
Acute Administration of Exogenous Lactate Increases Carbohydrate Metabolism during Exercise in Mice


Abstract
:1. Introduction
2. Results
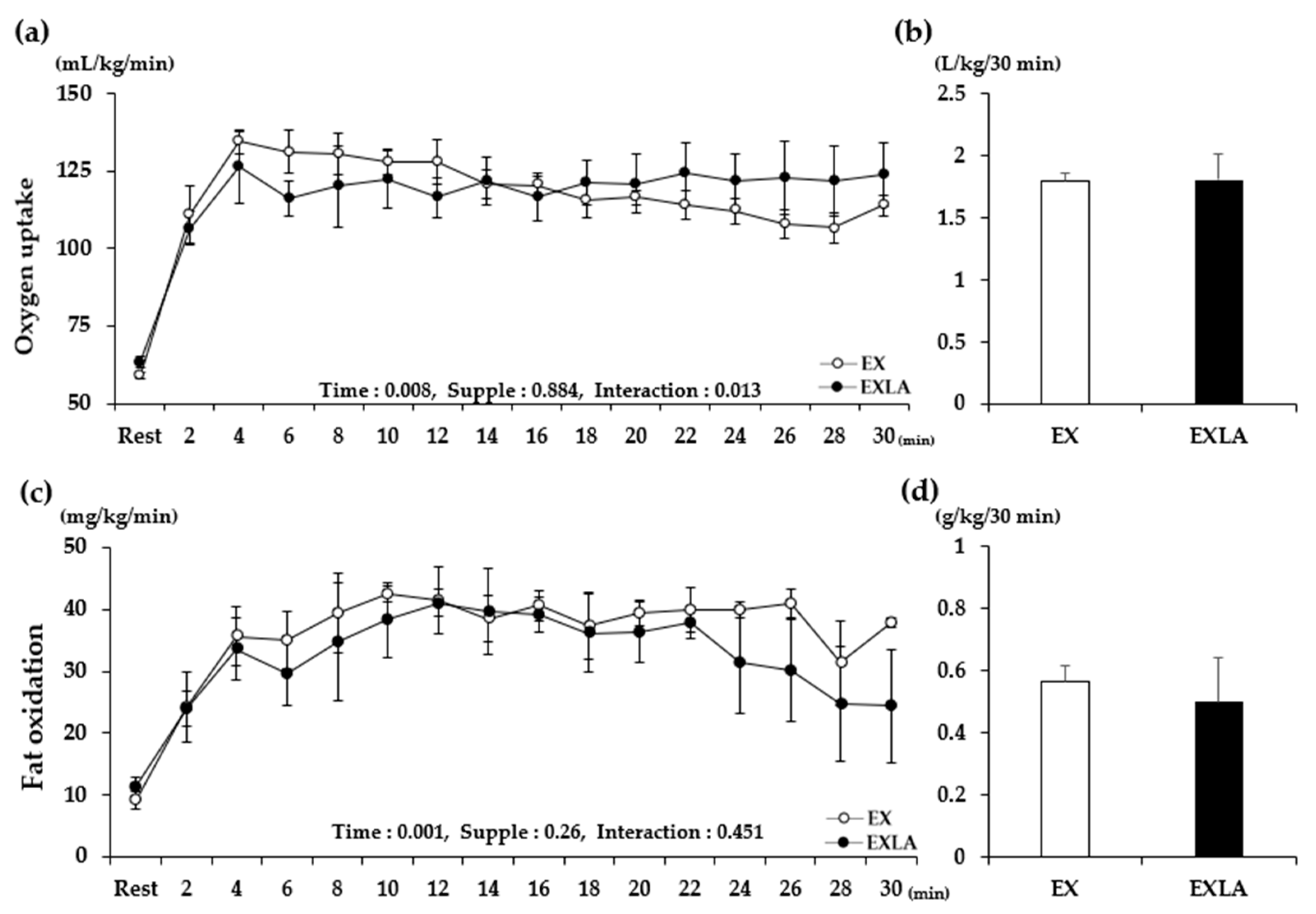
2.1. Energy Metabolism during Exercise
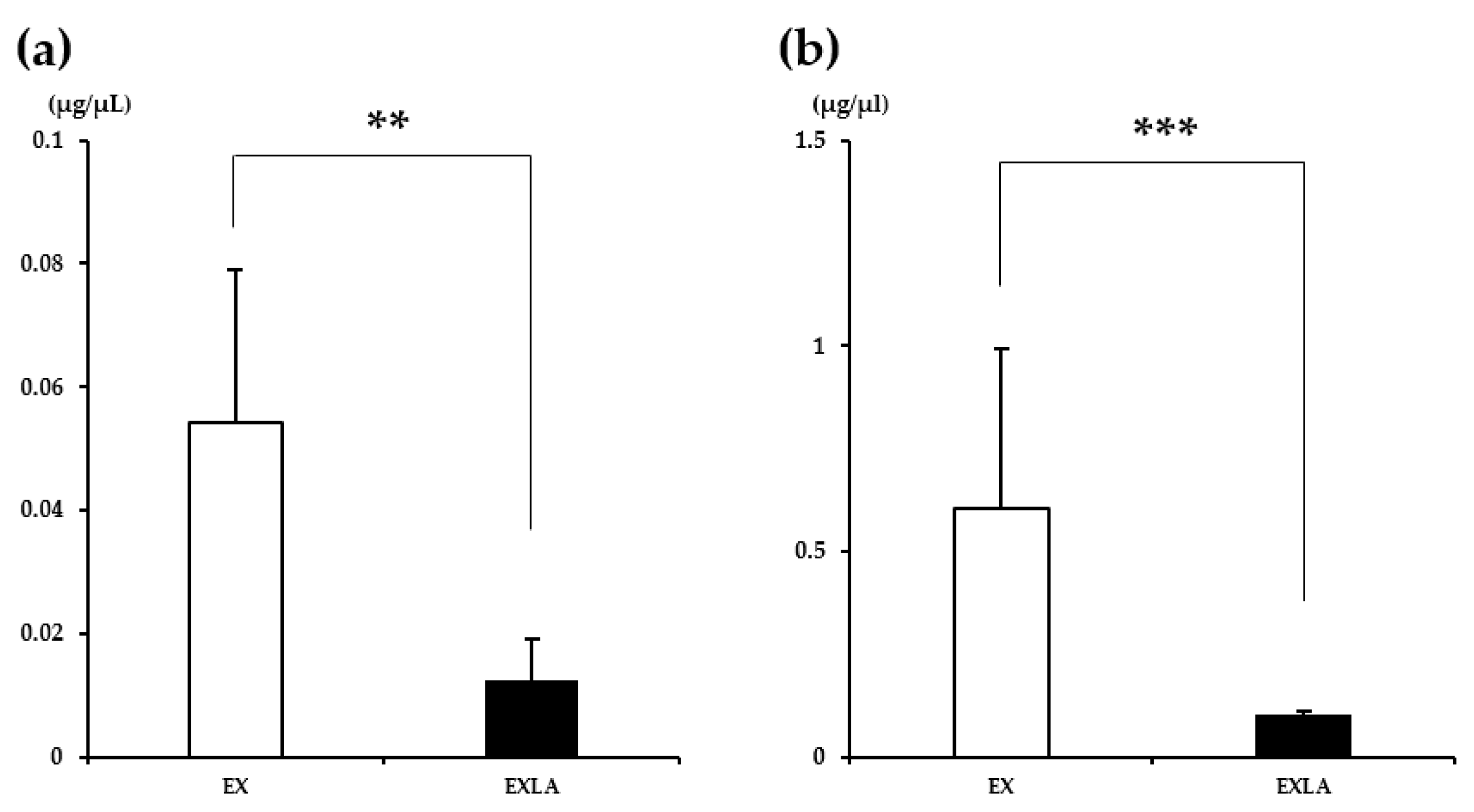
2.2. Glycogen Content
2.3. Blood Analysis-Whole Blood
2.4. Blood Analysis-Serum
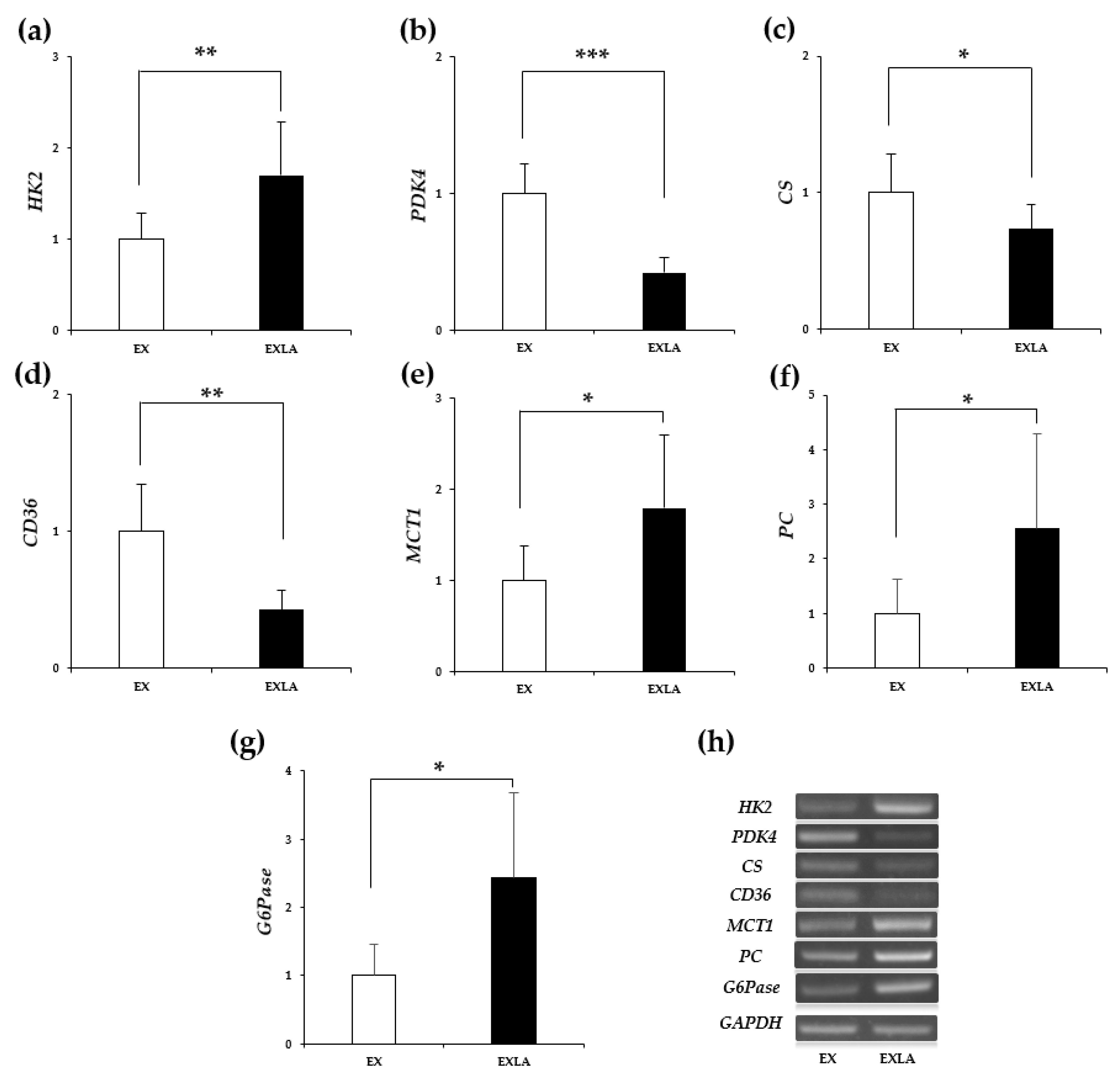
2.5. mRNA Expression of Energy Metabolism-Related Factors
3. Discussion
4. Materials and Methods
4.1. Animal Care
4.2. Study Design
4.3. Metabolic Analysis during Exercise
4.4. Determination of Blood Parameters
4.5. Glycogen Content Analysis
4.6. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Park, J.; Lim, K. Nutrition Supplements to Stimulate Lipolysis: A Review in Relation to Endurance Exercise Capacity. J. Nutr. Sci. Vitaminol. 2016, 62, 141–161. [Google Scholar] [CrossRef] [Green Version]
- Jeukendrup, A.E.; Randell, R. Fat burners: Nutrition supplements that increase fat metabolism. Obes. Rev. 2011, 12, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.O.; Yeo, N.H.; Kang, S. Autonomic Nervous Activity and Lipid Oxidation Postexercise with Capsaicin in the Humans. J. Sports Sci. Med. 2010, 9, 253–261. [Google Scholar] [PubMed]
- Ruiz-Moreno, C.; Gutiérrez-Hellín, J.; Amaro-Gahete, F.J.; González-García, J.; Giráldez-Costas, V.; Pérez-García, V.; Del Coso, J. Caffeine increases whole-body fat oxidation during 1 h of cycling at Fatmax. Eur. J. Nutr. 2021, 60, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Stephens, F.B.; Galloway, S.D. Carnitine and fat oxidation. Limits Hum. Endur. 2013, 76, 13–23. [Google Scholar]
- Lejeune, M.P.G.M.; Kovacs, E.M.R.; Westerterp-Plantenga, M.S. Effect of capsaicin on substrate oxidation and weight maintenance after modest body-weight loss in human subjects. Br. J. Nutr. 2003, 90, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, W.M.; Hopkins, F.G. Lactic acid in amphibian muscle1. J. Physiol. 1907, 35, 247–309. [Google Scholar] [CrossRef]
- Sahlin, K. Metabolic Factors in Fatigue. Sports Med. 1992, 13, 99–107. [Google Scholar] [CrossRef]
- Donaldson, S.K.B.; Hermansen, L.; Bolles, L. Differential, direct effects of H+ on Ca2+-activated force of Skinned fibers from the soleus, cardiac and adductor magnus muscles of rabbits. Pflügers Arch. 1978, 376, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Nalbandian, M.; Takeda, M. Lactate as a Signaling Molecule That Regulates Exercise-Induced Adaptations. Biology 2016, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Hollidge-Horvat, M.G.; Parolin, M.L.; Wong, D.; Jones, N.L.; Heigenhauser, G.J.F. Effect of induced metabolic alkalosis on human skeletal muscle metabolism during exercise. Am. J. Physiol. Metab. 2000, 278, E316–E329. [Google Scholar] [CrossRef] [Green Version]
- Cooke, R.; Franks, K.; Luciani, G.B.; Pate, E. The inhibition of rabbit skeletal muscle contraction by hydrogen ions and phosphate. J. Physiol. 1988, 395, 77–97. [Google Scholar] [CrossRef]
- Van Hall, G.; Stømstad, M.; Rasmussen, P.; Jans, Ø.; Zaar, M.; Gam, C.; Quistorff, B.; Secher, N.H.; Nielsen, H.B. Blood Lactate is an Important Energy Source for the Human Brain. Br. J. Pharmacol. 2009, 29, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.; Fattor, J.A.; Jacobs, K.; Horning, M.A.; Navazio, F.; Lindinger, M.; Brooks, G.A. Lactate and glucose interactions during rest and exercise in men: Effect of exogenous lactate infusion. J. Physiol. 2002, 544, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Intra- and extra-cellular lactate shuttles. Med. Sci. Sports Exerc. 2000, 32, 790–799. [Google Scholar] [CrossRef]
- Cori, C.F.; Cori, G.T. Carbohydrate metabolism. Annu. Rev. Biochem. 1946, 15, 193–218. [Google Scholar] [CrossRef]
- Stevenson, R.W.; Mitchell, D.R.; Hendrick, G.K.; Rainey, R.; Cherrington, A.D.; Frizzell, R.T. Lactate as substrate for glycogen resynthesis after exercise. J. Appl. Physiol. 1987, 62, 2237–2240. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P.; Wilson, M.C. The monocarboxylate transporter family-Role and regulation. IUBMB Life 2011, 64, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Cell-cell and intracellular lactate shuttles. J. Physiol. 2009, 587, 5591–5600. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, Y.; Takeda, K.; Tamura, Y.; Hatta, H. Lactate administration increases mRNA expression of PGC-1α and UCP3 in mouse skeletal muscle. Appl. Physiol. Nutr. Metab. 2016, 41, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Kitaoka, Y.; Yamamoto, K.; Matsunaga, Y.; Hatta, H. Oral Lactate Administration Additively Enhances Endurance Training-Induced Increase in Cytochrome C Oxidase Activity in Mouse Soleus Muscle. Nutrients 2020, 12, 770. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, D.; Hanawa, T.; Takahashi, Y.; Masuda, H.; Kato, M.; Hatta, H. Chronic Post-Exercise Lactate Administration with Endurance Training Increases Glycogen Concentration and Monocarboxylate Transporter 1 Protein in Mouse White Muscle. J. Nutr. Sci. Vitaminol. 2014, 60, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyun, S.; Yoo, C.; Park, H.Y.; Kim, J.; Lim, K. The Effects of Exogenous Lactate Administration on the IGF1/Akt/mTOR Pathway in Rat Skeletal Muscle. Int. J. Environ. Res. Public Health 2020, 17, 7805. [Google Scholar] [CrossRef]
- Kyun, S.; Yoo, C.; Hashimoto, T.; Tomi, H.; Teramoto, N.; Kim, J.; Lim, K. Effects of exogenous lactate administration on fat metabolism and glycogen synthesis factors in rats. J. Exerc. Nutr. Biochem. 2020, 24, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.E.; Schifino, A.G.; Leong, C.-H. Effect of lactate supplementation on VO2peak and onset of blood lactate accumulation: A double-blind, placebo-controlled trial. Acta Gymnica 2019, 49, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the exercise intensity that elicits maximal fat oxidation. Med. Sci. Sports Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.E.; Richter, E.A. Regulation of glucose and glycogen metabolism during and after exercise. J. Physiol. 2012, 590, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.T.; Fuchs, C.J.; Betts, J.A.; Van Loon, L.J. Liver glycogen metabolism during and after prolonged endurance-type exercise. Am. J. Physiology-Endocrinol. Metab. 2016, 311, E543–E553. [Google Scholar] [CrossRef]
- Brooks, G.A. Lactate as a fulcrum of metabolism. Redox Biol. 2020, 35, 101454. [Google Scholar] [CrossRef]
- Takahashi, K.; Kitaoka, Y.; Matsunaga, Y.; Hatta, H. Effects of lactate administration on mitochondrial enzyme activity and monocarboxylate transporters in mouse skeletal muscle. Physiol. Rep. 2019, 7, e14224. [Google Scholar] [CrossRef]
- Yin, X.; Choudhury, M.; Kang, J.-H.; Schaefbauer, K.J.; Jung, M.-Y.; Andrianifahanana, M.; Hernandez, D.M.; Leof, E.B. Hexokinase 2 couples glycolysis with the profibrotic actions of TGF-β. Sci. Signal. 2019, 12, eaax4067. [Google Scholar] [CrossRef]
- Robey, R.B.; Hay, N. Mitochondrial Hexokinases: Guardians of the Mitochondria. Cell Cycle 2005, 4, 654–658. [Google Scholar] [CrossRef]
- Owen, O.E.; Kalhan, S.; Hanson, R.W. The Key Role of Anaplerosis and Cataplerosis for Citric Acid Cycle Function. J. Biol. Chem. 2002, 277, 30409–30412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, P.M.; Donley, D.A.; Bryner, R.W.; Alway, S.E. Citrate synthase expression and enzyme activity after endurance training in cardiac and skeletal muscles. J. Appl. Physiol. 2003, 94, 555–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaoka, Y.; Takahashi, Y.; Machida, M.; Takeda, K.; Takemasa, T.; Hatta, H. Effect of AMPK activation on monocarboxylate transporter (MCT)1 and MCT4 in denervated muscle. J. Physiol. Sci. 2013, 64, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lim, K. Relationship between FAT/CD36 Protein in Skeletal Muscle and Whole-body Fat Oxidation in Endurance-trained Mice. J. Exerc. Nutr. Biochem. 2016, 20, 48–52. [Google Scholar] [CrossRef]
- Spriet, L.L.; Watt, M.J. Regulatory mechanisms in the interaction between carbohydrate and lipid oxidation during exercise. Acta Physiol. Scand. 2003, 178, 443–452. [Google Scholar] [CrossRef]
- Exton, J. Gluconeogenesis. Metabolism 1972, 21, 945–990. [Google Scholar] [CrossRef]
- Kwon, I. Protective effects of endurance exercise on skeletal muscle remodeling against doxorubicin-induced myotoxicity in mice. J. Exerc. Nutr. Biochem. 2020, 24, 11–21. [Google Scholar] [CrossRef]
- Lezi, E.; Lu, J.; Selfridge, J.E.; Burns, J.; Swerdlow, R.H. Lactate administration reproduces specific brain and liver exercise-related changes. J. Neurochem. 2013, 127, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.; Seo, J.-B.; Kim, J.; Lim, K. Effect of mild-intensity exercise training with capsiate intake on fat deposition and substrate utilization during exercise in diet-induced obese mice. Phys. Act. Nutr. 2020, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shihara, K.; Taniguchi, H. Fat max as an index of aerobic exercise performance in mice during uphill running. PLoS ONE 2018, 13, e0193470. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EX | EXLA | |
|---|---|---|
| Glucose (mg/dL) | 158.5 ± 26 | 65.5 ± 27.2 *** |
| Triglyceride (mg/dL) | 145.9 ± 13.3 | 92.3 ± 15.7 *** |
| Lactate (mmol/L) | 7.4 ± 1.8 | 10.8 ± 4.2 |
| EX | EXLA | |
|---|---|---|
| Glycerol (mmol/L) | 0.51 ± 0.05 | 0.4 ± 0.08 ** |
| Free fatty acid (mmol/L) | 0.59 ± 0.16 | 0.37 ± 0.07 ** |
| Gene | Sequence |
|---|---|
| HK2 | F-5′ ATC GCC GGA TTG GAA CAG AA 3′ |
| R-5′CTC CGT GAA TAA GCA GGC GA 3′ | |
| PDK4 | F-5′ CGC CTG GCC AAT ATC CTG AA 3′ |
| R-5′ GCC TTG AGC CAT TGT AGG GA 3′ | |
| CS | F-5′ CAA GTC ATC TAC GCC AGG GAC A 3′ |
| R-5′ CAA AGC GTC TCC AGC TAA CCA AG 3′ | |
| CD36 | F-5′ GGC CAA GCT ATT GCG ACA T 3′ |
| R-5′ CAG ATC CGA ACA CAG CGT AGA 3′ | |
| MCT1 | F-5′ GGC CTG AGC AAG TCA AGC TA 3′ |
| R-5′ GCA AAT CCA AAG ACT CCG GC 3′ | |
| PC | F-5′ GCA GCC TTT GGG AAT GGA 3′ |
| R-5′ GGT GAG ACG TGA GCG AAG TTG 3′ | |
| G6Pase | F-5′ CAG AAT GGG TCC ACC TTG ACA 3′ |
| R-5′ GGG CTT CAG AGA GTC AAA GAG ATG 3′ | |
| GAPDH | F-5′ AAC TTT GGC ATT GTG GAA GG 3′ |
| R-5′ ACA CAT TGG GGG TAG GAA CA 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, I.; Kim, J.; Kyun, S.; Hwang, D.; Lim, K. Acute Administration of Exogenous Lactate Increases Carbohydrate Metabolism during Exercise in Mice. Metabolites 2021, 11, 553. https://doi.org/10.3390/metabo11080553
Jang I, Kim J, Kyun S, Hwang D, Lim K. Acute Administration of Exogenous Lactate Increases Carbohydrate Metabolism during Exercise in Mice. Metabolites. 2021; 11(8):553. https://doi.org/10.3390/metabo11080553
Chicago/Turabian StyleJang, Inkwon, Jisu Kim, Sunghwan Kyun, Deunsol Hwang, and Kiwon Lim. 2021. "Acute Administration of Exogenous Lactate Increases Carbohydrate Metabolism during Exercise in Mice" Metabolites 11, no. 8: 553. https://doi.org/10.3390/metabo11080553
APA StyleJang, I., Kim, J., Kyun, S., Hwang, D., & Lim, K. (2021). Acute Administration of Exogenous Lactate Increases Carbohydrate Metabolism during Exercise in Mice. Metabolites, 11(8), 553. https://doi.org/10.3390/metabo11080553