
Exposure to Environmentally Relevant Levels of PFAS Causes Metabolic Changes in the Freshwater Amphipod Austrochiltonia subtenuis
 , ,
, , 
Abstract
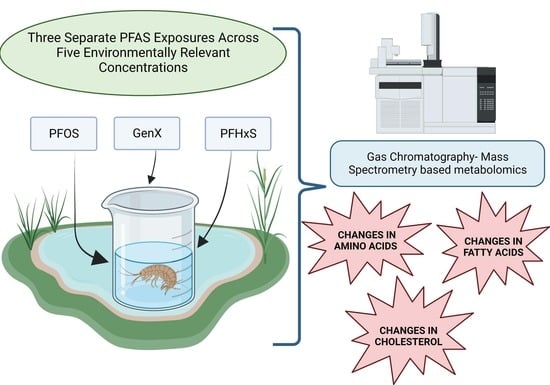
:
1. Introduction
2. Materials and Methods
2.1. Organism
2.2. Chemical and Concentrations
2.3. PFAS Exposure
2.4. Sample Preparation and Metabolite Extraction
2.5. Gas Chromatography–Mass Spectrometry
2.6. PFAS Determination Using Liquid Chromatography—Mass Spectrometry
2.7. Bioconcentration Factors (BCFs)
2.8. Statistical Analysis—Survival
2.9. Metabolite Data Analysis
3. Results and Discussion
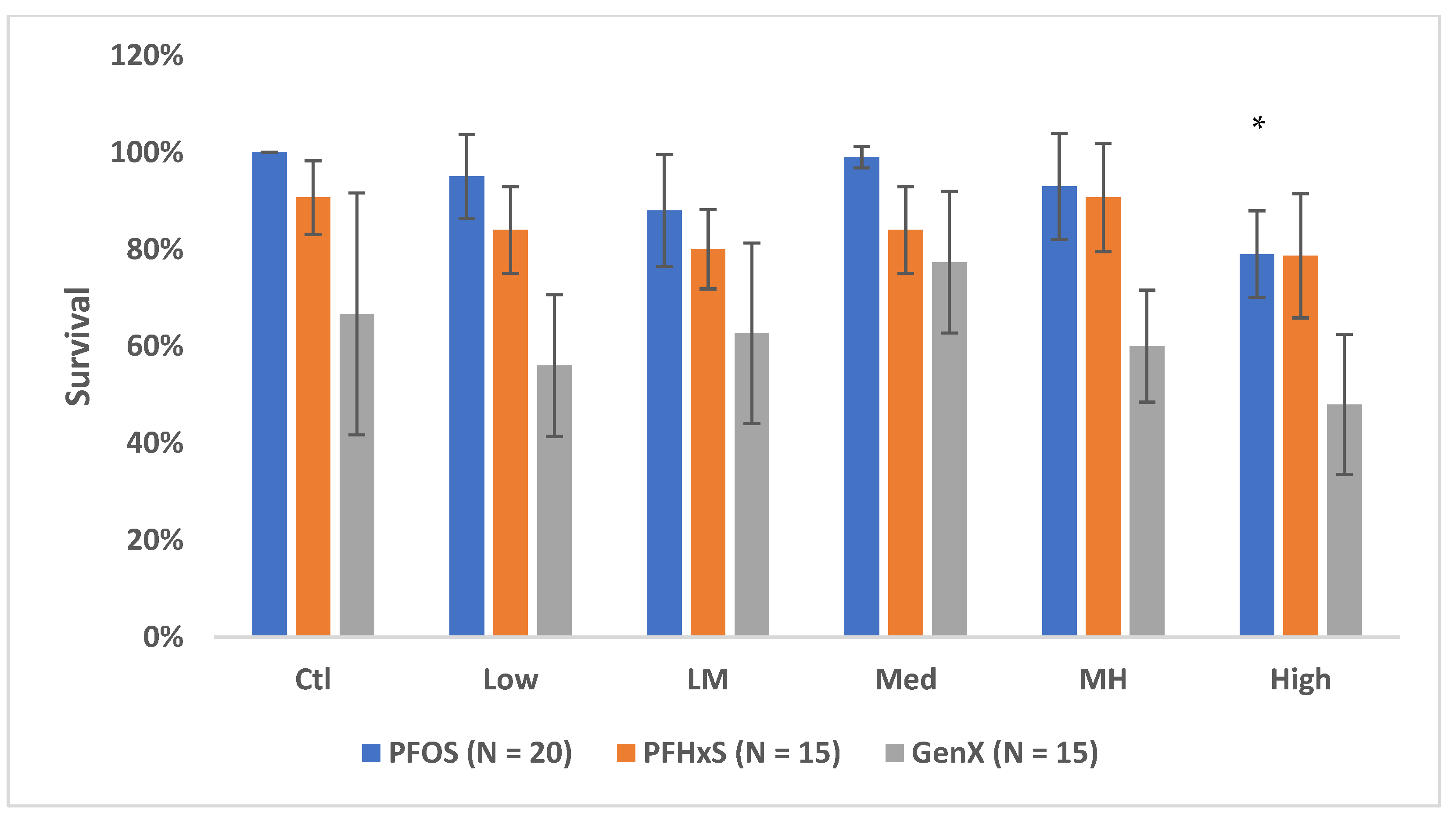
3.1. Survival
3.2. Metabolomics
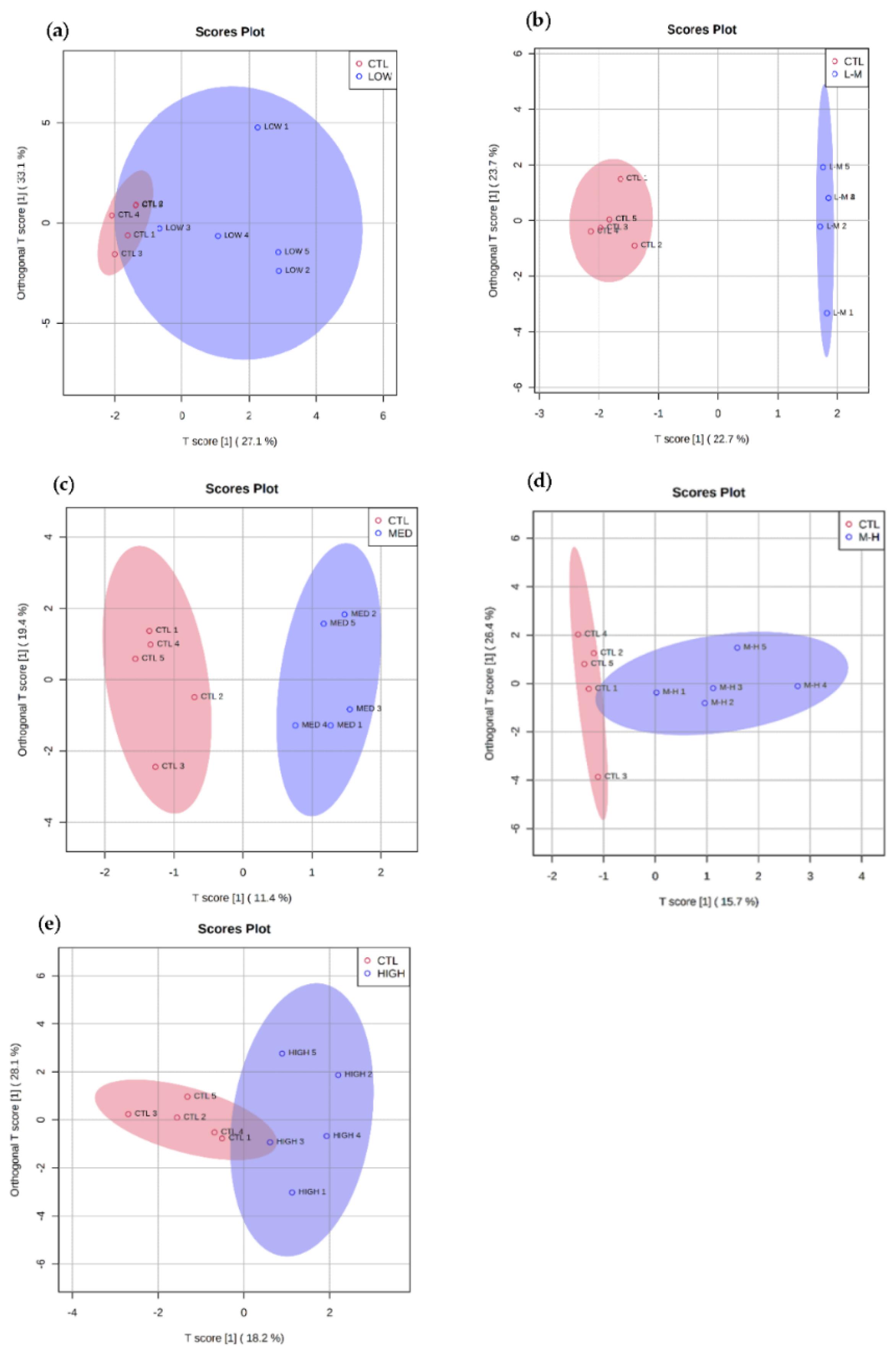
3.3. PFOS Exposure
3.3.1. Multivariate Data Analysis for PFOS Exposure
3.3.2. Univariate Data Analysis for PFOS Exposure
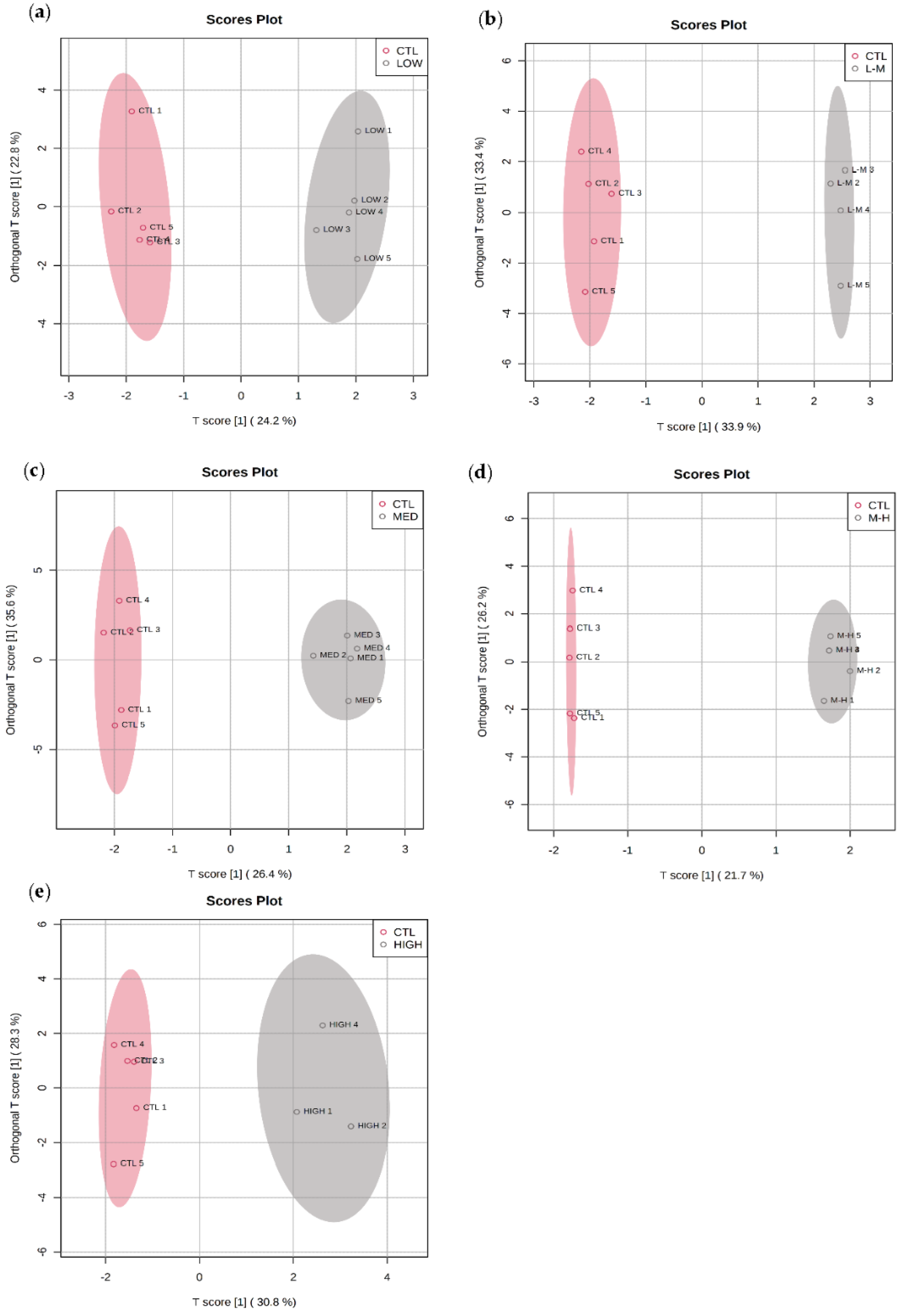
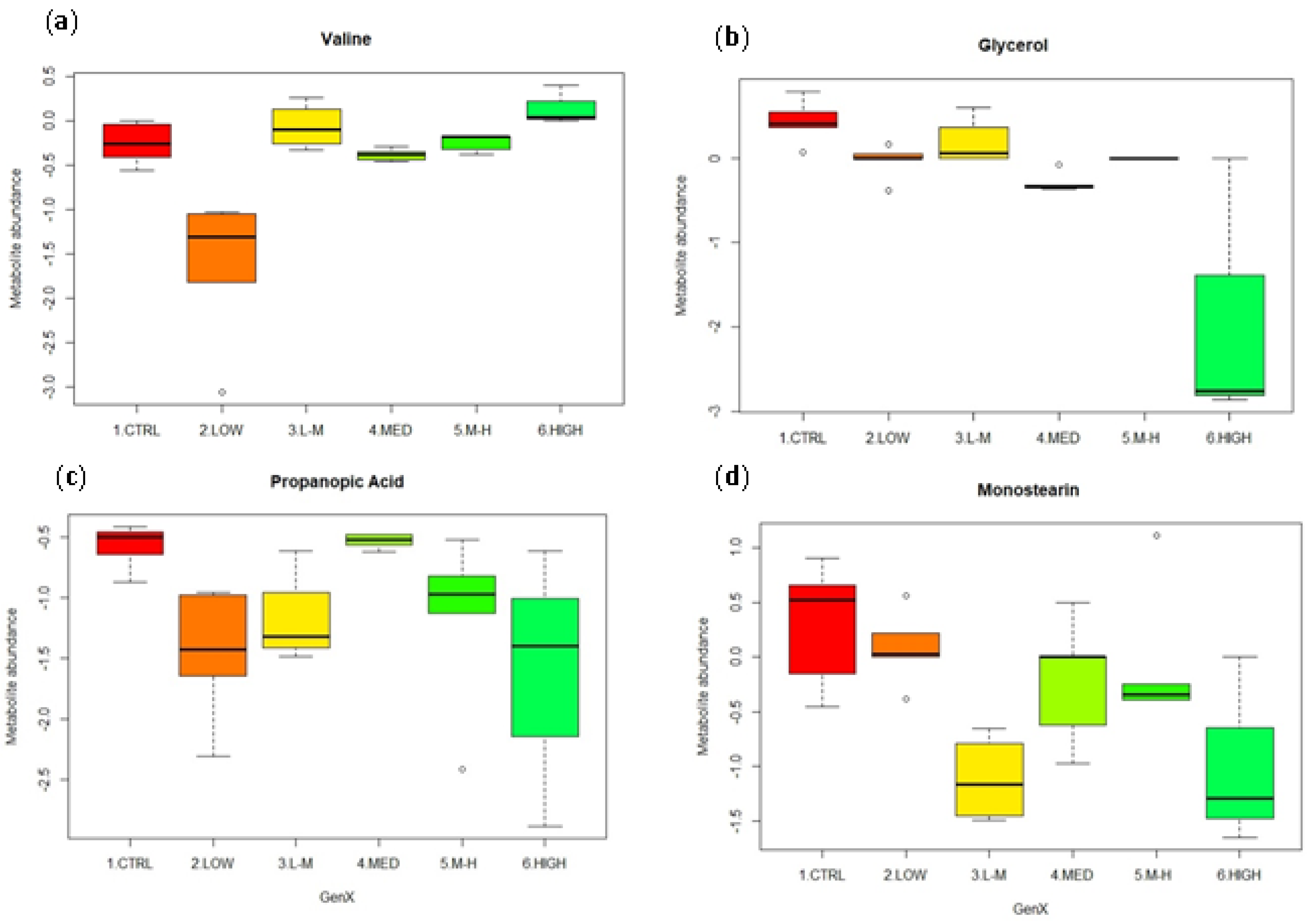
3.4. GenX Exposure
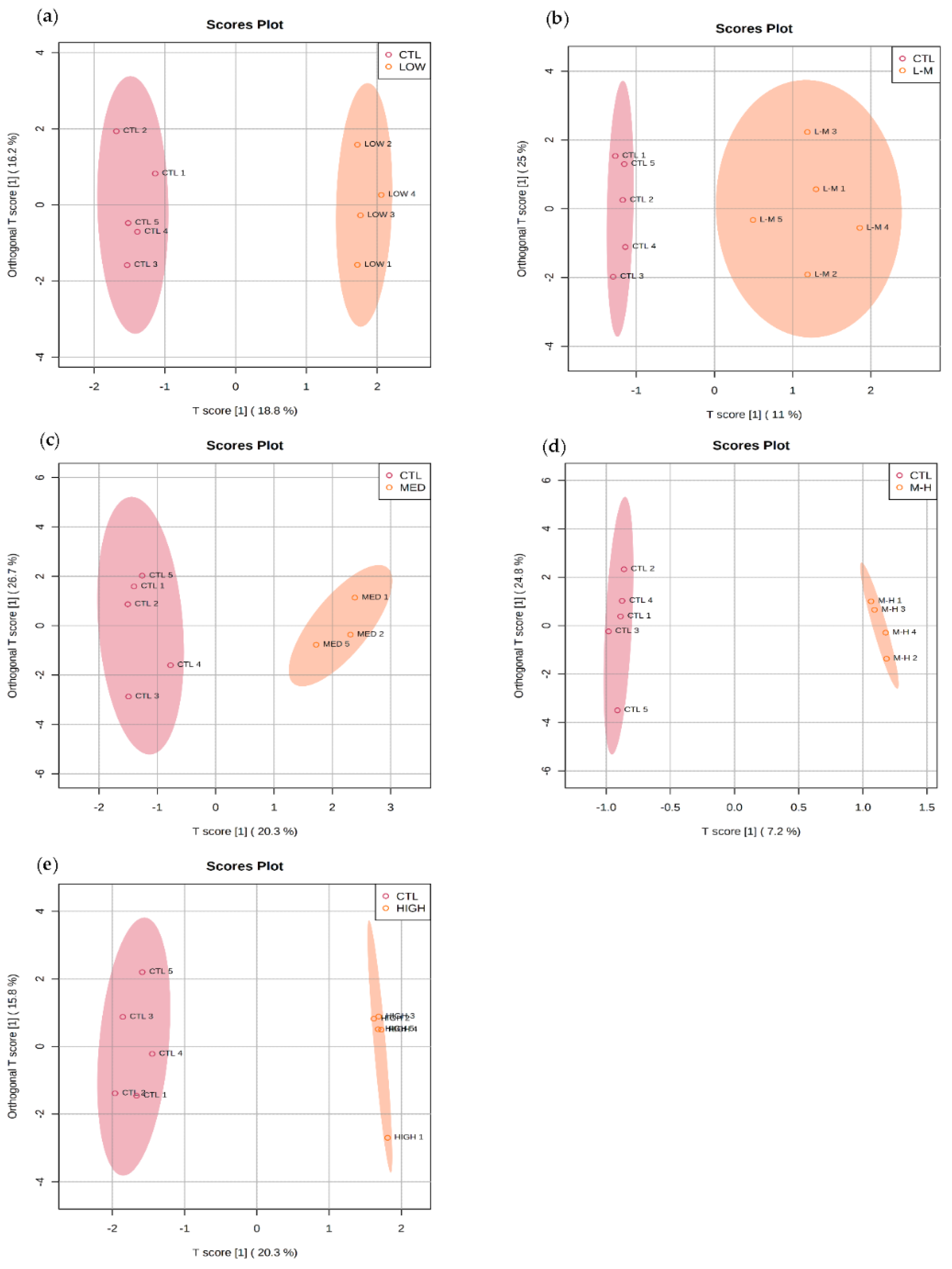
3.4.1. Multivariate Data Analysis for GenX Exposure
3.4.2. Univariate Data Analysis for GenX Exposure
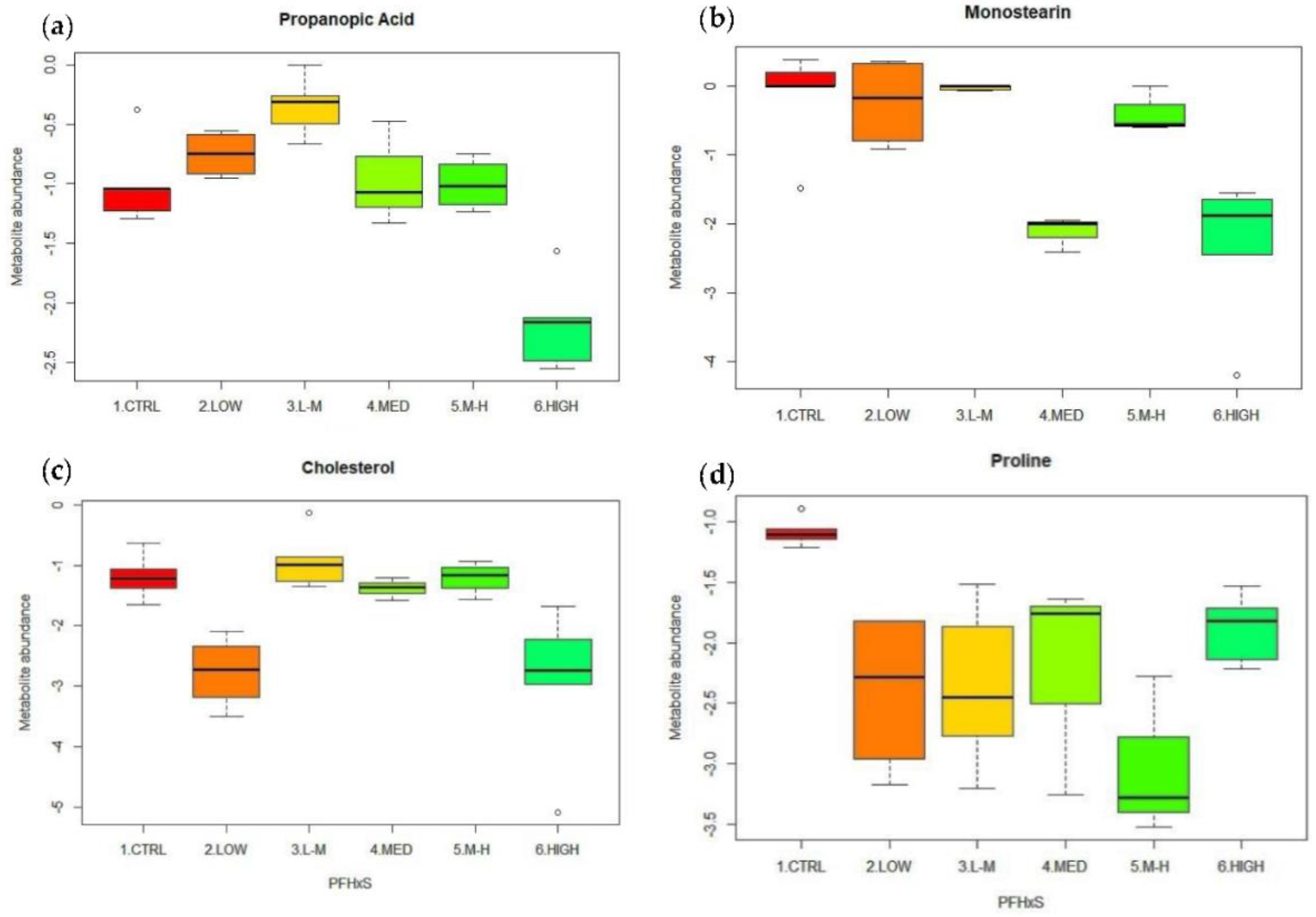
3.5. PFHxS Exposure
3.5.1. Multivariate Data Analysis for PFHxS Exposure
3.5.2. Univariate Data Analysis for PFHxS Exposure
3.6. Overall PFAS Effect on Metabolites
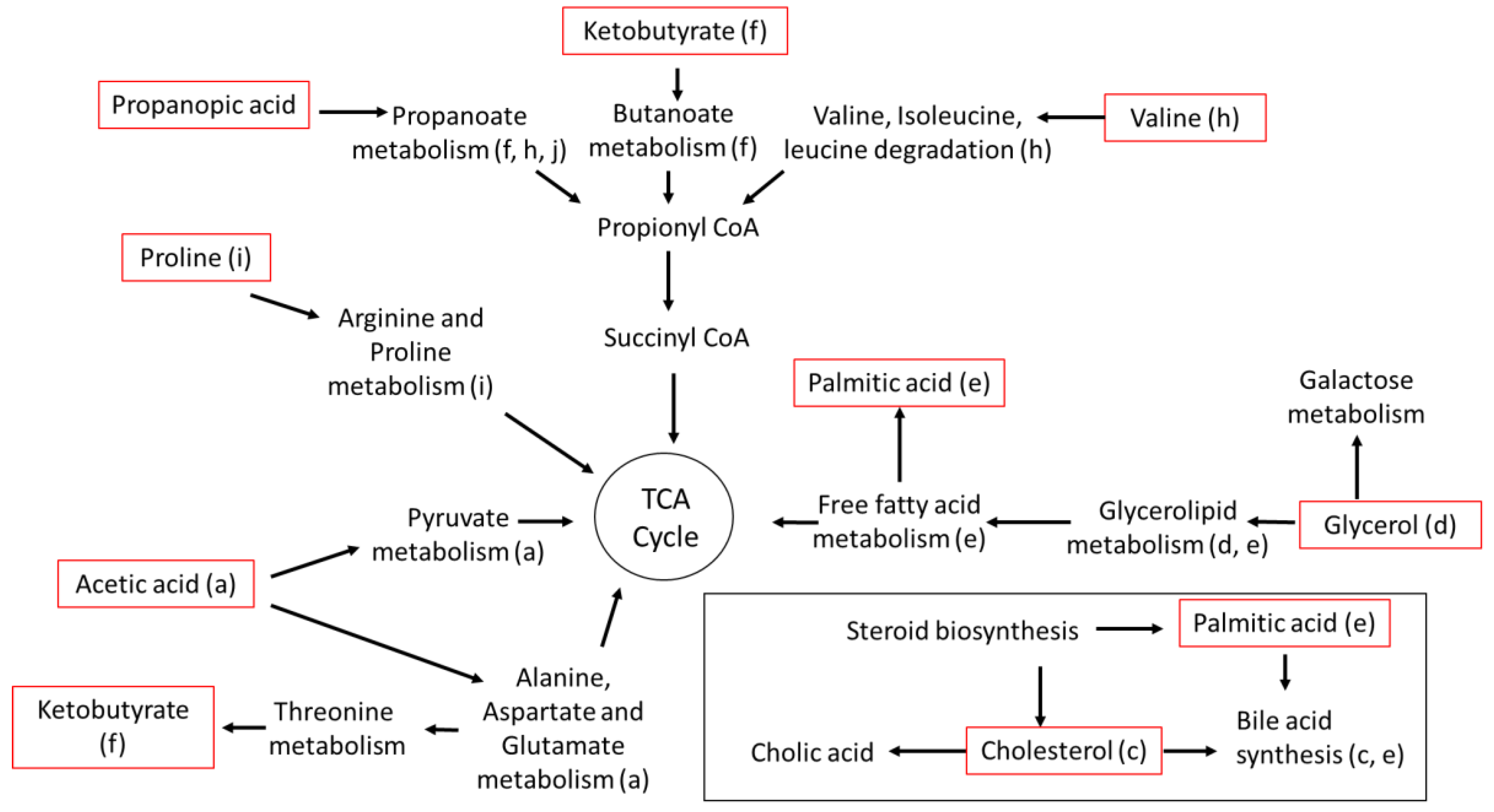
3.7. Metabolomic Pathways
3.8. PFAS Water Concentration
3.9. Bioconcentration Factors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pelch, K.E.; Reade, A.; Wolffe, T.A.M.; Kwiatkowski, C.F. PFAS Health Effects Database: Protocol for a Systematic Evidence Map. Environ. Int. 2019, 130, 104851. [Google Scholar] [CrossRef] [PubMed]
- Heads of Environmental Protection Agency Australia and New Zealand (HEPA) PFAS National Environmental Management Plan. Available online: https://www.dcceew.gov.au/sites/default/files/documents/pfas-nemp-2.pdf (accessed on 8 August 2019).
- Sinclair, G.M.; Long, S.M.; Jones, O.A.H. What Are the Effects of PFAS Exposure at Environmentally Relevant Concentrations? Chemosphere 2020, 258, 127340. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Hillyer, K.; Nilsson, S.; Limpus, D.; Bose, U.; Broadbent, J.A.; Vardy, S. Bioaccumulation and Metabolic Response of PFAS Mixtures in Wild-Caught Freshwater Turtles (Emydura macquarii macquarii) Using Omics-Based Ecosurveillance Techniques. Sci. Total Environ. 2021, 806, 151264. [Google Scholar] [CrossRef]
- Beale, D.J.; Sinclair, G.M.; Shah, R.; Paten, A.M.; Kumar, A.; Long, S.M.; Vardy, S.; Jones, O.A.H. A Review of Omics-Based PFAS Exposure Studies Reveals Common Biochemical Response Pathways. Sci. Total Environ. 2022, 845, 157255. [Google Scholar] [CrossRef] [PubMed]
- Jones, O.A.H.; Maguire, M.L.; Griffin, J.L.; Dias, D.A.; Spurgeon, D.J.; Svendsen, C. Metabolomics and Its Use in Ecology. Austral Ecol. 2013, 38, 713–720. [Google Scholar] [CrossRef]
- Buckley, N.; Sim, M.; Douglas, K.; Håkansson, H. Expert Health Panel for Per- and Poly-Fluoroalkyl Substances (PFAS). Available online: https://www.health.gov.au/internet/main/publishing.nsf/Content/C9734ED6BE238EC0CA2581BD00052C03/$File/expert-panel-report.pdf (accessed on 30 May 2019).
- Jones, O.A.H. Illuminating the Dark Metabolome to Advance the Molecular Characterisation of Biological Systems. Metabolomics 2018, 14, 101. [Google Scholar] [CrossRef]
- Yao, X.; Cao, D.; Wang, F.; Zhang, W.; Ma, C.; Song, M. An Overview of Omics Approaches to Characterize the Effect of Perfluoroalkyl Substances in Environmental Health. TrAC—Trends Anal. Chem. 2019, 121, 115367. [Google Scholar] [CrossRef]
- Hogue, C. Governments Agree to Eliminate PFHxS. C&EN Glob. Enterp. 2022, 100, 14. [Google Scholar]
- Brandsma, S.H.; Koekkoek, J.C.; van Velzen, M.J.M.; de Boer, J. The PFOA Substitute GenX Detected in the Environment near a Fluoropolymer Manufacturing Plant in the Netherlands. Chemosphere 2019, 220, 493–500. [Google Scholar] [CrossRef]
- Xiao, F. Emerging Poly- and Perfluoroalkyl Substances in the Aquatic Environment: A Review of Current Literature. Water Res. 2017, 124, 482–495. [Google Scholar] [CrossRef]
- Hassell, K.L.; Coggan, T.L.; Cresswell, T.; Kolobaric, A.; Berry, K.; Crosbie, N.D.; Blackbeard, J.; Pettigrove, V.J.; Clarke, B.O. Dietary Uptake and Depuration Kinetics of Perfluorooctane Sulfonate, Perfluorooctanoic Acid, and Hexafluoropropylene Oxide Dimer Acid (GenX) in a Benthic Fish. Environ. Toxicol. Chem. 2020, 39, 595–603. [Google Scholar] [CrossRef]
- Jeppe, K.J.; Kellar, C.R.; Marshall, S.; Colombo, V.; Sinclair, G.M.; Pettigrove, V. Bifenthrin Causes Toxicity in Urban Stormwater Wetlands: Field and Laboratory Assessment Using Austrochiltonia (Amphipoda). Environ. Sci. Technol. 2017, 51, 7254–7262. [Google Scholar] [CrossRef] [PubMed]
- Borgmann, U. Systematic Analysis of Aqueous Ion Requirements of Hyalella Azteca: A Standard Artificial Medium Including the Essential Bromide Ion. Arch. Environ. Contam. Toxicol 1996, 188, 356–363. [Google Scholar] [CrossRef]
- Morrison, N.; Dan, A.E.; Ae, B.; Bundy, J.G.; Tim, A.E.; Ae, C.; Currie, F.; Matthew, A.E.; Davey, P.; Nathan, A.E.; et al. Standard Reporting Requirements for Biological Samples in Metabolomics Experiments: Environmental Context. Metabolomics 2007, 3, 203–210. [Google Scholar] [CrossRef]
- Coggan, T.L.; Anumol, T.; Pyke, J.; Shimeta, J.; Clarke, B.O. A Single Analytical Method for the Determination of 53 Legacy and Emerging Per- and Polyfluoroalkyl Substances (PFAS) in Aqueous Matrices. Anal. Bioanal. Chem. 2019, 411, 3507–3520. [Google Scholar] [CrossRef]
- Lacalle-Bergeron, L.; Izquierdo-Sandoval, D.; Sancho, J.V.; López, F.J.; Hernández, F.; Portolés, T. Chromatography Hyphenated To High Resolution Mass Spectrometry In Untargeted Metabolomics For Investigation Of Food (BIO)Markers. TrAC Trends Anal. Chem. 2020, 135, 116161. [Google Scholar] [CrossRef]
- Sinclair, G.M.; O’Brien, A.L.; Keough, M.; De Souza, D.P.; Dayalan, S.; Kanojia, K.; Kouremenos, K.; Tull, D.L.; Coleman, R.A.; Jones, O.A.H.; et al. Using Metabolomics to Assess the Sub-Lethal Effects of Zinc and Boscalid on an Estuarine Polychaete Worm over Time. Metabolomics 2019, 15, 108. [Google Scholar] [CrossRef]
- Blasco, H.; Błaszczyński, J.; Billaut, J.C.; Nadal-Desbarats, L.; Pradat, P.F.; Devos, D.; Moreau, C.; Andres, C.R.; Emond, P.; Corcia, P.; et al. Comparative Analysis of Targeted Metabolomics: Dominance-Based Rough Set Approach versus Orthogonal Partial Least Square-Discriminant Analysis. J. Biomed. Inform. 2015, 53, 291–299. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.or (accessed on 15 January 2021).
- Simpson, S.L.; Liu, Y.; Spadaro, D.A.; Wang, X.; Kookana, R.S.; Batley, G.E. Chronic Effects and Thresholds for Estuarine and Marine Benthic Organism Exposure to Perfluorooctane Sulfonic Acid (PFOS )–Contaminated Sediments: Influence of Organic Carbon and Exposure Routes. Sci. Total Environ. 2021, 776, 146008. [Google Scholar] [CrossRef]
- Krupa, P.M.; Lotufo, G.R.; Mylroie, E.J.; May, L.K.; Gust, K.A.; Kimble, A.N.; Jung, M.G.; Boyda, J.A.; Moore, W. Chronic Aquatic Toxicity of Perfluorooctane Sulfonic Acid (PFOS) to Ceriodaphnia Dubia, Chironomus Dilutus, Danio Rerio, and Hyalella Azteca. Ecotoxicol. Environ. Saf. 2022, 241, 113838. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, T.M.; Sibley, P.K.; Mabury, S.A.; Muir, D.G.C.; Solomon, K.R. Laboratory Evaluation of the Toxicity of Perfluorooctane Sulfonate (PFOS) on Selenastrum Capricornutum, Chlorella Vulgaris, Lemna Gibba, Daphnia Magna, and Daphnia Pulicaria. Arch. Environ. Contam. Toxicol. 2003, 44, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; Cureton, P.; Hoke, R.A.; Houde, M.; Kumar, A.; Kurias, J.; Lanno, R.; McCarthy, C.; Newsted, J.; Salice, C.J.; et al. Assessing the Ecological Risks of Per- and Polyfluoroalkyl Substances: Current State-of-the Science and a Proposed Path Forward. Environ. Toxicol. Chem. 2020, 40, 564–605. [Google Scholar] [CrossRef]
- Lithodidae, D.A.; Beder, A.M.; Copeman, L.A.; Eckert, G.L. Crustacean Biology. J. Crustac. Biol. J. 2018, 38, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, S.; Liu, J.; Wang, X.; Chen, H.; Hao, H.; Wang, K. Vital Carbohydrate and Lipid Metabolites in Serum Involved in Energy Metabolism during Pubertal Molt of Mud Crab (Scylla paramamosain). Metabolites 2021, 11, 651. [Google Scholar] [CrossRef]
- Kyun, S.; Ding, N.; Han, D. Perfluoroalkyl Substances and Cognitive Function in Older Adults: Should We Consider Non-Monotonic Dose-Responses and Chronic Kidney Disease? Environ. Res. 2021, 192, 110346. [Google Scholar] [CrossRef]
- Park, S.; Ding, N.; Han, D. Perfluoroalkyl Substances and Cognitive Function: Is the Association Non-Monotonic in Humans? Environ. Epidemiol. 2019, 3, 305. [Google Scholar]
- Lind, L.; Zethelius, B.; Salihovic, S.; Lind, M. Circulating Levels of Perfluoroalkyl Substances and Prevalent Diabetes in the Elderly. Diabetologia 2014, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Tarapore, P.; Ouyang, B. Perfluoroalkyl Chemicals and Male Reproductive Health: Do PFOA and PFOS Increase Risk for Male Infertility? Int. J. Environ. Res. Public Health 2021, 18, 3794. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, M.N.; Nagato, E.G.; Lankadurai, B.P.; Simpson, A.J.; Simpson, M.J. Analysis of Sub-Lethal Toxicity of Perfluorooctane Sulfonate (PFOS) to Daphnia Magna Using1H Nuclear Magnetic Resonance-Based Metabolomics. Metabolites 2017, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.D.; Bräunig, J.; Mueller, J.F.; Crompton, M.; Dunstan, R.H.; Nilsson, S. Metabolomic Profiles Associated with Exposure to Per- And Polyfluoroalkyl Substances (PFASs) in Aquatic Environments. Environ. Sci. Process. Impacts 2019, 21, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, L.; Bundschuh, M. Fate and Effects of Poly- and Perfluoroalkyl Substances in the Aquatic Environment: A Review. Environ. Toxicol. Chem. 2014, 33, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Ghisi, R.; Vamerali, T.; Manzetti, S. Accumulation of Perfluorinated Alkyl Substances (PFAS) in Agricultural Plants: A Review. Environ. Res. 2019, 169, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Palmer, K.; Bangma, J.T.; Reiner, J.L.; Bonde, R.K.; Korte, J.E.; Boggs, A.S.P.; Bowden, J.A. Per- and Polyfluoroalkyl Substances (PFAS) in Plasma of the West Indian Manatee (Trichechus manatus). Mar. Pollut. Bull. 2019, 140, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Zhang, H.; Li, L.; Hu, X.; Liu, Y.; Shan, X.; Zhang, S. Bioavailability of Perfluorooctane Sulfonate (PFOS) and Perfluorooctanoic Acid (PFOA) in Biosolids-Amended Soils to Earthworms (Eisenia fetida). Chemosphere 2015, 118, 361–366. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.; Kappleman, W.; DiGuiseppi, W. Ecological Considerations of Per- and Polyfluoroalkyl Substances (PFAS). Curr. Pollut. Rep. 2017, 3, 289–301. [Google Scholar] [CrossRef]
- Wang, W.-X. Bioaccumulation and Biomonitoring. In Marine Ecotoxicology; Academic Press: Cambridge, MA, USA, 2016; pp. 99–119. [Google Scholar] [CrossRef]
- Karnjanapiboonwong, A.; Deb, S.K.; Subbiah, S.; Wang, D.; Anderson, T.A. Perfluoroalkylsulfonic and Carboxylic Acids in Earthworms (Eisenia fetida): Accumulation and Effects Results from Spiked Soils at PFAS Concentrations Bracketing Environmental Relevance. Chemosphere 2018, 199, 168–173. [Google Scholar] [CrossRef]
- Chiesa, L.M.; Nobile, M.; Pasquale, E.; Balzaretti, C.; Cagnardi, P.; Tedesco, D.; Panseri, S.; Arioli, F. Detection of Perfluoroalkyl Acids and Sulphonates in Italian Eel Samples by HPLC-HRMS Orbitrap. Chemosphere 2018, 193, 358–364. [Google Scholar] [CrossRef]
- D’eon, J.C.; Mabury, S.A. Exploring Indirect Sources of Human Exposure to Perfluoroalkyl Carboxylates (PFCAs): Evaluating Uptake, Elimination, and Biotransformation of Polyfluoroalkyl Phosphate Esters (PAPs) in the Rat. Environ. Health Perspect. 2011, 119, 344–350. [Google Scholar] [CrossRef]
- Land, M.; De Wit, C.A.; Bignert, A.; Cousins, I.T.; Herzke, D.; Johansson, J.H.; Martin, J.W. What Is the Effect of Phasing out Long-Chain per- and Polyfluoroalkyl Substances on the Concentrations of Perfluoroalkyl Acids and Their Precursors in the Environment? A Systematic Review. Environ. Evid. 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Whitacre, D.M. Reviews of Environmental Contamination and Toxicology. Rev. Environ. Contam. Toxicol. 2015, 233, 107–130. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, L.; Liu, L.; Liu, Z.; Zhang, Y. Bioaccumulation of Perfluoroalkyl Carboxylates (PFCAs) and Perfluoroalkane Sulfonates (PFSAs) by Earthworms (Eisenia fetida) in Soil. Environ. Pollut. 2013, 179, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Gomis, M.I.; Vestergren, R.; Borg, D.; Cousins, I.T. Comparing the Toxic Potency in Vivo of Long-Chain Perfluoroalkyl Acids and Fluorinated Alternatives. Environ. Int. 2018, 113, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Menger, F.; Pohl, J.; Ahrens, L.; Carlsson, G.; Örn, S. Behavioural Effects and Bioconcentration of Per- and Polyfluoroalkyl Substances (PFASs) in Zebrafish (Danio rerio) Embryos. Chemosphere 2020, 245, 125573. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Low | Low–Medium | Medium | Medium–High | High |
|---|---|---|---|---|---|
| Control | 0 | 0 | 0 | 0 | 0 |
| PFOS | 0.04 | 0.20 | 1.00 | 5.00 | 25.00 |
| GenX | 0.03 | 0.16 | 0.80 | 4.00 | 20.00 |
| PFHxS | 0.03 | 0.15 | 0.76 | 3.78 | 18.92 |
| Metabolite | Mean Square Residual | Comparison between Treatment and Control | ||||
|---|---|---|---|---|---|---|
| LOW-CTRL | L–M–CTRL | MED–CTRL | M–H–CTRL | HIGH–CTRL | ||
| Monostearin | 3 × 1011 | 0.09 | 0.02 | 1.00 | 0.01 | 0.85 |
| Ketobutyric acid | 2 × 1011 | 0.00 | 1.00 | 0.70 | 0.96 | 0.41 |
| Palmitic acid | 3 × 1011 | 0.17 | 1.00 | 1.00 | 0.08 | 0.00 |
| Glycerol | 1 × 1013 | 0.02 | 0.01 | 0.01 | 0.01 | 0.93 |
| Propanoic acid | 2 × 1011 | 0.74 | 0.95 | 0.99 | 0.52 | 0.50 |
| Acetic acid | 2 × 1014 | 1.00 | 0.11 | 1.00 | 0.99 | 0.94 |
| Valine | 1 × 1013 | 0.20 | 0.06 | 0.72 | 0.12 | 0.08 |
| Metabolite | Mean Square Residual | Comparison between Treatment and Control | ||||
|---|---|---|---|---|---|---|
| LOW-CTRL | L–M–CTRL | MED–CTRL | M–H–CTRL | HIGH–CTRL | ||
| Valine | 9 × 1010 | 0.00 | 1.00 | 0.75 | 0.75 | 0.56 |
| Glycerol | 9 × 1010 | 0.01 | 0.04 | 0.00 | 0.01 | 0.00 |
| Monostearin | 9 × 1010 | 0.44 | 0.74 | 0.03 | 0.87 | 0.70 |
| Propanopic acid | 9 × 1010 | 0.01 | 0.02 | 1.00 | 0.04 | 0.02 |
| Cholesterol | 9 × 1010 | 0.27 | 0.44 | 0.00 | 0.23 | 0.10 |
| Palmitic acid | 9 × 1010 | 0.65 | 0.48 | 0.19 | 0.12 | 0.02 |
| Metabolite | Mean Square Residual | Comparison between Treatment and Control | ||||
|---|---|---|---|---|---|---|
| LOW-CTRL | L–M-CTRL | MED-CTRL | M–H-CTRL | HIGH-CTRL | ||
| Propanoic acid | 0.10 | 0.86 | 0.04 | 1.00 | 1.00 | 0.00 |
| Monostearin | 0.43 | 1.00 | 1.00 | 0.01 | 0.99 | 0.00 |
| Cholesterol | 0.48 | 0.03 | 0.99 | 1.00 | 1.00 | 0.01 |
| Proline | 0.31 | 0.02 | 0.02 | 0.10 | 0.00 | 0.25 |
| Treatment | Day 0 (Nominal) | Day 7 (Measured) |
|---|---|---|
| PFOS | 25.00 | 0.11 |
| GenX | 20.00 | 19.71 |
| PFHxS | 18.92 | 12.3 |
| Treatment | High R1 | High R2 | High R3 | High R4 | High R5 |
|---|---|---|---|---|---|
| PFOS | 18.18 | 27.27 | 190.91 | 118.18 | 9.09 |
| GenX | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| PFHxS | 0.05 | 0.07 | 0.04 | 0.08 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinclair, G.M.; Long, S.M.; Singh, N.; Coggan, T.L.; Askeland, M.P.J.; Jones, O.A.H. Exposure to Environmentally Relevant Levels of PFAS Causes Metabolic Changes in the Freshwater Amphipod Austrochiltonia subtenuis. Metabolites 2022, 12, 1135. https://doi.org/10.3390/metabo12111135
Sinclair GM, Long SM, Singh N, Coggan TL, Askeland MPJ, Jones OAH. Exposure to Environmentally Relevant Levels of PFAS Causes Metabolic Changes in the Freshwater Amphipod Austrochiltonia subtenuis. Metabolites. 2022; 12(11):1135. https://doi.org/10.3390/metabo12111135
Chicago/Turabian StyleSinclair, Georgia M., Sara M. Long, Navneet Singh, Timothy L. Coggan, Matthew P. J. Askeland, and Oliver A. H. Jones. 2022. "Exposure to Environmentally Relevant Levels of PFAS Causes Metabolic Changes in the Freshwater Amphipod Austrochiltonia subtenuis" Metabolites 12, no. 11: 1135. https://doi.org/10.3390/metabo12111135
APA StyleSinclair, G. M., Long, S. M., Singh, N., Coggan, T. L., Askeland, M. P. J., & Jones, O. A. H. (2022). Exposure to Environmentally Relevant Levels of PFAS Causes Metabolic Changes in the Freshwater Amphipod Austrochiltonia subtenuis. Metabolites, 12(11), 1135. https://doi.org/10.3390/metabo12111135