
A Targeted Serum Metabolomics GC-MS Approach Identifies Predictive Blood Biomarkers for Retained Placenta in Holstein Dairy Cows

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
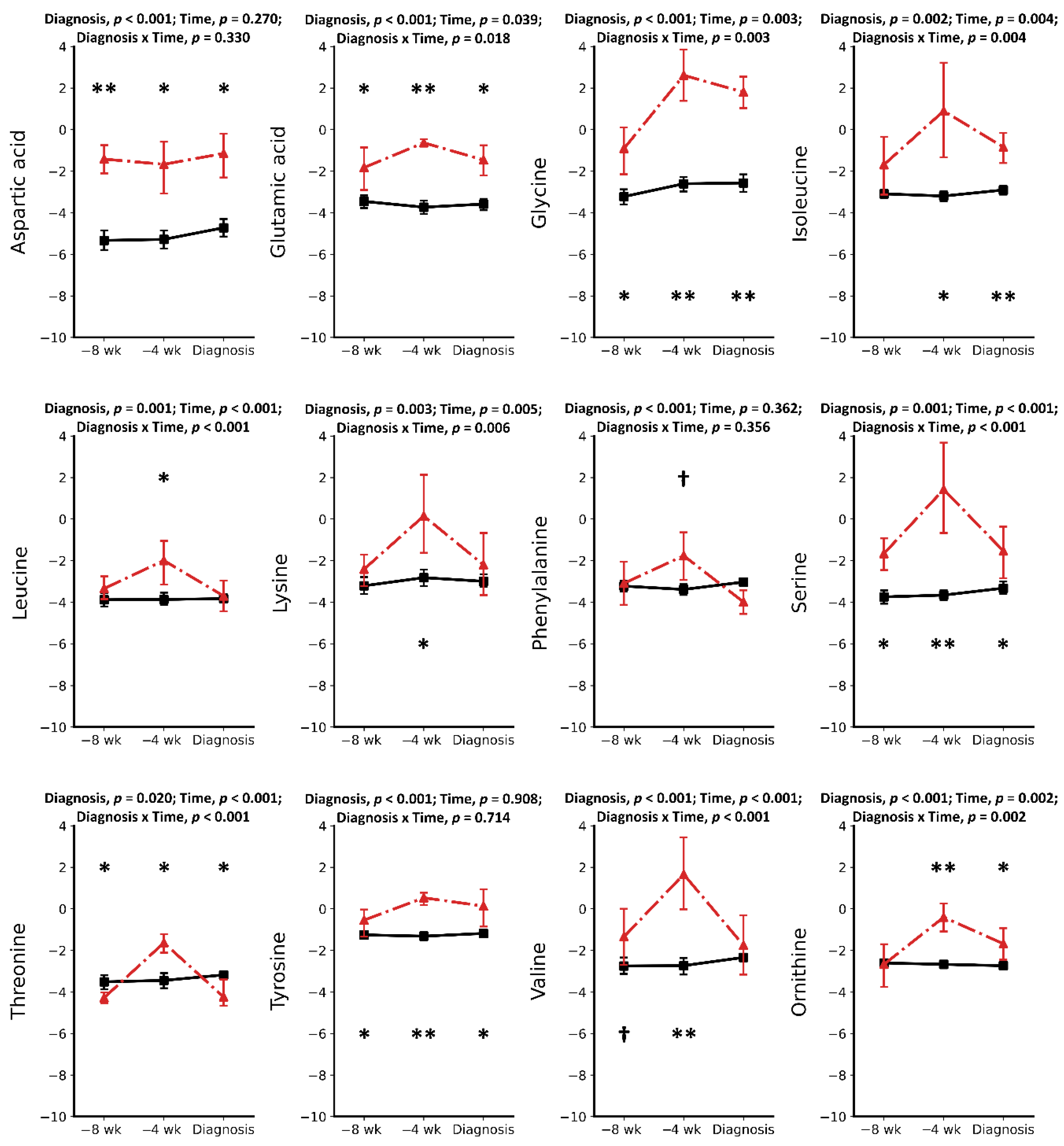
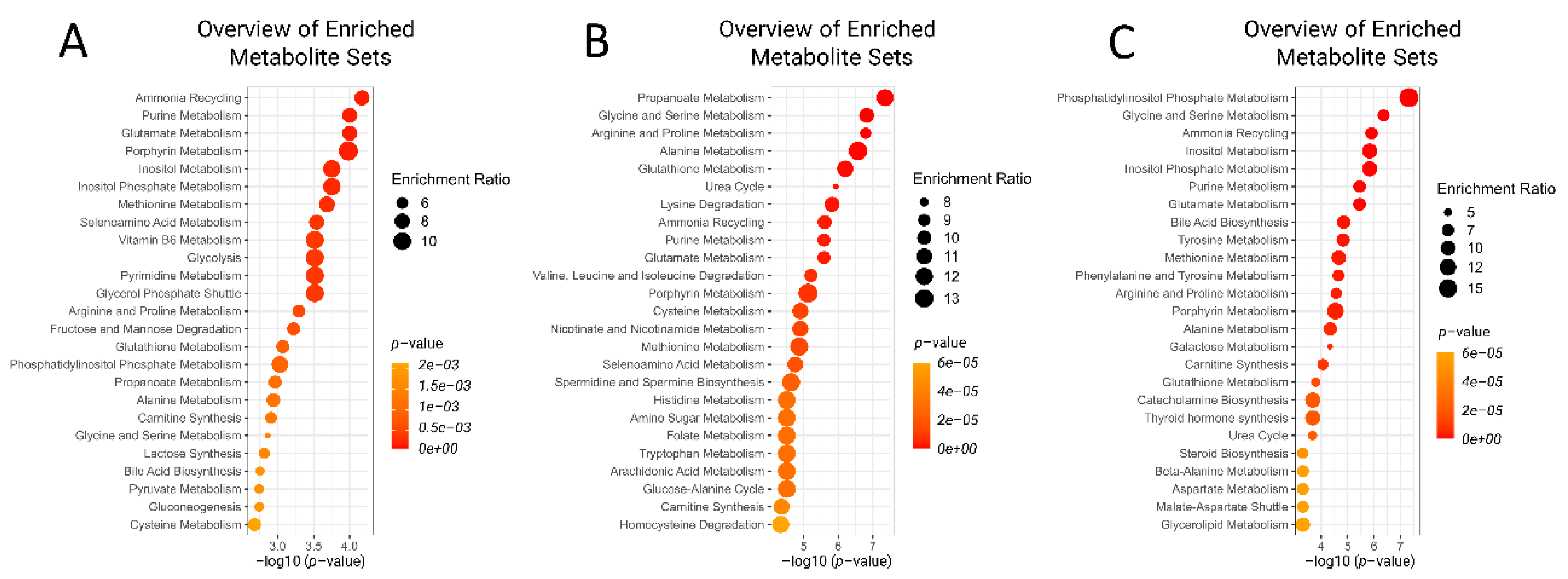
2.1. Metabolic Alterations Prior to Diagnosis of Retained Placenta
2.2. Metabolic Alterations during the Week of Diagnosis of Retained Placenta
3. Discussion
4. Materials and Methods
4.1. Animals and Diets
4.2. Blood Sample Collection
4.3. GC-MS Compound Identification and Quantification
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelton, D.F.; Lissemore, K.D.; Martin, R.E. Recommendations for recording and calculating the incidence of selected clinical diseases of dairy cattle. J. Dairy Sci. 1998, 81, 2502–2509. [Google Scholar] [CrossRef]
- Beagley, J.; Whitman, K.; Baptiste, K.; Scherzer, J. Physiology and Treatment of Retained Fetal Membranes in Cattle. J. Vet. Intern. Med. 2010, 24, 261–268. [Google Scholar] [CrossRef]
- Joosten, I.; Stelwagen, J.; Dijkhuizen, A.A. Economic and reproductive consequences of retained placenta in dairy cattle. Vet. Rec. 1988, 123, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.K.; Kim, I.H. Risk factors for retained placenta and the effect of retained placenta on the occurrence of postpartum diseases and subsequent reproductive performance in dairy cows. J. Vet. Sci. 2005, 6, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Occurrence of retained placenta is preceded by an inflammatory state and alterations of energy metabolism in transition dairy cows. J. Anim. Sci. Biotechno. 2016, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dervishi, E.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Targeted metabolomics: New insights into pathobiology of retained placenta in dairy cows and potential risk biomarkers. Animal 2018, 12, 1050–1059. [Google Scholar] [CrossRef]
- Martin, F.P.J.; Collino, S.; Rezzi, S.; Kochhar, S. Metabolomic applications to decipher gut microbial metabolic influence in health and disease. Front. Physilo. 2012, 3, 113. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailemariam, D.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Identification of serum metabolites associated with the risk of metritis in transition dairy cows. Can. J. Ani. Sci. 2017, 98, 525–537. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC-MS metabolomics identifies metabolite alterations that precede subclinical mastitis in the blood of transition dairy cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Serum metabolic fingerprinting of pre-lameness dairy cows by GC-MS reveals typical profiles that can identify susceptible cows. J. Proteomics 2020, 213, 103620. [Google Scholar] [CrossRef]
- Hailemariam, D.; Mandal, R.; Saleem, F.; Dunn, S.M.; Wishart, D.S.; Ametaj, B.N. Identification of predictive biomarkers of disease state in transition dairy cows. J. Dairy Sci. 2014, 97, 2680–2693. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, C.; Xia, C.; Zhang, H.; Sun, L.; Gao, Y. Plasma metabolic profiling of dairy cows affected with clinical ketosis using LC/MS technology. Vet. Q. 2014, 34, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wu, L.; Xu, C.; Xia, C.; Sun, L.; Shu, S. Plasma metabolomic profiling of dairy cows affected with ketosis using gas chromatography / mass spectrometry. BMC Vet. Res. 2013, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. A multi-platform metabolomics approach identifies urinary metabolite signatures that differentiate ketotic from healthy dairy cows. Front. Vet. Sci. 2021, 8, 595983. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, C. What’s next in ’omics: The metabolome. Science 2014, 345, 1519–1521. [Google Scholar] [CrossRef] [Green Version]
- Dervishi, E.; Ametaj, B.N. Retained Placenta: A Systems Veterinary Approach. In Periparturient Diseases of Dairy Cows; Ametaj, B., Ed.; Springer: Cham, Switzerland, 2017; pp. 121–138. [Google Scholar] [CrossRef]
- Ganeshan, K.; Chawla, A. Metabolic regulation of immune responses. Annu. Rev. Immunol. 2014, 32, 609–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Luan, H.H.; Medzhitov, R. An evolutionary perspective on immunometabolism. Science 2019, 363, eaar3932. [Google Scholar] [CrossRef] [PubMed]
- Villa, E.; Ali, E.S.; Sahu, U.; Ben-Sahra, I. Cancer cells tune the signaling pathways to empower de Novo synthesis of nucleotides. Cancers 2019, 11, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pareek, V.; Tian, H.; Winograd, N.; Benkovic, S.J. Metabolomics and mass spectrometry imaging reveal channeled de novo purine synthesis in cells. Science 2020, 368, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.; Pearce, E.L. Amino Assets: How Amino Acids Support Immunity. Cell Metab. 2020, 32, 154–175. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Shi, H.; Wang, X.; Zhu, H.; Pi, D.; Leng, W.; Li, S. Aspartate attenuates intestinal injury and inhibits TLR4 and NODs/NF-κB and p38 signaling in weaned pigs after LPS challenge. Eur. J. Nutr. 2016, 56, 1433–1443. [Google Scholar] [CrossRef]
- Bin, P.; Liu, S.; Chen, S.; Zeng, Z.; Huang, R.; Yin, Y.; Liu, G. The effect of aspartate supplementation on the microbial composition and innate immunity on mice. Amino Acids 2017, 49, 2045–2051. [Google Scholar] [CrossRef]
- Ganor, Y.; Levite, M. The neurotransmitter glutamate and human T cells: Glutamate receptors and glutamate-induced direct and potent effects on normal human T cells, cancerous human leukemia and lymphoma T cells, and autoimmune human T cells. J. Neural. Transm. 2014, 121, 983–1006. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lin, Y.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, R.; Gallart, T.; Lluis, C.; Franco, R. Role of glutamate on T-cell mediated immunity. J. Neuroimmunol. 2007, 185, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. Glutamate: A truly functional amino acid. Amino Acids 2013, 45, 413–418. [Google Scholar] [CrossRef]
- Shanshiashvili, L.; Tsitsilashvili, E.; Dabrundashvili, N.; Kalandadze, I.; Mikeladze, D. Metabotropic glutamate receptor 5 may be involved in macrophage plasticity. Biol. Res. 2017, 50, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Koning, T.J.; Fuchs, S.A.; Klomp, L.W.J. Serine, Glycine, and Threonine. In Handbook of Neurochemistry and Molecular Neurobiology; Lajtha, A., Oja, S.S., Schousboe, A., Saransaari, P., Eds.; Springer: Boston, MA, USA, 2007; pp. 23–45. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Tenenhouse, H.S.; Deutsch, H.F. Some physical–chemical properties of chicken gamma-globulins and their pepsin and papain digestion product. Immunochemistry 1966, 3, 11–20. [Google Scholar] [CrossRef]
- Wang, X.; Qiao, S.Y.; Liu, M.; Ma, Y.X. Effects of graded levels of true ileal digestible threonine on performance, serum parameters and immune function of 10–25 kg pigs. Anim. Feed Sci. Tech. 2006, 129, 264–278. [Google Scholar] [CrossRef]
- Mawal-Dewan, M.; Lorenzini, A.; Frisoni, L.; Zhang, H.; Cristofalo, V.J.; Sell, C. Regulation of collagenase expression during replicative senescence in human fibroblasts by Akt-forkhead signaling. J. Biol. Chem. 2002, 277, 7857–7864. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.E.; Ducker, G.S.; Billingham, L.K.; Martinez, C.A.; Mainolfi, N.; Suri, V.; Friedman, A.; Manfredi, M.G.; Weinberg, S.E.; Rabinowitz, J.D.; et al. Serine Metabolism Supports Macrophage IL-1β Production. Cell Metab. 2019, 29, 1003–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, A.; Yokote, Y.; Sakagami, H. Changes in amino acid metabolism during activation of mouse macrophage-like cell lines. In Vivo 2010, 24, 857–860. [Google Scholar] [PubMed]
- Xu, F.L.; You, H.B.; Li, X.H.; Chen, X.F.; Liu, Z.J.; Gong, J.P. Glycine attenuates endotoxin-induced liver injury by downregulating TLR4 signaling in Kupffer cells. Am. J. Surg. 2008, 196, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Zangerle, R.; Kurz, K.; Neurauter, G.; Kitchen, M.; Sarcletti, M.; Fuchs, D. Increased blood phenylalanine to tyrosine ratio in HIV-1 infection and correction following effective antiretroviral therapy. Brain Behav. Immun. 2010, 24, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Sikalidis, A.K. Amino Acids and Immune Response: A Role for Cysteine, Glutamine, Phenylalanine, Tryptophan and Arginine in T-cell Function and Cancer? Pathol. Oncol. Res. 2015, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Murr, C.; Grammar, T.B.; Meinitzer, A.; Kleber, M.E.; Marz, W.; Fuchs, D. Immune activation and inflammation in patients with cardiovascular disease are associated with higher phenylalanine to tyrosine ratios: The ludwigshafen risk and cardiovascular health study. J. Amino Acids 2014, 2014, 783730. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Mateo, R.D.; Yin, Y.L.; Wu, G. Functional amino acids and fatty acids for enhancing production performance of sows and piglets. Asian-Aust. J. Anim. Sci. 2007, 20, 295–306. [Google Scholar] [CrossRef]
- Kin, N.W.; Sanders, V.M. It takes nerve to tell T and B cells what to do. J. Leukoc. Biol. 2006, 79, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Bredahl, E.C.; Eckerson, J.M.; Tracy, S.M.; McDonald, T.L.; Drescher, K.M. The Role of Creatine in the Development and Activation ofImmune Responses. Nutrients 2021, 13, 751. [Google Scholar] [CrossRef] [PubMed]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Riesberg, L.A.; McDonald, T.L.; Wang, Y.; Chen, X.M.; Holzmer, S.W.; Tracy, S.M.; Drescher, K.M. Creatinine downregulates TNF-α in macrophage and T cell lines. Cytokine 2018, 110, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Chibisa, G.E.; Gozho, G.N.; Van Kessel, A.G.; Olkowski, A.A.; Mutsvangwa, T. Effects of peripartum propylene glycol supplementation on nitrogen metabolism, body composition, and gene expression for the major protein degradation pathways in skeletal muscle in dairy cows. J. Dairy Sci. 2008, 91, 3512–3527. [Google Scholar] [CrossRef] [Green Version]
- van der Drift, S.G.A.; Houweling, M.; Schonewille, J.T.; Tielens, A.G.M.; Jorritsma, R. Protein and fat mobilization and associations with serum β-hydroxybutyrate concentrations in dairy cows. J. Dairy Sci. 2012, 95, 4911–4920. [Google Scholar] [CrossRef]
- Li, J.P.; Lu, L.; Wang, L.J.; Zhang, F.R.; Shen, W.F. Increased serum levels of S100B are related to the severity of cardiac dysfunction, renal insufficiency and major cardiac events in patients with chronic heart failure. Clin. Biochem. 2011, 44, 984–988. [Google Scholar] [CrossRef] [PubMed]
- Niewczas, M.A.; Gohda, T.; Skupien, J.; Smiles, A.M.; Walker, W.H.; Rosetti, F.; Cullere, X.; Eckfeldt, J.H.; Doria, A.; Mayadas, T.N.; et al. Circulating TNF receptors 1 and 2 predict ESRD in type 2 diabetes. J. Am. Soc. Nephrol. 2012, 23, 507–515. [Google Scholar] [CrossRef]
- Seruga, B.; Zhang, H.; Bernstein, L.J.; Tannock, I.F. Cytokines and their relationship to the symptoms and outcome of cancer. Nat. Rev. Cancer. 2008, 8, 887–899. [Google Scholar] [CrossRef]
- Walston, J.; Fedarko, N.; Yang, H.; Leng, S.; Beamer, B.; Espinoza, S.; Lipton, A.; Zheng, H.; Becker, K. The physical and biological characterization of a frail mouse model. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Holub, B.J. The Nutritional Significance, Metabolism, and Function of Myo-Inositol and Phosphatidylinositol in Health and Disease. Adv. Nutr. Res. 1982, 4, 107–141. [Google Scholar] [CrossRef]
- Chhetri, D.R. Myo-Inositol and Its Derivatives: Their Emerging Role in the Treatment of Human Diseases. Front. Pharmacol. 2019, 10, 1172. [Google Scholar] [CrossRef] [Green Version]
- Corrado, F.; Santamaria, A. Chapter 19—Myoinositol Supplementation on Insulin Resistance in Gestational Diabetes. In Glucose Intake and Utilization in Pre-Diabetes and Diabetes; Watson, R.R., Dokken, B.B., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 229–234. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, B.; Li, H.; Peng, X. Myo-inositol improves the host’s ability to eliminate balofloxacin-resistant Escherichia coli. Sci. Report. 2015, 5, 10720. [Google Scholar] [CrossRef] [Green Version]
- Unver, N.; Delgado, O.; Zeleke, K.; Cumpian, A.; Tang, X.; Caetano, M.S.; Wang, H.; Katayama, H.; Yu, H.; Szabo, E.; et al. Reduced IL-6 levels and tumor-associated phospho-STAT3 are associated with reduced tumor development in a mouse model of lung cancer chemoprevention with myo-inositol. Int. J. Cancer. 2018, 142, 1405–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laganà, A.S.; Unfer, V.; Garzon, S.; Bizzarri, M. Role of inositol to improve surfactant functions and reduce IL-6 levels: A potential adjuvant strategy for SARS-CoV-2 pneumonia? Med. Hypotheses 2020, 144, 110262. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir Med. 2020, 4, 420–422. [Google Scholar] [CrossRef]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (SLC25A1) in TNFα- and IFNγ-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Gu, C.; Ren, M.; Chen, D.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, J.; Luo, Y.; et al. L-Isoleucine Administration Alleviates Rotavirus Infection and Immune Response in the Weaned Piglet Model. Front. Immunol. 2018, 16, 1654. [Google Scholar] [CrossRef] [Green Version]
- Olfert, E.D.; Cross, B.M.; McWilliam, A.A. Guide to the Care and use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993; Volume 1. [Google Scholar]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Deng, Q.; Goldansaz, S.A.; Dunn, S.M.; Ametaj, B. Alterations of innate immunity reactants in transition dairy cows before clinical signs of lameness. Animals 2015, 5, 717–747. [Google Scholar] [CrossRef]
- Zhang, G.; Dervishi, E.; Dunn, S.M.; Mandal, R.; Liu, P.; Han, B.; Wishart, D.S.; Ametaj, B.N. Metabotyping reveals distinct metabolic alterations in ketotic cows and identifies early predictive serum biomarkers for the risk of disease. Metabolomics 2017, 13, 43. [Google Scholar] [CrossRef]
- Wishart, D.S.; Lewis, M.J.; Morrissey, J.A.; Flegel, M.D.; Jeroncic, K.; Xiong, Y.; Dean Cheng, D.; Eisner, R.; Gautam, B.; Tzur, D.; et al. The human cerebrospinal fluid metabolome. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2008, 871, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite. mM | 8 Weeks before Parturition | |||||
|---|---|---|---|---|---|---|
| Pre-RP | CON | p-Value | Log (BF₁₀) | Fold Change | RP/CON | |
| Number of cases | 6 | 16 | - | - | - | |
| Aspartic acid | 0.45 (0.12) | 0.03 (0.01) | <0.001 | 7.07 | 13.35 | Up |
| Cholesterol | 0.66 (0.12) | 0.28 (0.06) | 0.01 | 2.05 | 2.35 | Up |
| Citric acid | 0.11 (0.04) | 0.07 (0.01) | 0.35 | −0.20 | 1.50 | Up |
| Creatinine | 0.35 (0.07) | 0.30 (0.02) | 0.30 | −0.57 | 1.19 | Up |
| D-Mannose | 0.07 (0.02) | 0.03 (0.00) | 0.19 | 2.45 | 2.74 | Up |
| Galactose | 3.03 (0.83) | 3.63 (0.38) | 0.54 | −0.68 | 0.83 | Down |
| Glutamic acid | 0.43 (0.15) | 0.1 (0.01) | 0.02 | 3.11 | 4.25 | Up |
| Glycine | 0.80 (0.20) | 0.12 (0.02) | 0.01 | 6.74 | 6.45 | Up |
| Isoleucine | 0.68 (0.32) | 0.12 (0.01) | 0.17 | 1.73 | 5.49 | Up |
| Lactose | 0.12 (0.06) | 0.17 (0.04) | 0.18 | −0.74 | 0.70 | Down |
| Leucine | 0.12 (0.03) | 0.08 (0.02) | 0.14 | −0.66 | 1.37 | Up |
| L-Lactic acid | 0.80 (0.41) | 0.66 (0.15) | 0.79 | −0.82 | 1.22 | Up |
| Lysine | 0.25 (0.08) | 0.14 (0.03) | 0.10 | 0.12 | 1.84 | Up |
| Myo-Inositol | 0.04 (0.01) | 0.01 (0.00) | 0.01 | 7.63 | 6.17 | Up |
| Oleic acid | 1.27 (0.44) | 0.19 (0.04) | 0.02 | 3.86 | 6.86 | Up |
| Ornithine | 0.25 (0.08) | 0.17 (0.01) | 0.43 | −0.24 | 1.43 | Up |
| Oxalate | 1.71 (0.75) | 1.58 (0.41) | 0.84 | −0.87 | 1.08 | Up |
| Palmitic acid | 0.21 (0.08) | 0.07 (0.01) | 0.25 | 1.52 | 2.85 | Up |
| Phenylalanine | 0.19 (0.07) | 0.12 (0.01) | 0.85 | 0.08 | 1.68 | Up |
| Phosphoric acid | 1.31 (0.39) | 0.35 (0.08) | 0.02 | 3.04 | 3.75 | Up |
| Pyroglutamic acid | 0.13 (0.04) | 0.06 (0.01) | 0.13 | 1.03 | 2.15 | Up |
| Serine | 0.41 (0.13) | 0.09 (0.01) | 0.003 | 3.68 | 4.76 | Up |
| Stearic acid | 0.63 (0.28) | 0.21 (0.05) | 0.25 | 0.85 | 3.03 | Up |
| Threonine | 0.05 (0.01) | 0.10 (0.01) | 0.01 | 1.08 | 0.54 | Down |
| Tyrosine | 0.80 (0.14) | 0.43 (0.03) | 0.02 | 3.56 | 1.85 | Up |
| Urea | 5.79 (1.46) | 1.78 (0.24) | 0.01 | 4.08 | 3.26 | Up |
| Valine | 0.91 (0.46) | 0.18 (0.03) | 0.09 | 1.29 | 5.06 | Up |
| Metabolite. mM | −4 Weeks before Parturition | |||||
|---|---|---|---|---|---|---|
| Pre-RP | CON | p-Value | Log (BF₁₀) | Fold Change | RP/CON | |
| Number of cases | 6 | 16 | - | - | - | |
| Aspartic acid | 0.53 (0.15) | 0.03 (0.01) | 0.003 | 6.65 | 15.97 | Up |
| Cholesterol | 0.70 (0.20) | 0.29 (0.05) | 0.10 | 1.85 | 2.39 | Up |
| Citric acid | 0.33 (0.07) | 0.08 (0.01) | 0.01 | 6.30 | 4.34 | Up |
| Creatinine | 0.87 (0.20) | 0.32 (0.02) | 0.02 | 4.65 | 2.68 | Up |
| D-Mannose | 0.10 (0.03) | 0.04 (0.01) | 0.31 | 0.45 | 2.46 | Up |
| Galactose | 4.33 (1.02) | 3.56 (0.37) | 0.35 | −0.60 | 1.22 | Up |
| Glutamic acid | 0.65 (0.05) | 0.09 (0.01) | <0.001 | 22.98 | 7.69 | Up |
| Glycine | 12.24 (5.31) | 0.19 (0.03) | <0.001 | 3.39 | 64.09 | Up |
| Isoleucine | 10.43 (5.20) | 0.12 (0.01) | 0.02 | 2.51 | 88.36 | Up |
| Lactose | 0.07 (0.02) | 0.13 (0.03) | 0.25 | −0.26 | 0.52 | Down |
| Leucine | 0.36 (0.10) | 0.08 (0.02) | 0.03 | 4.23 | 4.39 | Up |
| L-Lactic acid | 0.21 (0.08) | 1.03 (0.39) | 0.22 | −0.34 | 0.21 | Down |
| Lysine | 3.95 (2.02) | 0.19 (0.05) | 0.01 | 2.14 | 20.78 | Up |
| Myo-Inositol | 0.09 (0.03) | 0.01 (0.00) | 0.01 | 6.82 | 13.43 | Up |
| Oleic acid | 0.39 (0.10) | 0.34 (0.11) | 0.23 | −0.86 | 1.14 | Up |
| Ornithine | 0.96 (0.28) | 0.16 (0.01) | <0.001 | 5.38 | 6.03 | Up |
| Oxalate | 2.79 (0.67) | 2.03 (0.43) | 0.24 | −0.59 | 1.37 | Up |
| Palmitic acid | 0.17 (0.04) | 0.11 (0.02) | 0.16 | −0.08 | 1.62 | Up |
| Phenylalanine | 0.46 (0.13) | 0.10 (0.01) | 0.05 | 4.91 | 4.42 | Up |
| Phosphoric acid | 1.58 (0.42) | 0.33 (0.05) | 0.01 | 5.32 | 4.85 | Up |
| Pyroglutamic acid | 0.41 (0.03) | 0.06 (0.01) | <0.001 | 19.12 | 6.53 | Up |
| Serine | 12.87 (6.72) | 0.09 (0.01) | <0.001 | 2.27 | 151.35 | Up |
| Stearic acid | 0.64 (0.20) | 0.28 (0.06) | 0.14 | 0.87 | 2.30 | Up |
| Threonine | 0.36 (0.07) | 0.11 (0.01) | 0.001 | 6.94 | 3.35 | Up |
| Tyrosine | 1.50 (0.16) | 0.42 (0.03) | <0.001 | 14.31 | 3.55 | Up |
| Urea | 4.32 (0.83) | 2.05 (0.28) | 0.01 | 2.49 | 2.11 | Up |
| Valine | 10.44 (5.27) | 0.18 (0.02) | <0.001 | 2.40 | 59.3 | Up |
| Metabolite. mM | RP Diagnosis Week | |||||
|---|---|---|---|---|---|---|
| RP | CON | p-Value | Log (BF₁₀) | Fold Change | RP/CON | |
| Number of cases | 6 | 16 | - | - | - | |
| Aspartic acid | 0.73 (0.29) | 0.05 (0.01) | 0.002 | 3.64 | 15.6 | Up |
| Cholesterol | 0.63 (0.13) | 0.34 (0.05) | 0.08 | 1.24 | 1.88 | Up |
| Citric acid | 0.21 (0.05) | 0.08 (0.01) | 0.01 | 3.72 | 2.78 | Up |
| Creatinine | 1.08 (0.25) | 0.33 (0.02) | <0.001 | 5.50 | 3.26 | Up |
| D-Mannose | 0.05 (0.03) | 0.03 (0.00) | 0.39 | −0.42 | 1.67 | Up |
| Galactose | 4.25 (0.58) | 4.64 (0.26) | 0.85 | −0.70 | 0.91 | Down |
| Glutamic acid | 0.45 (0.12) | 0.09 (0.01) | 0.001 | 5.46 | 5.01 | Up |
| Glycine | 4.41 (1.10) | 0.21 (0.04) | <0.001 | 8.34 | 20.72 | Up |
| Isoleucine | 0.71 (0.21) | 0.14 (0.01) | <0.001 | 4.62 | 5.05 | Up |
| Lactose | 0.20 (0.10) | 0.12 (0.03) | 0.58 | −0.48 | 1.66 | Up |
| Leucine | 0.10 (0.04) | 0.07 (0.01) | 0.31 | −0.42 | 1.41 | Up |
| L-Lactic acid | 0.49 (0.22) | 0.74 (0.18) | 0.60 | −0.68 | 0.67 | Down |
| Lysine | 0.61 (0.37) | 0.15 (0.04) | 0.48 | 0.46 | 4.04 | Up |
| Myo-Inositol | 0.09 (0.02) | 0.01 (0.00) | <0.001 | 9.90 | 17.60 | Up |
| Oleic acid | 2.64 (0.77) | 0.56 (0.15) | 0.003 | 3.68 | 4.70 | Up |
| Ornithine | 0.42 (0.15) | 0.15 (0.01) | 0.02 | 1.92 | 2.71 | Up |
| Oxalate | 1.99 (0.94) | 2.73 (0.46) | 0.20 | −0.67 | 0.73 | Down |
| Palmitic acid | 0.63 (0.20) | 0.16 (0.03) | 0.004 | 2.99 | 4.02 | Up |
| Phenylalanine | 0.07 (0.02) | 0.13 (0.01) | 0.01 | 1.94 | 0.59 | Down |
| Phosphoric acid | 1.10 (0.38) | 0.33 (0.05) | 0.04 | 2.29 | 3.36 | Up |
| Pyroglutamic acid | 0.13 (0.03) | 0.06 (0.01) | 0.06 | 2.14 | 2.15 | Up |
| Serine | 0.59 (0.19) | 0.12 (0.02) | 0.02 | 3.42 | 4.97 | Up |
| Stearic acid | 1.83 (0.69) | 0.57 (0.14) | 0.02 | 1.40 | 3.19 | Up |
| Threonine | 0.07 (0.03) | 0.11 (0.01) | 0.02 | 0.36 | 0.63 | Down |
| Tyrosine | 1.63 (0.61) | 0.45 (0.03) | 0.01 | 2.31 | 3.61 | Up |
| Urea | 10.65 (3.47) | 2.28 (0.24) | 0.006 | 3.69 | 4.67 | Up |
| Valine | 0.70 (0.33) | 0.20 (0.01) | 0.31 | 1.20 | 3.43 | Up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Tobolski, D.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. A Targeted Serum Metabolomics GC-MS Approach Identifies Predictive Blood Biomarkers for Retained Placenta in Holstein Dairy Cows. Metabolites 2021, 11, 633. https://doi.org/10.3390/metabo11090633
Zhang G, Tobolski D, Zwierzchowski G, Mandal R, Wishart DS, Ametaj BN. A Targeted Serum Metabolomics GC-MS Approach Identifies Predictive Blood Biomarkers for Retained Placenta in Holstein Dairy Cows. Metabolites. 2021; 11(9):633. https://doi.org/10.3390/metabo11090633
Chicago/Turabian StyleZhang, Guanshi, Dawid Tobolski, Grzegorz Zwierzchowski, Rupasri Mandal, David S. Wishart, and Burim N. Ametaj. 2021. "A Targeted Serum Metabolomics GC-MS Approach Identifies Predictive Blood Biomarkers for Retained Placenta in Holstein Dairy Cows" Metabolites 11, no. 9: 633. https://doi.org/10.3390/metabo11090633
APA StyleZhang, G., Tobolski, D., Zwierzchowski, G., Mandal, R., Wishart, D. S., & Ametaj, B. N. (2021). A Targeted Serum Metabolomics GC-MS Approach Identifies Predictive Blood Biomarkers for Retained Placenta in Holstein Dairy Cows. Metabolites, 11(9), 633. https://doi.org/10.3390/metabo11090633