
Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows

 ,
, 
 ,
,
Abstract
:1. Introduction
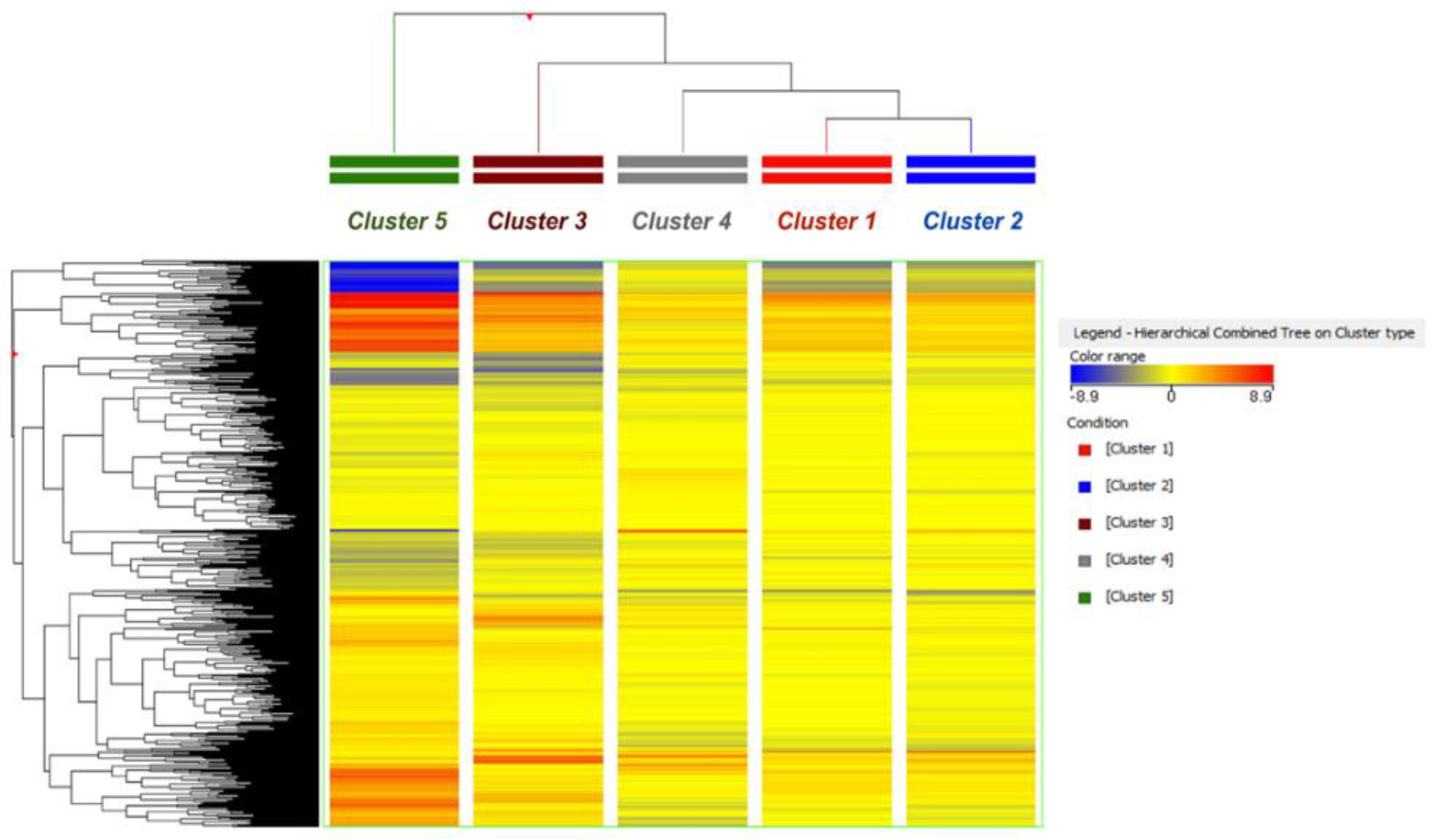
2. Results and Discussion
2.1. Metabolomic Discrimination of Milk Samples According to Corn Silages’ Contamination
2.2. Pathway Analysis and Significant Changes of Milk Metabolites
3. Materials and Methods
3.1. Collection of Milk Samples
3.2. Extraction of Milk Metabolites
3.3. Untargeted Metabolomics Analysis
3.4. Multivariate Statistics and Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cattani, M.; Guzzo, N.; Mantovani, R.; Bailoni, L. Effects of total replacement of corn silage with sorghum silage on milk yield, composition, and quality. J. Anim. Sci. Biotechnol. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandicke, J.; De Visschere, K.; Ameye, M.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Multi-mycotoxin contamination of maize silages in Flanders, Belgium: Monitoring mycotoxin levels from seed to feed. Toxins 2021, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. Part A 2008, 25, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Ghilardelli, F.; Doupovec, B.; Faas, J.; Schatzmayr, D.; Masoero, F. Kinetics of gas production in the presence of Fusarium mycotoxins in rumen fluid of lactating dairy cows. JDS Commun. 2021. [Google Scholar] [CrossRef]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; De Gelder, L.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage review: Mycotoxins in silage: Occurrence, effects, prevention, and mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef] [PubMed]
- Kemboi, D.C.; Antonissen, G.; Ochieng, P.E.; Croubels, S.; Okoth, S.; Kangethe, E.K.; Faas, J.; Lindahl, J.F.; Gathumbi, J.K. A review of the impact of mycotoxins on dairy cattle health: Challenges for food safety and dairy production in Sub-Saharan Africa. Toxins 2020, 12, 222. [Google Scholar] [CrossRef] [Green Version]
- Loh, Z.H.; Ouwerkerk, D.; Klieve, A.V.; Hungerford, N.L.; Fletcher, M.T. Toxin degradation by rumen microorganisms: A review. Toxins 2020, 12, 664. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Ghilardelli, F.; Atzori, A.S.; Zara, S.; Novak, B.; Faas, J.; Fancello, F. Co-occurrence of regulated and emerging mycotoxins in corn silage: Relationships with fermentation quality and bacterial communities. Toxins 2021, 13, 232. [Google Scholar] [CrossRef]
- Schmidt, P.; Novinski, C.O.; Junges, D.; Almeida, R.; de Souza, C.M. Concentration of mycotoxins and chemical composition of corn silage: A farm survey using infrared thermography. J. Dairy Sci. 2015, 98, 6609–6619. [Google Scholar] [CrossRef]
- Signorini, M.L.; Gaggiotti, M.; Molineri, A.; Chiericatti, C.A.; Zapata de Basílico, M.L.; Basílico, J.C.; Pisani, M. Exposure assessment of mycotoxins in cow’s milk in Argentina. Food Chem. Toxicol. 2012, 50, 250–257. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Bruschi, S.; Fortunati, P.; Masoero, F. Use of principal factor analysis to generate a corn silage fermentative quality index to rank well- or poorly preserved forages. J. Sci. Food Agric. 2016, 96, 1686–1696. [Google Scholar] [CrossRef]
- Tharangani, R.M.H.; Yakun, C.; Zhao, L.S.; Ma, L.; Liu, H.L.; Su, S.L.; Shan, L.; Yang, Z.N.; Kononoff, P.J.; Weiss, W.P.; et al. Corn silage quality index: An index combining milk yield, silage nutritional and fermentation parameters. Anim. Feed Sci. Technol. 2021, 273, 114817. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Zheng, N.; Guo, L.; Song, X.; Zhao, S.; Wang, J. Biological system responses of dairy cows to aflatoxin B1 exposure revealed with metabolomic changes in multiple biofluids. Toxins 2019, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.-Z.; Wang, D.-M.; Wang, B.; Wang, J.-K.; Liu, H.-Y.; Guan, L.L.; Liu, J.-X. Metabolomics of four biofluids from dairy cows: Potential biomarkers for milk production and quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef]
- Eshelli, M.; Mallique Qader, M.; Jambi, E.J.; Hursthouse, A.S.; Rateb, M.E. Current status and future opportunities of omics tools in mycotoxin research. Toxins 2018, 10, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.-K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR metabolomic analysis of dairy cows reveals milk glycerophosphocholine to phosphocholine ratio as prognostic biomarker for risk of ketosis. J. Proteome Res. 2012, 11, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated metabolomics study of the milk of heat-stressed lactating dairy cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Rocchetti, G.; O’Callaghan, T.F. Application of metabolomics to assess milk quality and traceability. Curr. Opin. Food Sci. 2021, 40, 168–178. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gallo, A.; Nocetti, M.; Lucini, L.; Masoero, F. Milk metabolomics based on ultra-high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry to discriminate different cows feeding regimens. Food Res. Int. 2020, 134, 109279. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 13. [Google Scholar] [CrossRef]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef]
- Zhu, W.; Wei, Z.; Xu, N.; Yang, F.; Yoon, I.; Chung, Y.; Liu, J.; Wang, J. Effects of Saccharomyces cerevisiae fermentation products on performance and rumen fermentation and microbiota in dairy cows fed a diet containing low quality forage. J. Anim. Sci. Biotechnol. 2017, 8, 36. [Google Scholar] [CrossRef]
- Zhang, H.; Tong, J.; Zhang, Y.; Xiong, B.; Jiang, L. Metabolomics reveals potential biomarkers in the rumen fluid of dairy cows with different levels of milk production. Anim. Biosci. 2020, 33, 79–90. [Google Scholar] [CrossRef]
- Chahin, A.; Guiavarc’h, Y.P.; Dziurla, M.-A.; Toussaint, H.; Feidt, C.; Rychen, G. 1-Hydroxypyrene in milk and urine as a bioindicator of polycyclic aromatic hydrocarbon exposure of ruminants. J. Agric. Food Chem. 2008, 56, 1780–1786. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.D.; Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.E.; Powell, J.L.; Drewnoski, M.E.; O’Neil, M.; Beitz, D.C.; Weiss, W.P. Vitamin D status of dairy cattle: Outcomes of current practices in the dairy industry. J. Dairy Sci. 2016, 99, 10150–10160. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Fink-Gremmels, J. Mycotoxin syndrome in dairy cattle: Characterisation and intervention results. World Mycotoxin J. 2014, 7, 357–366. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Giamouri, E.; Tavrizelou, S.; Zacharioudaki, M.; Danezis, G.; Simitzis, P.E.; Zoidis, E.; Tsiplakou, E.; Pappas, A.C.; Georgiou, C.A.; et al. Impact of mycotoxins on animals’ oxidative status. Antioxidants 2021, 10, 214. [Google Scholar] [CrossRef]
- Fujihara, T.; Shem, M.N. Metabolism of microbial nitrogen in ruminants with special reference to nucleic acids. Anim. Sci. J. 2011, 82, 198–208. [Google Scholar] [CrossRef]
- Stentoft, C.; Røjen, B.A.; Jensen, S.K.; Kristensen, N.B.; Vestergaard, M.; Larsen, M. Absorption and intermediary metabolism of purines and pyrimidines in lactating dairy cows. Br. J. Nutr. 2015, 113, 560–573. [Google Scholar] [CrossRef] [Green Version]
- Schager, W.M.; Harrison, J.H.; Gaskins, C.T.; Davidson, D. Factors affecting application of milk allantoin as an estimator of microbial protein flow to the duodenum under commercial conditions. J. Dairy Sci. 2003, 86, 1716–1721. [Google Scholar] [CrossRef]
- Gallo, A.; Minuti, A.; Bani, P.; Bertuzzi, T.; Piccioli Cappelli, F.; Doupovec, B.; Faas, J.; Schatzmayr, D.; Trevisi, E. A mycotoxin-deactivating feed additive counteracts the adverse effects of regular levels of Fusarium mycotoxins in dairy cows. J. Dairy Sci. 2020, 103, 11314–11331. [Google Scholar] [CrossRef] [PubMed]
- Bellassi, P.; Rocchetti, G.; Nocetti, M.; Lucini, L.; Masoero, F.; Morelli, L. A combined metabolomic and metagenomic approach to discriminate raw milk for the production of hard cheese. Foods 2021, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.; Kwok, L.-Y.; Wang, Y.; Ma, C.; Mi, Z.; Zhang, H. Ultra-performance liquid chromatography-quadrupole-time of flight mass spectrometry MSE-based untargeted milk metabolomics in dairy cows with subclinical or clinical mastitis. J. Dairy Sci. 2017, 100, 4884–4896. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Rebecchi, A.; Dallolio, M.; Braceschi, G.; Domínguez, R.; Dallolio, G.; Trevisan, M.; Lorenzo, J.M.; Lucini, L. Changes in the chemical and sensory profile of ripened Italian salami following the addition of different microbial starters. Meat Sci. 2021, 180, 108584. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-Dial: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods. 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen rearrangement rules: Computational MS/MS fragmentation and structure elucidation using MS-Finder software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Senizza, A.; Rocchetti, G.; Callegari, M.L.; Lucini, L.; Morelli, L. Linoleic acid induces metabolic stress in the intestinal microorganism Bifidobacterium breve DSM 20213. Sci. Rep. 2020, 10, 5997. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Unit | Mean | S.D. | Q25 | Q75 |
|---|---|---|---|---|---|
| Herd composition and characteristics | |||||
| Lactating cows | n | 199.7 | 138.2 | 94.0 | 270.0 |
| Dry cows | n | 27.9 | 25.8 | 10.0 | 40.0 |
| Calves | n | 193.6 | 140.4 | 94.0 | 264.0 |
| Milk yield and quality | |||||
| Milk yield | Kg/cow/day | 32.3 | 4.6 | 29.8 | 35.2 |
| 4% Fat corrected milk | Kg/cow/day | 33.40 | 4.92 | 30.16 | 36.30 |
| Energy corrected milk | Kg/cow/day | 35.50 | 5.14 | 32.23 | 38.75 |
| Milk fat | % | 4.03 | 0.17 | 3.94 | 4.13 |
| Milk protein | % | 3.50 | 0.10 | 3.44 | 3.55 |
| Milk casein | % | 2.72 | 0.09 | 2.68 | 2.77 |
| SCC | log10 cells/mL × 1000 | 5.23 | 0.16 | 5.11 | 5.34 |
| Dry matter intake, dietary composition a and ingredient inclusion | |||||
| Dry matter intake | Kg DM/cow/day | 23.76 | 2.24 | 22.79 | 25.49 |
| Net energy for lactation 3x b | Mcal/kg DM | 1.63 | 0.04 | 1.61 | 1.66 |
| CP | % DM | 16.42 | 1.04 | 15.75 | 17.19 |
| Soluble CP | % DM | 4.77 | 0.37 | 4.57 | 5.02 |
| NDF | % DM | 29.85 | 3.26 | 27.65 | 31.38 |
| ADF | % DM | 19.12 | 2.43 | 17.71 | 20.16 |
| NFC | % DM | 41.67 | 2.55 | 39.55 | 43.62 |
| Starch | % DM | 27.46 | 3.41 | 25.99 | 29.78 |
| Corn silage c | % | 30.51 | 5.84 | 25.98 | 35.64 |
| Cluster | ||||||
|---|---|---|---|---|---|---|
| Items | Units | 1 | 2 | 3 | 4 | 5 |
| Dry matter | % | 34.5 | 34.4 | 37.9 | 37.4 | 36.2 |
| pH | 3.69 | 3.77 | 3.84 | 3.80 | 3.89 | |
| 1,2-propanediol | % DM | 0.57 | 0.42 | 0.73 | 0.51 | 0.19 |
| Acetic acid | % DM | 3.38 | 3.24 | 3.28 | 2.87 | 3.03 |
| Propionic acid | % DM | 0.12 | 0.16 | 0.04 | 0.07 | 0.25 |
| Butyric acid | % DM | 0.001 | 0.003 | 0.001 | 0.003 | 0.003 |
| Lactic acid | % DM | 3.33 | 4.29 | 1.91 | 3.47 | 2.51 |
| Mycotoxins concentrations a and incidence | ||||||
| Zearalenone | µg/kg DM | 4.61 ± 2.93 | 28.84 ± 45.20 | 7.11 ± 4.05 | 2.93 ± 1.69 | 1.63 ± 0.34 |
| Incidence | 33.3% | 18.8% | 100.0% | 20.0% | 66.7% | |
| Deoxynivalenol | µg/kg DM | 43.65 ± 46.79 | 25.59 ± 39.08 | 151.67 ± 8.89 | 32.33 ± 26.77 | 96.87 ± 130.93 |
| Incidence | 61.1% | 68.8% | 100.0% | 100% | 66.7% | |
| Fumonisin B1 | µg/kg DM | 105.72 ± 110.69 | 210.10 ± 214.76 | 181.80 ± 123.04 | 268.15 ± 131.28 | 1576.64 ± 265.53 |
| Incidence | 94.4% | 100% | 100% | 100% | 100% | |
| Fumonisin B2 | µg/kg DM | 27.37 ± 33.70 | 68.49 ± 83.85 | 139.13 ± 187.23 | 69.90 ± 17.84 | 404.73 ± 50.32 |
| Incidence | 94.4% | 93.8% | 100% | 100% | 100% | |
| Fumonisin B3 | µg/kg DM | 11.40 ± 8.47 | 18.72 ± 16.69 | 69.00 ± 59.72 | 24.97 ± 6.05 | 232.54 ± 120.66 |
| Incidence | 66.7% | 100% | 100% | 100% | 100% | |
| Moniliformin | µg/kg DM | 12.47 ± 13.36 | 7.12 ± 7.73 | 39.90 ± 5.61 | 12.97 ± 9.04 | 36.22 ± 34.38 |
| Incidence | 94.4% | 93.8% | 100% | 100% | 100% | |
| Fusaric acid | µg/kg DM | 165.93 ± 92.69 | 210.62 ± 105.41 | 195.48 ± 88.71 | 209.49 ± 174.44 | 375.11 ± 408.74 |
| Incidence | 100% | 100% | 100% | 100% | 100% | |
| Sum of Aspergillus toxins | µg/kg DM | 161.52 ± 143.74 | 93.09 ± 62.34 | 565.23 ± 230.06 | 46.40 ± 33.42 | 236.22 ± 190.16 |
| Incidence | 100% | 100% | 100% | 100% | 100% | |
| Sum of Alternaria toxins | µg/kg DM | 7.71 ± 9.86 | 1.35 ± 4.28 | 18.65 ± 13.87 | 3.65 ± 8.15 | 38.39 ± 29.49 |
| Incidence | 100% | 100% | 100% | 100% | 100% | |
| Sum of Fusarium b toxins | µg/kg DM | 230.19 ± 99.40 | 716.42 ± 117.73 | 619.71 ± 99.76 | 1567.17 ± 340.46 | 739.80 ± 445.34 |
| Incidence | 100% | 100% | 100% | 100% | 100% | |
| Sum of Penicillium toxins | µg/kg DM | 162.23 ± 138.24 | 96.78 ± 93.55 | 708.24 ± 116.77 | 80.51 ± 41.46 | 189.66 ± 139.68 |
| Incidence | 100% | 100% | 100% | 100% | 100% | |
| Superclass | Most Discriminant Compound | LogFC Cluster 3 vs. Cluster 1 | LogFC Cluster 3 vs. Cluster 2 | LogFC Cluster 4 vs. Cluster 1 | LogFC Cluster 4 vs. Cluster 2 | LogFC Cluster 5 vs. Cluster 1 | LogFC Cluster 5 vs. Cluster 2 |
|---|---|---|---|---|---|---|---|
| Alkaloids and derivatives | Ecgonine (VIP score: 1.54; p-value: 2.55 × 10−4) | 0.061 | −0.020 | 0.526 | 0.445 | −1.031 | −1.112 |
| Amines | Norspermidine (VIP score:1.57; p-value: 2.82 × 10−5) | −0.342 | −0.232 | 0.052 | 0.162 | 0.583 | 0.692 |
| Amino acids and peptides | O-Succinyl-L-homoserine (VIP score: 1.78; p-value: 5.36 × 10−5) | 0.144 | 0.325 | −0.199 | -0.018 | 0.775 | 0.956 |
| Benzenoids | 1-Hydroxypyrene glucuronide (VIP score: 1.74; p-value: 2.25 × 10−4) | 0.139 | 0.198 | −0.278 | −0.219 | 0.447 | 0.507 |
| Diazines | Aripiprazole (VIP score: 1.49; p-value: 1.93 × 10−3) | −0.227 | −0.127 | −0.759 | −0.659 | 0.989 | 1.089 |
| Fatty Acyls | Nephritogenoside (VIP score: 1.29; p-value: 2.83 × 10−5) | −0.200 | −0.174 | −0.160 | −0.135 | −0.119 | −0.093 |
| Glycerolipids | TG(15:0/24:0/24:1(15Z)) (VIP score: 1.40; p-value: 1.67 × 10−10) | 0.528 | 0.435 | 0.297 | 0.204 | 0.728 | 0.635 |
| Glycerophospholipids | DG(18:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) (VIP score: 1.42; p-value: 1.67 × 10−12) | 0.172 | 0.088 | 0.191 | 0.107 | −0.0002 | −0.085 |
| Indoles and derivatives | Indoleacetic acid (VIP score: 1.79; p-value: 4.49 × 10−5) | 0.043 | 0.156 | 0.135 | 0.249 | 0.169 | 0.282 |
| Keto acids and derivatives | 4-Fumarylacetoacetic acid (VIP score: 1.03; p-value: 1.01 × 10−2) | −0.512 | −0.268 | 0.105 | 0.349 | 0.475 | 0.718 |
| Nucleosides, nucleotides, and analogues | 8-Oxo-dGMP (VIP score: 1.19; p-value: 3.69 × 10−5) | −0.459 | −0.496 | 0.673 | 0.635 | −1.556 | −1.593 |
| Organic acids and derivatives | Taurine (VIP score: 1.25; p-value: 6.19 × 10−7) | 1.130 | 1.437 | −0.152 | 0.1544 | 1.817 | 2.124 |
| Organooxygen compounds | Chitobiose (VIP score: 1.97; p-value: 2.08 × 10−3) | 0.810 | 0.767 | 0.134 | 0.092 | 0.696 | 0.654 |
| Polyphenols | Troxerutin (VIP score: 1.42; p-value: 5.69 × 10−10) | 0.010 | 0.389 | −0.223 | 0.155 | 0.044 | 0.423 |
| Prenol lipids | Farnesol (VIP score: 1.26; p-value: 1.19 × 10−2) | −0.244 | −0.348 | 0.573 | 0.468 | −1.260 | −1.365 |
| Pteridines and derivatives | Pteroyl-D-glutamic acid (VIP score: 1.21; p-value: 2.08 × 10−4) | 0.734 | 0.900 | −0.214 | −0.048 | 1.546 | 1.712 |
| Purine nucleotides | ADP-ribose 2′-phosphate (VIP score: 1.31; p-value: 1.19 × 10−5) | 1.496 | 2.329 | −0.332 | 0.501 | 2.810 | 3.644 |
| Purine nucleosides | Guanosine (VIP score: 1.34; p-value: 2.82 × 10−4) | 0.820 | 0.696 | 0.987 | 0.863 | 0.426 | 0.302 |
| Pyridines and derivatives | N-Methylnicotinium (VIP score: 0.99; p-value: 8.94 × 10−5) | −0.454 | −0.961 | −0.070 | −0.576 | −1.718 | −2.225 |
| Pyrimidine nucleosides | Thymidine (VIP score: 1.09; p-value: 5.41 × 10−3) | −0.602 | −0.886 | −0.738 | −1.022 | −0.198 | −0.481 |
| Pyrimidine nucleotides | dUMP (VIP score: 1.29; p-value: 2.93 × 10−6) | 1.496 | 2.329 | −0.332 | 0.501 | 2.810 | 3.644 |
| Imidazopyrimidines | Hypoxanthine (VIP score: 1.04; p-value: 1.61 × 10−3) | −0.480 | -0.409 | 0.054 | 0.125 | −0.039 | 0.031 |
| Quinolines and derivatives | Salsoline-1-carboxylate (VIP score: 1.05; p-value: 2.98 × 10−3) | −0.317 | −0.237 | −0.594 | −0.515 | 0.068 | 0.148 |
| Sphingolipids | Phosphatidylinositol-3,4,5-trisphosphate (VIP score: 1.34; p-value: 1.47 × 10−3) | 0.552 | 0.595 | 0.362 | 0.405 | 0.587 | 0.630 |
| Steroids and steroid derivatives | Vitamin D2 3-glucuronide (VIP score: 1.42; p-value: 1.45 × 10−13) | −0.135 | 0.120 | −0.179 | 0.077 | 0.554 | 0.809 |
| Tetrapyrroles and derivatives | Mesoporphyrin IX (VIP score: 1.55; p-value: 1.33 × 10−15) | 0.859 | 1.184 | −0.578 | −0.254 | 2.706 | 3.031 |
| Other metabolites | Choline (VIP score: 1.25; p-value: 2.12 × 10−4) | −0.711 | −0.219 | −0.748 | −0.256 | 0.205 | 0.697 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocchetti, G.; Ghilardelli, F.; Bonini, P.; Lucini, L.; Masoero, F.; Gallo, A. Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows. Metabolites 2021, 11, 475. https://doi.org/10.3390/metabo11080475
Rocchetti G, Ghilardelli F, Bonini P, Lucini L, Masoero F, Gallo A. Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows. Metabolites. 2021; 11(8):475. https://doi.org/10.3390/metabo11080475
Chicago/Turabian StyleRocchetti, Gabriele, Francesca Ghilardelli, Paolo Bonini, Luigi Lucini, Francesco Masoero, and Antonio Gallo. 2021. "Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows" Metabolites 11, no. 8: 475. https://doi.org/10.3390/metabo11080475
APA StyleRocchetti, G., Ghilardelli, F., Bonini, P., Lucini, L., Masoero, F., & Gallo, A. (2021). Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows. Metabolites, 11(8), 475. https://doi.org/10.3390/metabo11080475