
Increased Fibroblast Growth Factor 21 (FGF21) Concentration in Early Second Trimester Amniotic Fluid and Its Association with Fetal Growth

 ,
,  ,
, 
Abstract
:1. Introduction
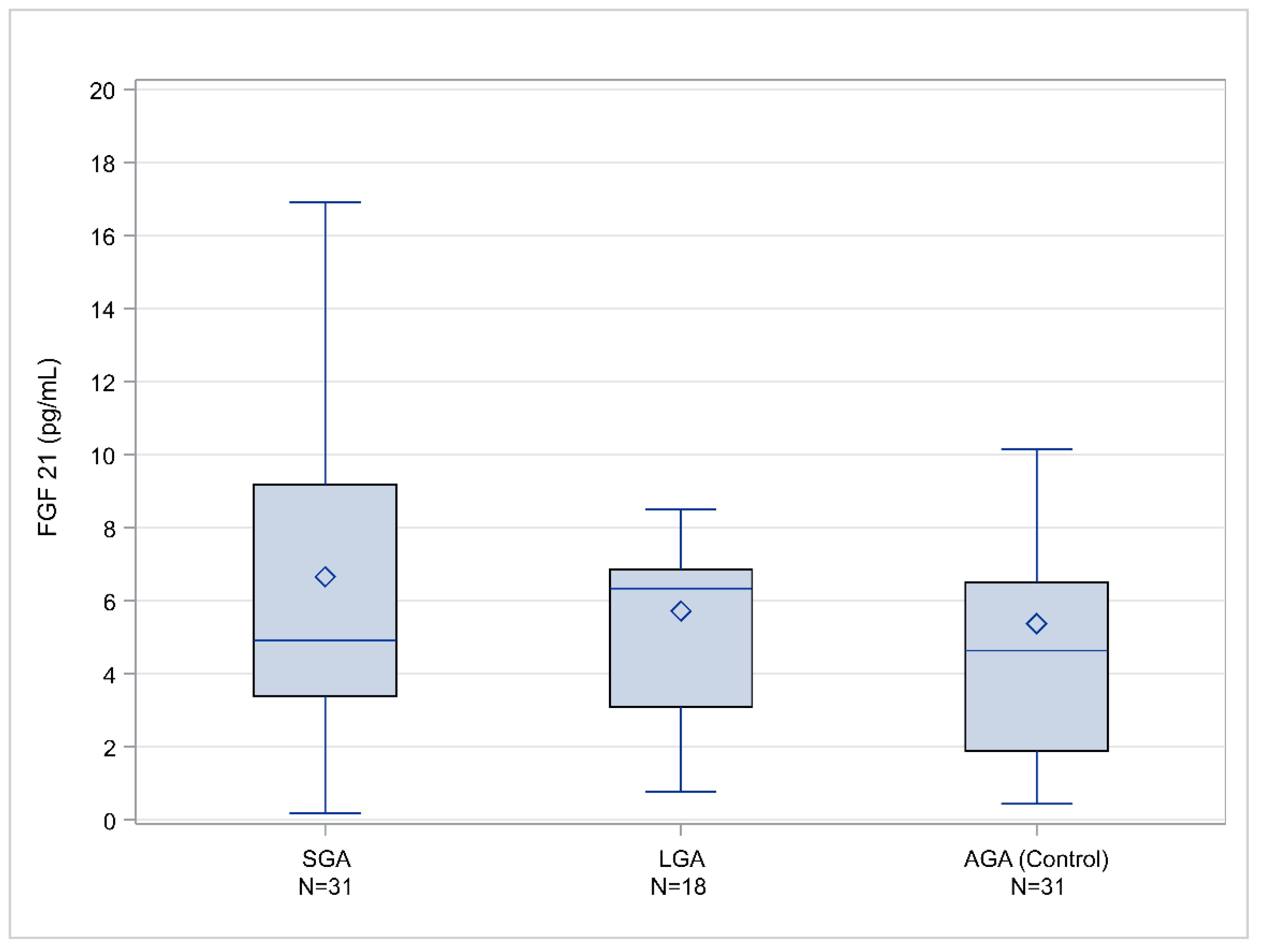
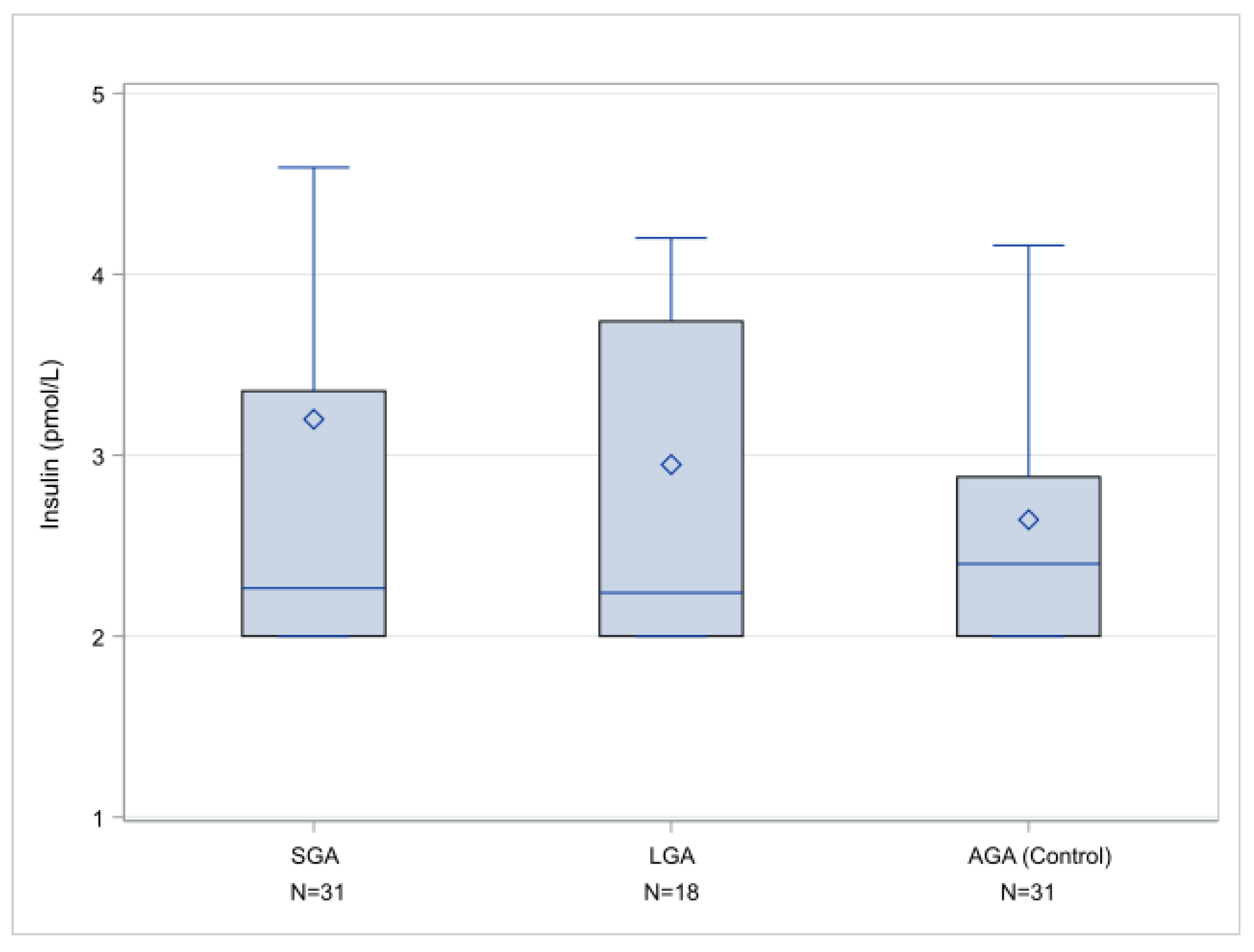
2. Results
3. Discussion
4. Materials and Methods
4.1. Groups
4.2. Measurements
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vasak, B.; Koenen, S.V.; Koster, M.P.H.; Hukkelhoven, C.; Franx, A.; Hanson, M.A.; Visser, G.H.A. Human fetal growth is constrained below optimal for perinatal survival. Ultrasound Obstet. Gynecol. 2015, 45, 162–167. [Google Scholar] [CrossRef]
- Vrachnis, N.; Botsis, D.; Iliodromiti, Z. The Fetus That Is Small for Gestational Age. Ann. N. Y. Acad. Sci. 2006, 1092, 304–309. [Google Scholar] [CrossRef]
- Boulet, S.; Alexander, G.R.; Salihu, H.M.; Pass, M. Macrosomic births in the united states: Determinants, outcomes, and proposed grades of risk. Am. J. Obstet. Gynecol. 2003, 188, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.G.; Beall, M.H. Adult Sequelae of Intrauterine Growth Restriction. Semin. Perinatol. 2008, 32, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lees, C.; Marlow, N.; Arabin, B.; Bilardo, C.M.; Brezinka, C.; Derks, J.B.; Duvekot, J.; Frusca, T.; Diemert, A.; Ferrazzi, E.; et al. Perinatal morbidity and mortality in early-onset fetal growth restriction: Cohort outcomes of the trial of randomized umbilical and fetal flow in Europe (TRUFFLE). Ultrasound Obstet. Gynecol. 2013, 42, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Vitoratos, N.; Iliodromiti, Z.; Sifakis, S.; Deligeoroglou, E.; Creatsas, G. Intrauterine inflammation and preterm delivery. Ann. N. Y. Acad. Sci. 2010, 1205, 118–122. [Google Scholar] [CrossRef]
- Savchev, S.; Figueras, F.; Cruz-Martinez, R.; Illa, M.; Botet, F.; Gratacos, E. Estimated weight centile as a predictor of perinatal outcome in small-for-gestational-age pregnancies with normal fetal and maternal Doppler indices. Ultrasound Obstet. Gynecol. 2012, 39, 299–303. [Google Scholar] [CrossRef]
- Figueras, F.; Savchev, S.; Triunfo, S.; Crovetto, F.; Gratacos, E. An integrated model with classification criteria to predict small-for-gestational-age fetuses at risk of adverse perinatal outcome. Ultrasound Obstet. Gynecol. 2015, 45, 279–285. [Google Scholar] [CrossRef]
- Vrachnis, N.; Kalampokas, E.; Sifakis, S.; Vitoratos, N.; Kalampokas, T.; Botsis, D.; Iliodromiti, Z. Placental growth factor (PlGF): A key to optimizing fetal growth. J. Matern. Neonatal Med. 2013, 26, 995–1002. [Google Scholar] [CrossRef]
- Mifsud, W.; Sebire, N.J. Placental Pathology in Early-Onset and Late-Onset Fetal Growth Restriction. Fetal Diagn. Ther. 2014, 36, 117–128. [Google Scholar] [CrossRef]
- Gerretsen, G.; Huisjes, H.J.; Elema, J.D. Morphological changes of the spiral arteries in the placental bed in relation to pre-eclampsia and fetal growth retardation. Br. J. Obstet. Gynaecol. 1981, 88, 876–881. [Google Scholar] [CrossRef]
- Albrecht, E.D.; Pepe, G.J. Regulation of Uterine Spiral Artery Remodeling: A Review. Reprod. Sci. 2020, 27, 1932–1942. [Google Scholar] [CrossRef]
- Lyall, F.; Robson, S.C.; Bulmer, J.N. Spiral artery remodeling and trophoblast invasion in preeclampsia and fetal growth restriction: Relationship to clinical outcome. Hypertension 2013, 62, 1046–1054. [Google Scholar] [CrossRef] [Green Version]
- Staff, A.C.; Fjeldstad, H.E.; Fosheim, I.K.; Moe, K.; Turowski, G.; Johnsen, G.M.; Alnaes-Katjavivi, P.; Sugulle, M. Failure of physiological transformation and spiral artery atherosis: Their roles in preeclampsia. Am. J. Obstet. Gynecol. 2020. [Google Scholar] [CrossRef]
- Brosens, I.; Puttemans, P.; Benagiano, G. Placental bed research: I. The placental bed: From spiral arteries remodeling to the great obstetrical syndromes. Am. J. Obstet. Gynecol. 2019, 221, 437–456. [Google Scholar] [CrossRef] [PubMed]
- Malamitsi-Puchner, A.; Economou, E.; Boutsikou, T.; Nikolaou, K.E.; Vrachnis, N. Neurotrophin-3 and FLT3 Tyrosine Kinase Receptor in Perinatal Life. Mediat. Inflamm. 2005, 2005, 53–56. [Google Scholar] [CrossRef]
- Kalampokas, E.; Vrachnis, N.; Samoli, E.; Rizos, D.; Iliodromiti, Z.; Sifakis, S.; Kalampokas, T.; Vitoratos, N.; Creatsas, G.; Botsis, D. Association of adiponectin and placental growth factor in amniotic fluid with second trimester fetal growth. In Vivo 2012, 26, 327–333. [Google Scholar] [PubMed]
- Vrachnis, N.; Gkogkas, L.; Iliodromiti, S.; Grigoriadis, C.; Samoli, E.; Pangalos, C.; Pappa, K.; Drakoulis, N.; Creatsas, G.; Botsis, D. Resistin in mid-trimester amniotic fluid in trisomy 21. J. Matern. Neonatal Med. 2013, 26, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Dalainas, I.; Papoutsis, D.; Samoli, E.; Rizos, D.; Iliodromiti, Z.; Siristatidis, C.; Tsikouras, P.; Creatsas, G.; Botsis, D. Soluble Fas and Fas-ligand levels in mid-trimester amniotic fluid and their associations with severe small for gestational age fetuses: A prospective observational study. J. Reprod. Immunol. 2013, 98, 39–44. [Google Scholar] [CrossRef]
- Vrachnis, N.; Malamitsi-Puchner, A.; Samoli, E.; Botsis, D.; Iliodromiti, Z.; Baka, S.; Hassiakos, D.; Creatsas, G. Elevated mid-trimester amniotic fluid DAM-8 concentrations as a potential risk factor for preterm delivery. J. Soc. Gynecol. Investig. 2006, 13, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Gilbert, W.M.; Sherman, M.P. Amniotic Fluid: Not Just Fetal Urine Anymore. J. Perinatol. 2005, 25, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lloyd, D.; Hale, C.; Stanislaus, S.; Chen, M.; Sivits, G.; Vonderfecht, S.; Hecht, R.; Li, Y.-S.; Lindberg, R.A.; et al. Fibroblast Growth Factor 21 Reverses Hepatic Steatosis, Increases Energy Expenditure, and Improves Insulin Sensitivity in Diet-Induced Obese Mice. Diabetes 2008, 58, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Hoo, R.L.-C.; Konishi, M.; Itoh, N.; Lee, P.-C.; Ye, H.-Y.; Lam, K.S.-L.; Xu, A. Growth Hormone Induces Hepatic Production of Fibroblast Growth Factor 21 through a Mechanism Dependent on Lipolysis in Adipocytes. J. Biol. Chem. 2011, 286, 34559–34566. [Google Scholar] [CrossRef] [Green Version]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mraz, M.; Bartlova, M.; Lacinová, Z.; Michalsky, D.; Kasalicky, M.; Haluzíková, D.; Matoulek, M.; Dostalova, I.; Humenanska, V.; Haluzik, M. Serum concentrations and tissue expression of a novel endocrine regulator fibroblast growth factor-21 in patients with type 2 diabetes and obesity. Clin. Endocrinol. 2009, 71, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xu, A.; Law, L.S.-C.; Chen, C.; Li, H.; Li, X.; Yang, L.; Liu, S.; Zhou, Z.; Lam, K.S.L. Distinct Changes in Serum Fibroblast Growth Factor 21 Levels in Different Subtypes of Diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E54–E58. [Google Scholar] [CrossRef] [Green Version]
- Vrachnis, N.; Iavazzo, C.; Iliodromiti, Z.; Sifakis, S.; Alexandrou, A.; Siristatidis, C.; Grigoriadis, C.; Botsis, D.; Creatsas, G. Diabetes mellitus and gynecologic cancer: Molecular mechanisms, epidemiological, clinical and prognostic perspectives. Arch. Gynecol. Obstet. 2016, 293, 239–246. [Google Scholar] [CrossRef]
- Mericq, V.; De Luca, F.; Hernandez, M.I.; Peña, V.; Rossel, K.; García, M.; Ávila, A.; Cavada, G.; Iñiguez, G. Serum Fibroblast Growth Factor 21 Levels Are Inversely Associated with Growth Rates in Infancy. Horm. Res. Paediatr. 2014, 82, 324–331. [Google Scholar] [CrossRef]
- Nicolini, U.; Hubinont, C.; Santolaya, J.; Fisk, N.M.; Coe, A.M.; Rodeck, C.H. Maternal-fetal glucose gradient in normal pregnancies and in pregnancies complicated by alloimmunization and fetal growth restriction. Am. J. Obstet. Gynecol. 1989, 161, 924–927. [Google Scholar] [CrossRef]
- Economides, D.L.; Nicolaides, K.H. Blood Glucose and Oxygen Tension Levels in Small-for-Gestational-Age Fetuses. Obstet. Anesth. Dig. 1989, 9, 143. [Google Scholar] [CrossRef]
- Ashworth, M.A.; Leach, F.N.; Milner, R.D. Development of insulin secretion in the human fetus. Arch. Dis. Child. 1973, 48, 151–152. [Google Scholar] [CrossRef] [Green Version]
- Nawathe, A.R.; Christian, M.; Kim, S.H.; Johnson, M.; Savvidou, M.D.; Terzidou, V. Insulin-like growth factor axis in pregnancies affected by fetal growth disorders. Clin. Epigenetics 2016, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Nardozza, L.M.M.; Caetano, A.C.R.; Zamarian, A.C.P.; Mazzola, J.B.; Silva, C.P.; Marçal, V.; Lobo, T.F.; Peixoto, A.B.; Júnior, E.A. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef]
- Bernstein, P.S.; Divon, M.Y. Etiologies of Fetal Growth Restriction. Clin. Obstet. Gynecol. 1997, 40, 723–729. [Google Scholar] [CrossRef]
- McCowan, L.; Horgan, R.P. Risk factors for small for gestational age infants. Best Pr. Res. Clin. Obstet. Gynaecol. 2009, 23, 779–793. [Google Scholar] [CrossRef]
- Murphy, V.E.; Smith, R.; Giles, W.B.; Clifton, V. Endocrine Regulation of Human Fetal Growth: The Role of the Mother, Placenta, and Fetus. Endocr. Rev. 2006, 27, 141–169. [Google Scholar] [CrossRef] [PubMed]
- Rehder, P.M.; Pereira, B.G.; Silva, J.L.P.E. The prognostic value of a normal oral glucose tolerance test in pregnant women who tested positive at screening: A validation study. Diabetol. Metab. Syndr. 2012, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausson, B.; Gardosi, J.; Francis, A.; Cnattingius, S. Perinatal outcome in SGA births defined by customised versus population-based birthweight standards. BJOG Int. J. Obstet. Gynaecol. 2001, 108, 830–834. [Google Scholar] [CrossRef]
- Beta, J.; Khan, N.; Khalil, A.; Fiolna, M.; Ramadan, G.; Akolekar, R. Maternal and neonatal complications of fetal macrosomia: Systematic review and meta-analysis. Ultrasound Obstet. Gynecol. 2019, 54, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Antonakopoulos, N.; Iliodromiti, Z.; Mastorakos, G.; Iavazzo, C.; Valsamakis, G.; Salakos, N.; Papageorghiou, A.; Margeli, A.; Kalantaridou, S.; Creatsas, G.; et al. Association between Brain-Derived Neurotrophic Factor (BDNF) Levels in 2nd Trimester Amniotic Fluid and Fetal Development. Mediat. Inflamm. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas-Ramos, D.; Aguilar-Salinas, C. Modulation of energy balance by fibroblast growth factor 21. Horm. Mol. Biol. Clin. Investig. 2016, 30. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Antonakopoulos, N.; Iliodromiti, Z.; Dafopoulos, K.; Siristatidis, C.; Pappa, K.I.; Deligeoroglou, E.; Vitoratos, N. Impact of Maternal Diabetes on Epigenetic Modifications Leading to Diseases in the Offspring. Exp. Diabetes Res. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Belitsos, P.; Sifakis, S.; Dafopoulos, K.; Siristatidis, C.; Pappa, K.I.; Iliodromiti, Z. Role of Adipokines and Other Inflammatory Mediators in Gestational Diabetes Mellitus and Previous Gestational Diabetes Mellitus. Int. J. Endocrinol. 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Wallner, W.; Sengenberger, R.; Strick, R.; Strissel, P.L.; Meurer, B.; Beckmann, M.W.; Schlembach, D. Angiogenic growth factors in maternal and fetal serum in pregnancies complicated by intrauterine growth restriction. Clin. Sci. 2006, 112, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Regnault, T.R.; Barker, P.L.; Botting, K.J.; McMillen, I.C.; McMillan, C.M.; Roberts, C.T.; Morrison, J.L. Placental Adaptations in Growth Restriction. Nutrients 2015, 7, 360–389. [Google Scholar] [CrossRef] [Green Version]
- Barut, F.; Barut, A.; Gun, B.D.; Kandemir, N.O.; Harma, M.I.; Harma, M.; Aktunc, E.; Ozdamar, S.O. Intrauterine growth restriction and placental angiogenesis. Diagn. Pathol. 2010, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Gardosi, J.; Francis, A.; Turner, S.; Williams, M. Customized growth charts: Rationale, validation and clinical benefits. Am. J. Obstet. Gynecol. 2018, 218, S609–S618. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variable | SGA (n = 31) | LGA (n = 18) | AGA (n = 31) | p-Value |
|---|---|---|---|---|
| Maternal age (years) | 35 (32–37) | 35 (32–37) | 37 (36–38) | 0.01 |
| Maternal weight (kg) | 61.5 (56.5–72) | 60.5 (55–64) | 66 (60–74) | 0.15 |
| Maternal height (cm) | 167 (165–171) | 166 (159–170) | 168 (163.5–170) | 0.60 |
| Maternal parity | 0 (0–1) | 1 (0–1) | 1 (0–1.5) | 0.24 |
| Maternal smoking | 23 (76.7%) | 17 (94.4%) | 29 (93.6%) | 0.13 |
| Fetal sex female | 19 (63.3%) | 4 (23.5%) | 11 (36.7%) | 0.02 |
| Birth week | 38 (38–39) | 38 (37–39) | 38 (37–39) | 0.11 |
| Birth weight (gr) | 3300 (3200–3510) | 3870 (3690–4180) | 2580 (2440–2750) | 0.01 |
| Subgroup | n of Cases | Median Value (Q1–Q3) |
|---|---|---|
| SGA < 10th centile | 31 | 4.906 (3.395–9.186) |
| SGA < 5th centile | 18 | 4.898 (3.247–7.555) |
| SGA < 3rd centile | 10 | 4.069 (2.524–6.769) |
| AGA | 31 | 4.636 (1.891–6.494) |
| LGA > 90th centile | 18 | 6.337 (3.088–6.852) |
| LGA > 95th centile | 05 | 6.337 (3.631–8.508) |
| Subgroup | n of Cases | Median Value (Q1–Q3) |
|---|---|---|
| SGA < 10th centile | 24 | 2.27 (2–3.355) |
| SGA < 5th centile | 14 | 2.34 (2–4.03) |
| SGA < 3rd centile | 08 | 3.46 (2.155–6.885) |
| AGA | 27 | 2.40 (2–2.88) |
| LGA > 90th centile | 17 | 2.24 (2–3.74) |
| LGA > 95th centile | 04 | 2.44 (2.16–3.25) |
| SGA and AGA | LGA and AGA | |||
|---|---|---|---|---|
| Parameter | OR (95%CI) | p | OR (95%CI) | p |
| Maternal age | 1.31 (1.05–1.65) | 0.0179 | 0.95 (0.77–1.18) | 0.64 |
| Maternal weight | 1.05 (0.99–1.1) | 0.1147 | 0.95 (0.87–1.03) | 0.2221 |
| Maternal height | 1.07 (0.93–1.21) | 0.3453 | 0.98 (0.86–1.11) | 0.7284 |
| Fetal sex (ref: male) | 4.38 (1.06–18.12) | 0.0416 | 0.36 (0.06–2.12) | 0.2568 |
| Maternal smoking (ref: no) | 8.99 (1.04–77.86) | 0.0461 | 0.74 (0.04–12.97) | 0.8335 |
| FGF 21 | 1.07 (1.01–1.21) | 0.0303 | 1.03 (0.89–1.2) | 0.0406 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrachnis, N.; Argyridis, S.; Vrachnis, D.; Antonakopoulos, N.; Valsamakis, G.; Iavazzo, C.; Zygouris, D.; Salakos, N.; Rodolakis, A.; Vlahos, N.; et al. Increased Fibroblast Growth Factor 21 (FGF21) Concentration in Early Second Trimester Amniotic Fluid and Its Association with Fetal Growth. Metabolites 2021, 11, 581. https://doi.org/10.3390/metabo11090581
Vrachnis N, Argyridis S, Vrachnis D, Antonakopoulos N, Valsamakis G, Iavazzo C, Zygouris D, Salakos N, Rodolakis A, Vlahos N, et al. Increased Fibroblast Growth Factor 21 (FGF21) Concentration in Early Second Trimester Amniotic Fluid and Its Association with Fetal Growth. Metabolites. 2021; 11(9):581. https://doi.org/10.3390/metabo11090581
Chicago/Turabian StyleVrachnis, Nikolaos, Savvas Argyridis, Dionysios Vrachnis, Nikolaos Antonakopoulos, Georgios Valsamakis, Christos Iavazzo, Dimitrios Zygouris, Nikolaos Salakos, Alexandros Rodolakis, Nikolaos Vlahos, and et al. 2021. "Increased Fibroblast Growth Factor 21 (FGF21) Concentration in Early Second Trimester Amniotic Fluid and Its Association with Fetal Growth" Metabolites 11, no. 9: 581. https://doi.org/10.3390/metabo11090581
APA StyleVrachnis, N., Argyridis, S., Vrachnis, D., Antonakopoulos, N., Valsamakis, G., Iavazzo, C., Zygouris, D., Salakos, N., Rodolakis, A., Vlahos, N., Mastorakos, G., Drakakis, P., & Iliodromiti, Z. (2021). Increased Fibroblast Growth Factor 21 (FGF21) Concentration in Early Second Trimester Amniotic Fluid and Its Association with Fetal Growth. Metabolites, 11(9), 581. https://doi.org/10.3390/metabo11090581