preeclamptic Mice" Metabolites 11, no. 6: 376. https://doi.org/10.3390/metabo11060376
APA StyleMarkovic, S., Roussel, T., Neeman, M., & Frydman, L. (2021). Deuterium Magnetic Resonance Imaging and the Discrimination of Fetoplacental Metabolism in Normal and L-NAME-Induced Preeclamptic Mice. Metabolites, 11(6), 376. https://doi.org/10.3390/metabo11060376