



Response of Intestinal Microbiota to the Variation in Diets in Grass Carp (Ctenopharyngodon idella)

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Illumina Sequencing of Bacterial 16S rRNA Gene
2.3. Bioinformatics and Statistical Analysis
3. Results
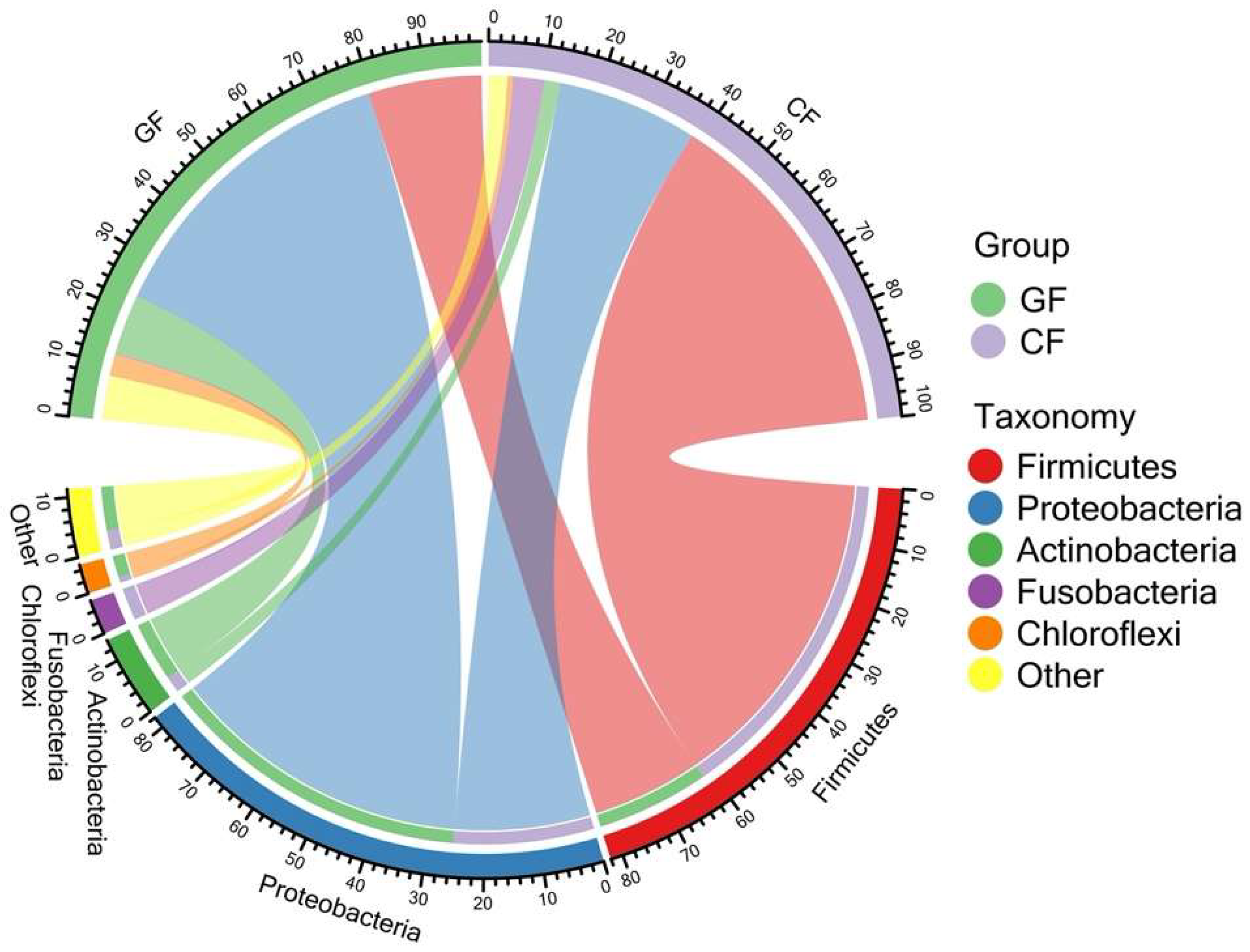
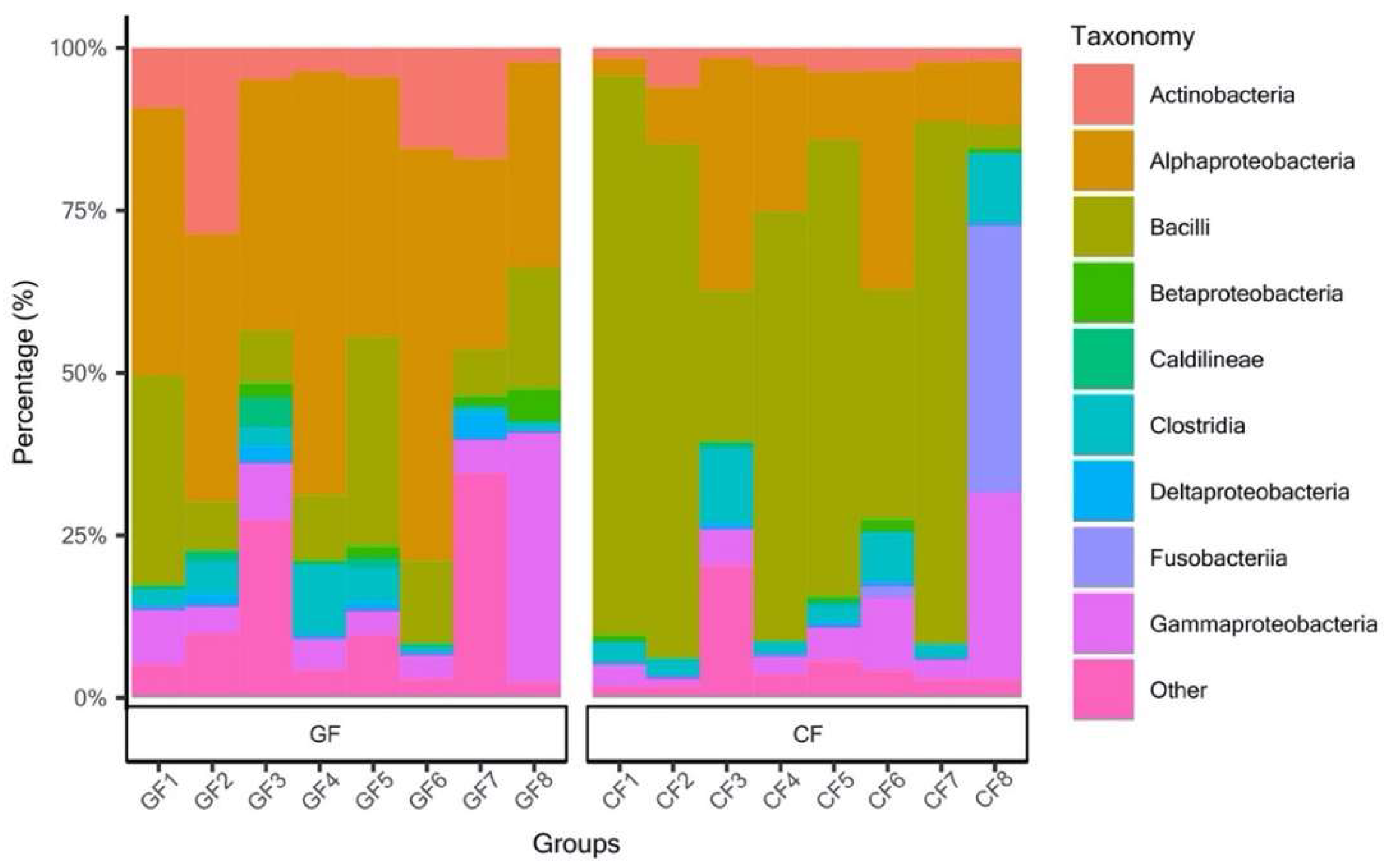
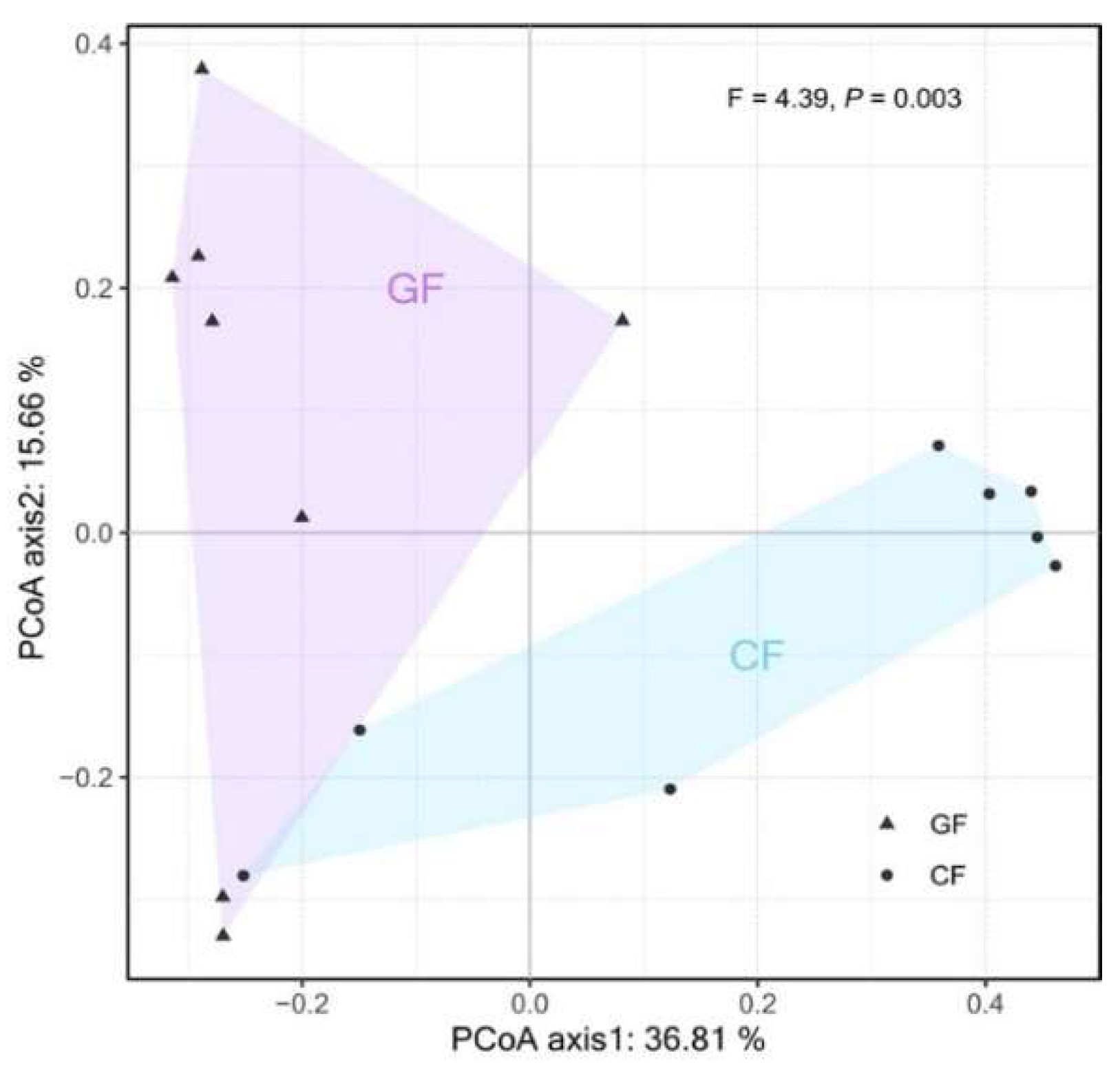
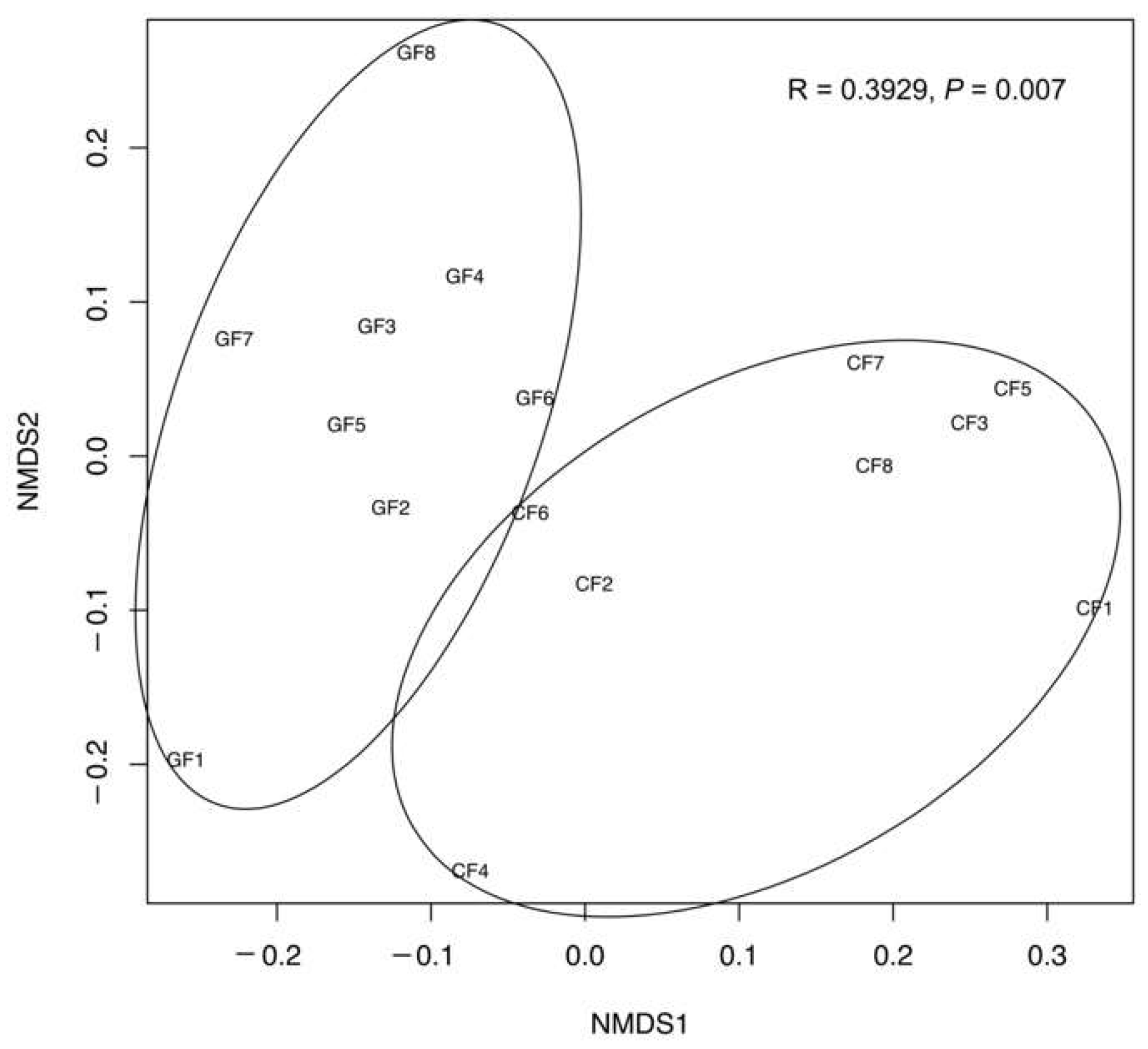
3.1. Diversity and Composition of the Bacterial Community
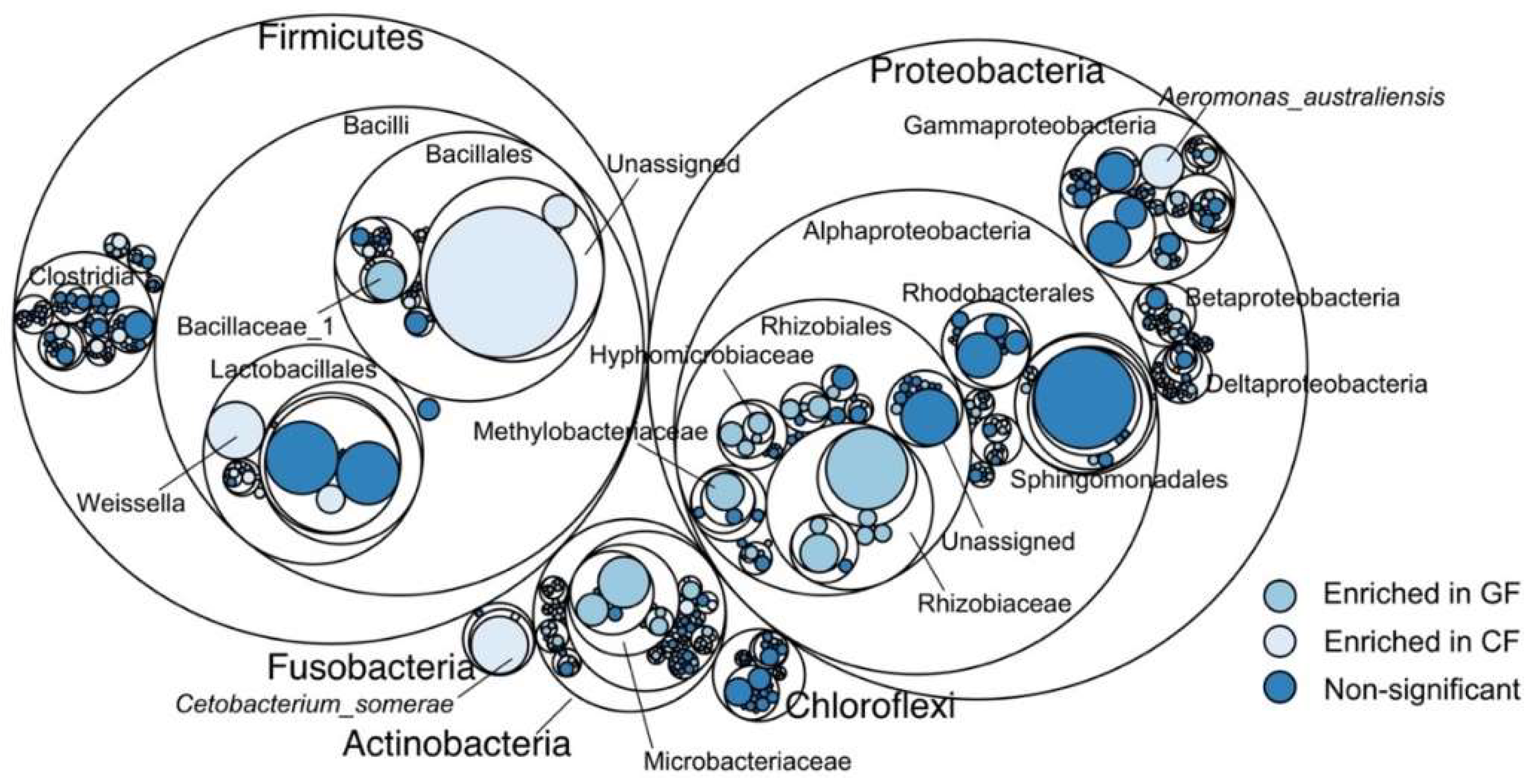
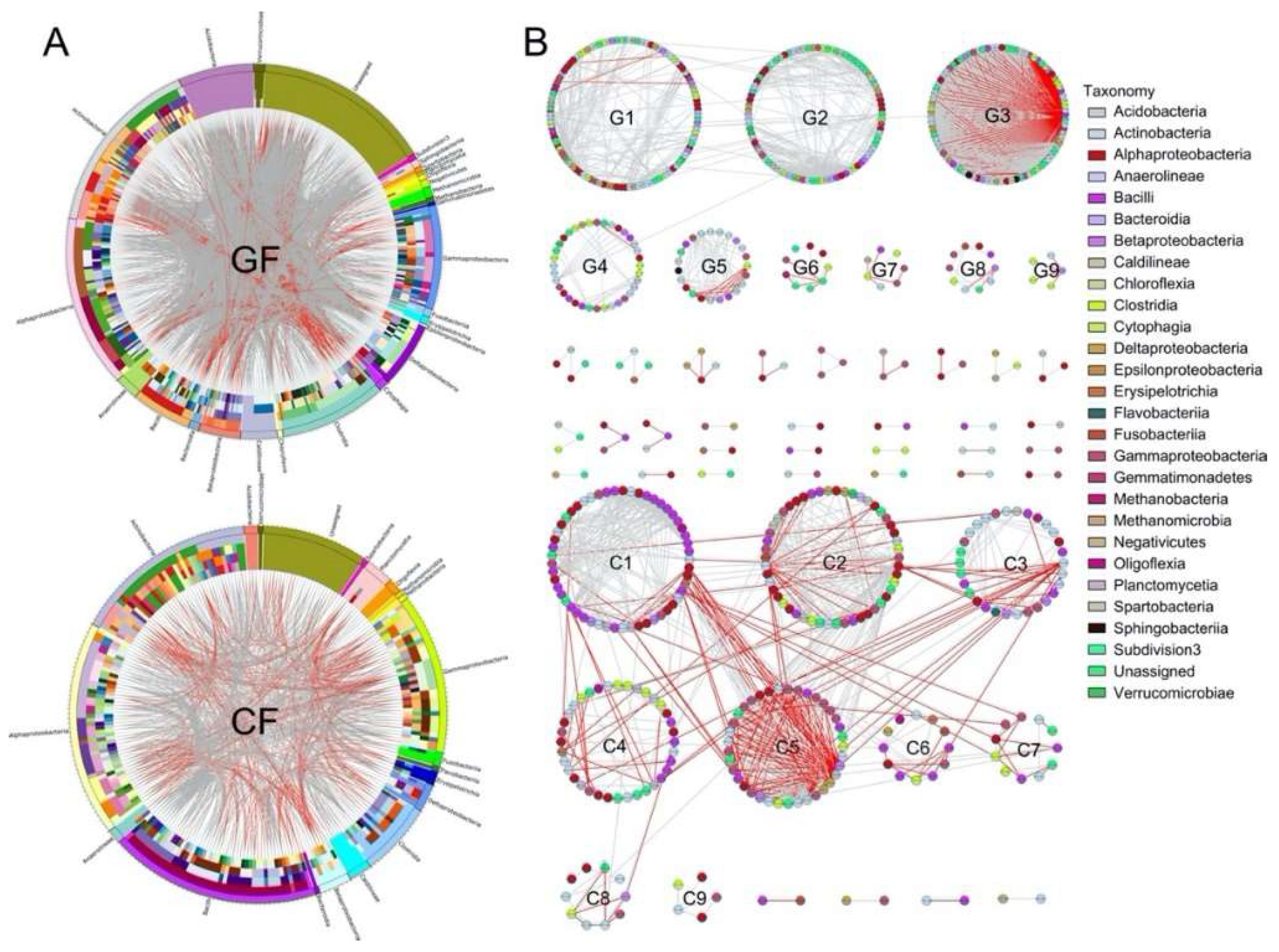
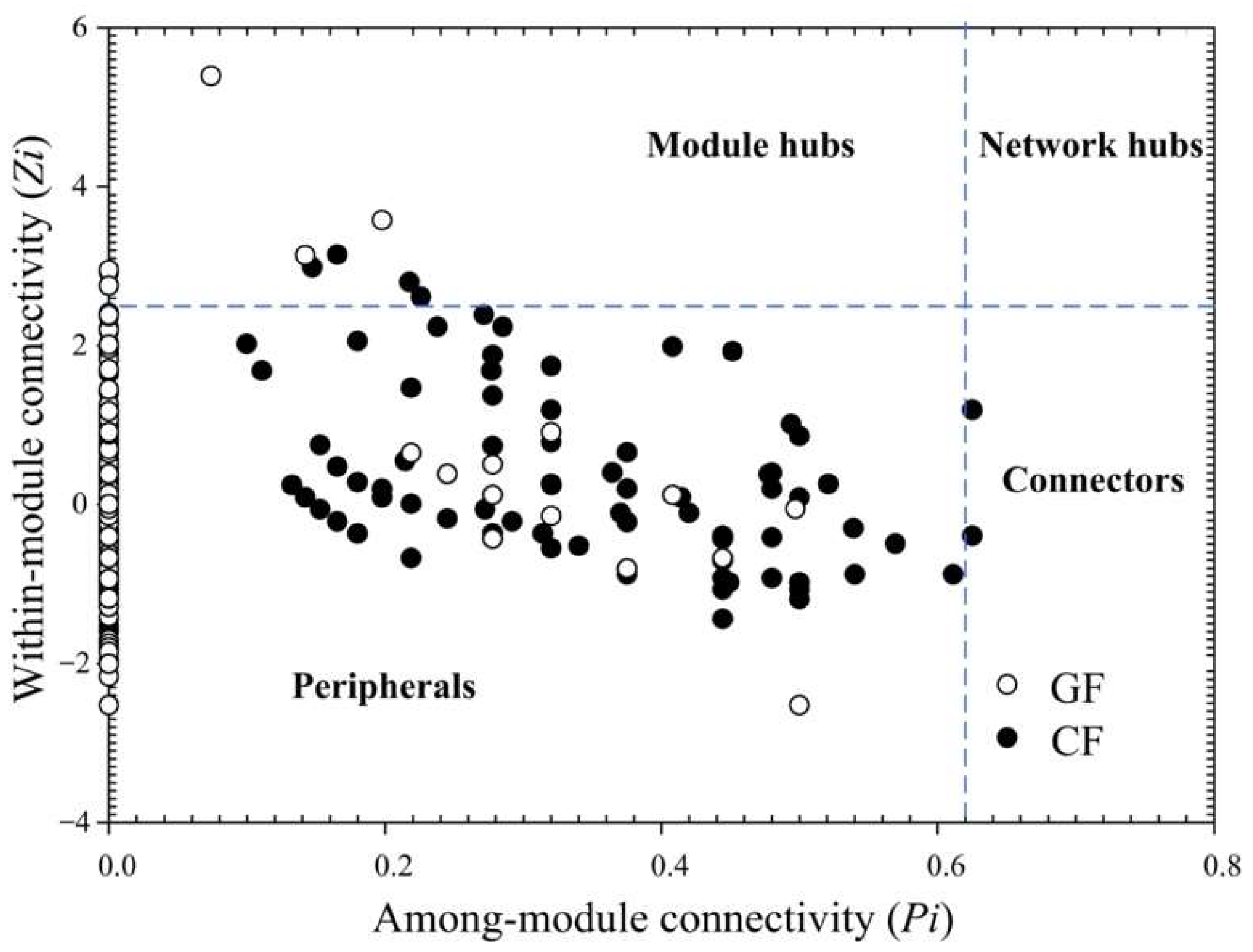
3.2. Ecological Network Analysis
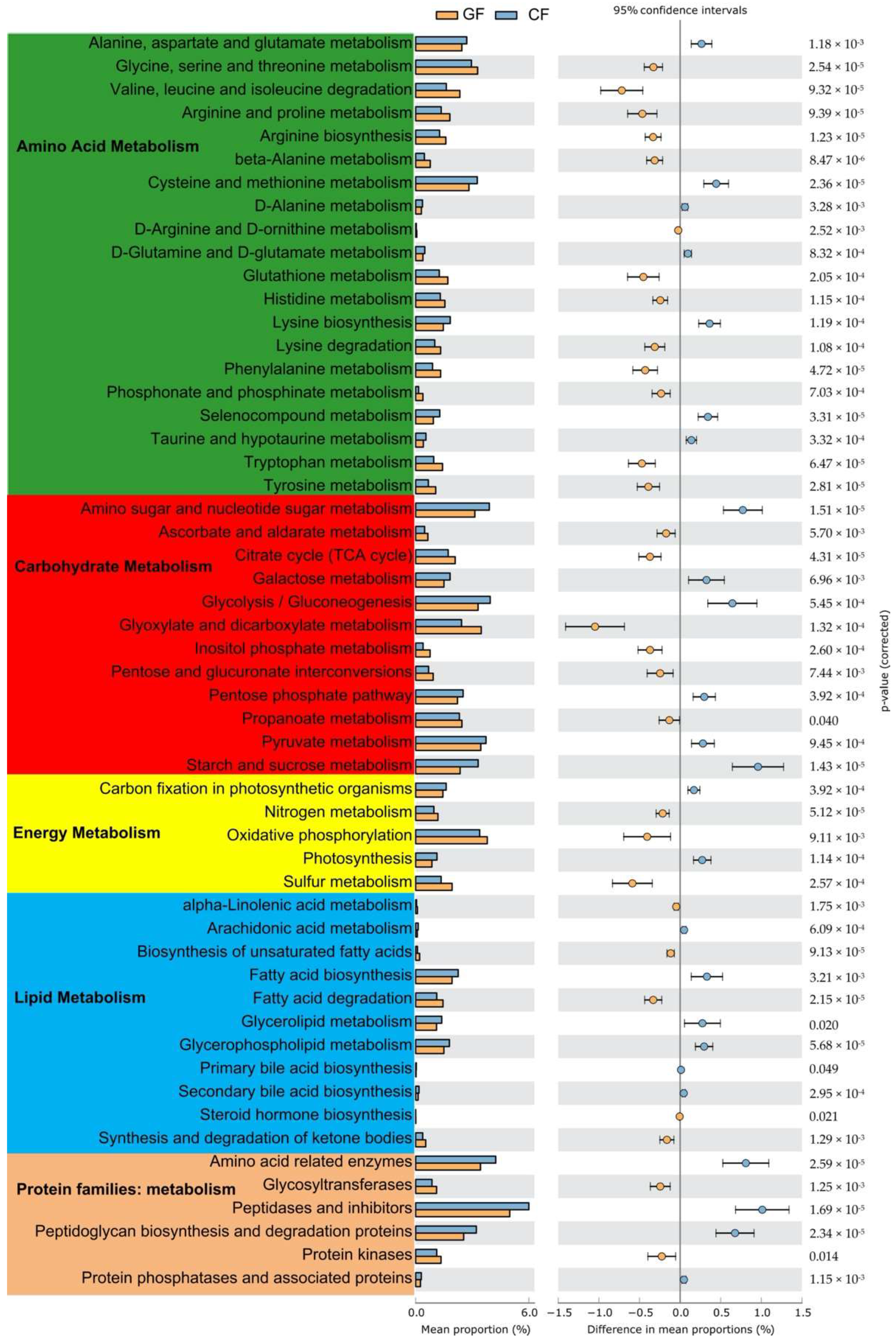
3.3. Functional Predictions of Intestinal Microbiota with PICRUSt2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tarnecki, A.M.; Burgos, F.A.; Ray, C.L.; Arias, C.R. Fish intestinal microbiome: Diversity and symbiosis unravelled by metagenomics. J. Appl. Microbiol. 2017, 123, 2–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish gut microbiome: Current approaches and future perspectives. Indian J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.S.; Shin, N.-R.; Lee, J.-B.; Kim, M.-S.; Whon, T.W.; Hyun, D.-W.; Yun, J.-H.; Jung, M.-J.; Kim, J.Y.; Bae, J.-W. Host habitat is the major determinant of the gut microbiome of fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.C.; Weese, J.S. Understanding the intestinal microbiome in health and disease. Vet. Clin. N. Am. Equine Pract. 2018, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dupont, H.L.; Jiang, Z.-D.; Dupont, A.W.; Utay, N.S. The intestinal microbiome in human health and disease. Trans. Am. Clin. Climatol. Assoc. 2020, 131, 178. [Google Scholar] [PubMed]
- Power, S.E.; O’Toole, P.W.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. Intestinal microbiota, diet and health. Br. J. Nutr. 2014, 111, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Guo, X.; Gooneratne, R.; Lai, R.; Zeng, C.; Zhan, F.; Wang, W. The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 2016, 6, 24340. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Ren, Y.; Peng, C.; Hao, Y.; Xiong, F.; Wang, G.; Li, W.; Zou, H.; Angert, E.R. Metatranscriptomic discovery of plant biomass-degrading capacity from grass carp intestinal microbiomes. FEMS Microbiol. Ecol. 2015, 91, fiv107. [Google Scholar] [CrossRef]
- Dearing, M.D.; Kohl, K.D. Beyond fermentation: Other important services provided to endothermic herbivores by their gut microbiota. Integr. Comp. Biol. 2017, 57, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.S.; Koller, K.R.; Ramaboli, M.C.; Nesengani, L.T.; Ocvirk, S.; Chen, C.; Flanagan, C.A.; Sapp, F.R.; Merritt, Z.T.; Bhatti, F. Diet and the human gut microbiome: An international review. Digest. Dis. Sci. 2020, 65, 723–740. [Google Scholar] [CrossRef]
- Agboola, J.O.; Chikwati, E.M.; Hansen, J.Ø.; Kortner, T.M.; Mydland, L.T.; Krogdahl, Å.; Djordjevic, B.; Schrama, J.W.; Øverland, M. A meta-analysis to determine factors associated with the severity of enteritis in Atlantic salmon (Salmo salar) fed soybean meal-based diets. Aquaculture 2022, 555, 738214. [Google Scholar] [CrossRef]
- Zhang, W.; Tan, B.; Deng, J.; Haitao, Z. Multiomics analysis of soybean meal induced marine fish enteritis in juvenile pearl gentian grouper, Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂. Sci. Rep. 2021, 11, 23319. [Google Scholar] [CrossRef] [PubMed]
- Infante-Villamil, S.; Huerlimann, R.; Jerry, D.R. Microbiome diversity and dysbiosis in aquaculture. Rev. Aquac. 2021, 13, 1077–1096. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Yang, G.; Jian, S.Q.; Cao, H.; Wen, C.; Hu, B.; Peng, M.; Peng, L.; Yuan, J.; Liang, L. Changes in microbiota along the intestine of grass carp (Ctenopharyngodon idella): Community, interspecific interactions, and functions. Aquaculture 2019, 498, 151–161. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, X.; Li, L. Human gut microbiome: The second genome of human body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clément, K. Metabolism and metabolic disorders and the microbiome: The intestinal microbiota associated with obesity, lipid metabolism, and metabolic health—Pathophysiology and therapeutic strategies. Gastroenterology 2021, 160, 573–599. [Google Scholar] [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.-H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Lin, R.; Liu, W.; Piao, M.; Zhu, H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 2017, 49, 2083–2090. [Google Scholar] [CrossRef]
- Nieuwdorp, M.; Gilijamse, P.W.; Pai, N.; Kaplan, L.M. Role of the microbiome in energy regulation and metabolism. Gastroenterology 2014, 146, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.T.; Zhang, J.; Xiong, F.; Wang, G.-T.; Li, W.-X.; Wu, S.-G. Altered gut microbiota associated with intestinal disease in grass carp (Ctenopharyngodon idellus). World J. Microb. Biot. 2018, 34, 71. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.-Y.; Yang, H.-X.; He, W.; Tian, D.-Y.; Kou, H.-Y.; Wu, K.; Yang, C.-G.; Cheng, Z.-Q.; Song, X.-H. A grass carp model with an antibiotic-disrupted intestinal microbiota. Aquaculture 2021, 541, 736790. [Google Scholar] [CrossRef]
- Zhang, J.; Xiong, F.; Wang, G.T.; Li, W.X.; Li, M.; Zou, H.; Wu, S.G. The influence of diet on the grass carp intestinal microbiota and bile acids. Aquac. Res. 2017, 48, 4934–4944. [Google Scholar] [CrossRef]
- Wu, S.; Wang, G.; Angert, E.R.; Wang, W.; Li, W.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, e30440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, N.T.; Wang, G.T.; Wu, S.G. A review of intestinal microbes in grass carp Ctenopharyngodon idellus (Valenciennes). Aquac. Res. 2017, 48, 3287–3297. [Google Scholar] [CrossRef]
- Yang, G.; Xu, Z.; Tian, X.; Dong, S.; Peng, M. Intestinal microbiota and immune related genes in sea cucumber (Apostichopus japonicus) response to dietary β-glucan supplementation. Biochem. Biophys. Res. Commun. 2015, 458, 98–103. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; De Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.; Gomez-Exposito, R.; Elsayed, S.S. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational multivariate analysis of variance. Dep. Stat. Univ. Auckl. Auckl. 2015, 26, 32–46. [Google Scholar]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinf. 2012, 13, 113. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Mallott, E.K.; Amato, K.R. Host specificity of the gut microbiome. Nat. Rev. Microbiol. 2021, 19, 639–653. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Zhang, J.; Jakovlić, I.; Xiong, F.; Wu, S.; Zou, H.; Li, W.; Li, M.; Wang, G. Gut segments outweigh the diet in shaping the intestinal microbiota composition in grass carp Ctenopharyngodon idellus. AMB Express 2019, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Fečkaninová, A.; Koščová, J.; Mudroňová, D.; Popelka, P.; Toropilova, J. The use of probiotic bacteria against Aeromonas infections in salmonid aquaculture. Aquaculture 2017, 469, 1–8. [Google Scholar] [CrossRef]
- Igbinosa, I.H.; Beshiru, A.; Odjadjare, E.E.; Ateba, C.N.; Igbinosa, E.O. Pathogenic potentials of Aeromonas species isolated from aquaculture and abattoir environments. Microb. Pathogen. 2017, 107, 185–192. [Google Scholar] [CrossRef]
- Song, X.; Zhao, J.; Bo, Y.; Liu, Z.; Wu, K.; Gong, C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): An experimental model. Aquaculture 2014, 434, 171–178. [Google Scholar] [CrossRef]
- Song, X.; Hu, X.; Sun, B.; Bo, Y.; Wu, K.; Xiao, L.; Gong, C. A transcriptome analysis focusing on inflammation-related genes of grass carp intestines following infection with Aeromonas hydrophila. Sci. Rep. 2017, 7, 40777. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yang, Y.; Liu, J.; Awan, F.; Lu, C.; Liu, Y. Inhibition of Aeromonas hydrophila-induced intestinal inflammation and mucosal barrier function damage in crucian carp by oral administration of Lactococcus lactis. Fish Shellfish Immunol. 2018, 83, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Gewaily, M.S.; Shukry, M.; Abdel-Kader, M.F.; Alkafafy, M.; Farrag, F.A.; Moustafa, E.M.; Doan, H.V.; Abd-Elghany, M.F.; Abdelhamid, A.F.; Eltanahy, A. Dietary Lactobacillus plantarum relieves Nile tilapia (Oreochromis niloticus) juvenile from oxidative stress, immunosuppression, and inflammation induced by deltamethrin and Aeromonas hydrophila. Front. Mar. Sci. 2021, 8, 621558. [Google Scholar]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Yang, Y. Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2. mBio 2011, 2, e00122-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pande, S.; Kaftan, F.; Lang, S.; Svatoš, A.; Germerodt, S.; Kost, C. Privatization of cooperative benefits stabilizes mutualistic cross-feeding interactions in spatially structured environments. ISME J. 2016, 10, 1413–1423. [Google Scholar] [CrossRef]
- Ren, X.; Murray, R.M. Cooperation enhances robustness of coexistence in spatially structured consortia. In Proceedings of the 2019 18th European Control Conference (ECC), Naples, Italy, 25–28 June 2019; pp. 2651–2656. [Google Scholar]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.; Sugrue, I.; Arendt, E.; Hill, C.; Stanton, C.; Ross, R.P. Recent advances in microbial fermentation for dairy and health. F1000Research 2017, 6, 751. [Google Scholar] [CrossRef] [Green Version]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Shefat, S.H.T.; Karim, M.A. Nutritional diseases of fish in aquaculture and their management: A review. Acta Sci. Pharm. Sci. 2018, 2, 50–58. [Google Scholar]
- Yao, T.; Gu, X.; Liang, X.; Fall, F.N.; Cao, A.; Zhang, S.; Guan, Y.; Sun, B.; Xue, M. Tolerance assessment of dietary bile acids in common carp (Cyprinus carpio L.) fed a high plant protein diet. Aquaculture 2021, 543, 737012. [Google Scholar] [CrossRef]
- Million, M.; Lagier, J.-C.; Yahav, D.; Paul, M. Gut bacterial microbiota and obesity. Clin. Microbiol. Infect. 2013, 19, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoun, A.; Darwish, F.; Hamod, N. The influence of the gut microbiome on obesity in adults and the role of probiotics, prebiotics, and synbiotics for weight loss. Prev. Nutr. Food Sci. 2020, 25, 113. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Factors influencing the grass carp gut microbiome and its effect on metabolism. FEMS Microbiol. Ecol. 2014, 87, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Leo, V.V.; Asem, D.; Singh, B.P. Actinobacteria: A highly potent source for holocellulose degrading enzymes. In Actinobacteria, New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, J.S., Singh, D.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 191–205. ISBN 978-0-444-63994-3. [Google Scholar]
- Smid, E.; Lacroix, C. Microbe–microbe interactions in mixed culture food fermentations. Curr. Opin. Biotechnol. 2013, 24, 148–154. [Google Scholar] [CrossRef]
- Beltrán, J.; Esteban, M. Nature-identical compounds as feed additives in aquaculture. Fish Shellfish Immunol. 2022, 123, 409–416. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; Martínez-Córdova, L.; Hernández-Mendoza, A.; Cicala, F.; Lago-Lestón, A.; Martínez-Porchas, M. Therapeutic modulation of fish gut microbiota, a feasible strategy for aquaculture? Aquaculture 2021, 544, 737050. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Observed OTUs | Chao1 | ACE | Shannon | InvSimpson |
|---|---|---|---|---|---|
| GF | 518.88 ± 21.96 b | 618.72 ± 16.8 | 605.7 ± 17.79 b | 3.78 ± 0.23 b | 16.8 ± 3.49 b |
| CF | 304.13 ± 41.13 a | 351.08 ± 51.7 | 344.53 ± 50.91 a | 2.76 ± 0.2 a | 5.86 ± 1.35 a |
| Index | GF | CF |
|---|---|---|
| Acidobacteria | 29 | 3 |
| Actinobacteria | 69 | 40 |
| Alphaproteobacteria | 66 | 47 |
| Anaerolineae | 9 | 3 |
| Bacilli | 23 | 46 |
| Bacteroidia | 4 | 1 |
| Betaproteobacteria | 16 | 7 |
| Caldilineae | 14 | 5 |
| Chloroflexia | 2 | 0 |
| Clostridia | 41 | 16 |
| Cytophagia | 4 | 0 |
| Deltaproteobacteria | 25 | 6 |
| Epsilonproteobacteria | 1 | 0 |
| Erysipelotrichia | 3 | 3 |
| Flavobacteriia | 0 | 1 |
| Fusobacteriia | 3 | 3 |
| Gammaproteobacteria | 40 | 35 |
| Gemmatimonadetes | 1 | 0 |
| Methanomicrobia | 4 | 0 |
| Methanobacteria | 2 | 2 |
| Negativicutes | 3 | |
| Oligoflexia | 2 | 3 |
| Planctomycetia | 1 | 6 |
| Spartobacteria | 2 | 1 |
| Sphingobacteriia | 3 | 0 |
| Subdivision3 | 2 | 0 |
| Verrucomicrobiae | 4 | 1 |
| Unassigned | 68 | 23 |
| Total number of OTUs | 441 | 252 |
| The number of modules (≥5 OTUs) | 9 | 9 |
| The number of module hubs | 7 | 4 |
| The number of connectors | 0 | 3 |
| The number of gray edges | 2099 | 592 |
| The number of red edges | 847 | 264 |
| Total number of edges | 2946 | 856 |
| Treatment | Topological Roles | OTUs | Module Number | Phylogenetic Associations |
|---|---|---|---|---|
| GF | Module hubs | OTU_376 | 1 | Erysipelotrichia |
| Module hubs | OTU_568 | 4 | Actinobacteria | |
| Module hubs | OTU_891 | 2 | Alphaproteobacteria | |
| Module hubs | OTU_756 | 2 | Chloroflexi | |
| Module hubs | OTU_153 | 4 | Gammaproteobacteria | |
| Module hubs | OTU_901 | 1 | Alphaproteobacteria | |
| Module hubs | OTU_911 | 1 | Alphaproteobacteria | |
| CF | Module hubs | OTU_549 | 3 | Actinobacteria |
| Module hubs | OTU_1171 | 2 | Clostridia | |
| Module hubs | OTU_892 | 2 | Alphaproteobacteria | |
| Module hubs | OTU_149 | 2 | Gammaproteobacteria | |
| Connectors | OTU_522 | 4 | Planctomycetia | |
| Connectors | OTU_894 | 4 | Alphaproteobacteria | |
| Connectors | OTU_376 | 4 | Erysipelotrichia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Xiang, Y.; Wang, S.; Tao, Y.; Xie, L.; Bao, L.; Shen, K.; Li, J.; Hu, B.; Wen, C.; et al. Response of Intestinal Microbiota to the Variation in Diets in Grass Carp (Ctenopharyngodon idella). Metabolites 2022, 12, 1115. https://doi.org/10.3390/metabo12111115
Yang G, Xiang Y, Wang S, Tao Y, Xie L, Bao L, Shen K, Li J, Hu B, Wen C, et al. Response of Intestinal Microbiota to the Variation in Diets in Grass Carp (Ctenopharyngodon idella). Metabolites. 2022; 12(11):1115. https://doi.org/10.3390/metabo12111115
Chicago/Turabian StyleYang, Gang, Yuhan Xiang, Shanshan Wang, Yujie Tao, Lichen Xie, Lixin Bao, Kaikai Shen, Jiamin Li, Baoqing Hu, Chungen Wen, and et al. 2022. "Response of Intestinal Microbiota to the Variation in Diets in Grass Carp (Ctenopharyngodon idella)" Metabolites 12, no. 11: 1115. https://doi.org/10.3390/metabo12111115
APA StyleYang, G., Xiang, Y., Wang, S., Tao, Y., Xie, L., Bao, L., Shen, K., Li, J., Hu, B., Wen, C., Kumar, V., & Peng, M. (2022). Response of Intestinal Microbiota to the Variation in Diets in Grass Carp (Ctenopharyngodon idella). Metabolites, 12(11), 1115. https://doi.org/10.3390/metabo12111115