
Mining Small Molecules from Teredinibacter turnerae Strains Isolated from Philippine Teredinidae

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Shipworm Collection, Symbionts Isolation, and Processing
2.2. Fractionation of the Crude Methanol Extracts
2.3. LC-MS Profiling and Dereplication of T. turnerae Strains
2.4. Data Processing
3. Results and Discussion
3.1. Dereplication Based on Previous Literature Studies on T. turnerae
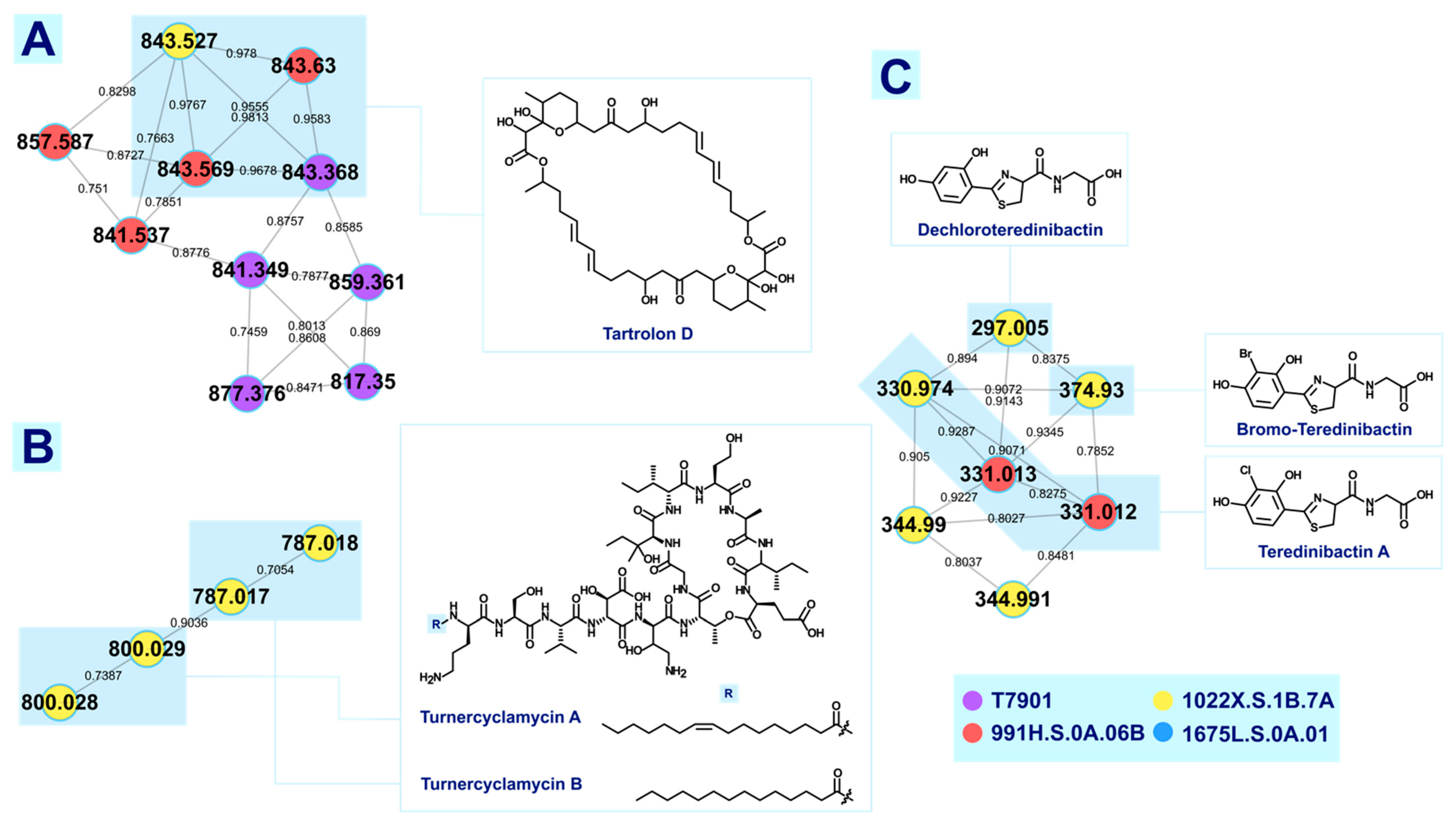
3.1.1. Tartrolons
3.1.2. Turnercyclamycins
3.1.3. Oxylipins
3.1.4. Teredinibactins
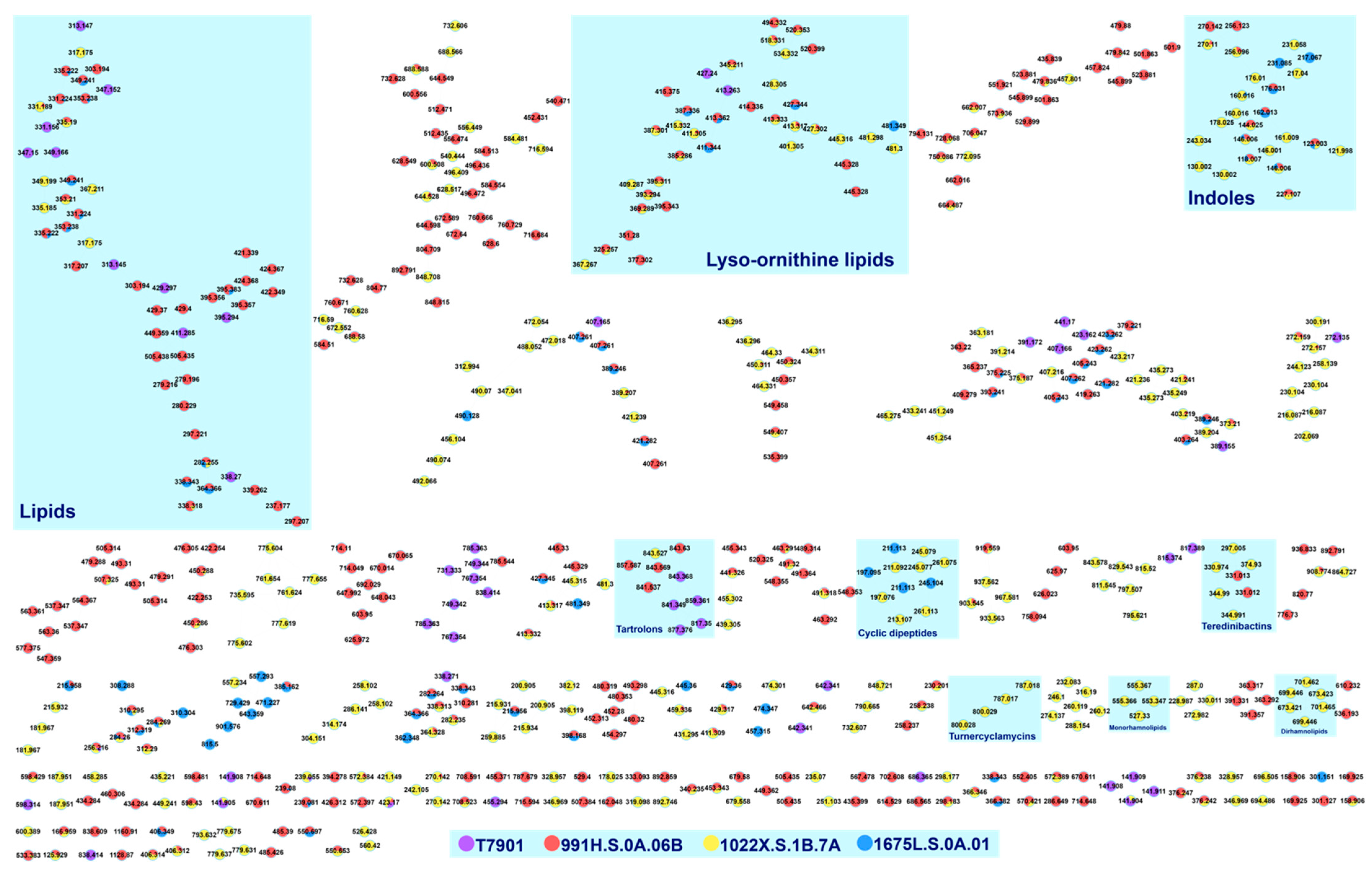
3.2. Dereplication Based on GNPS Molecular Networking
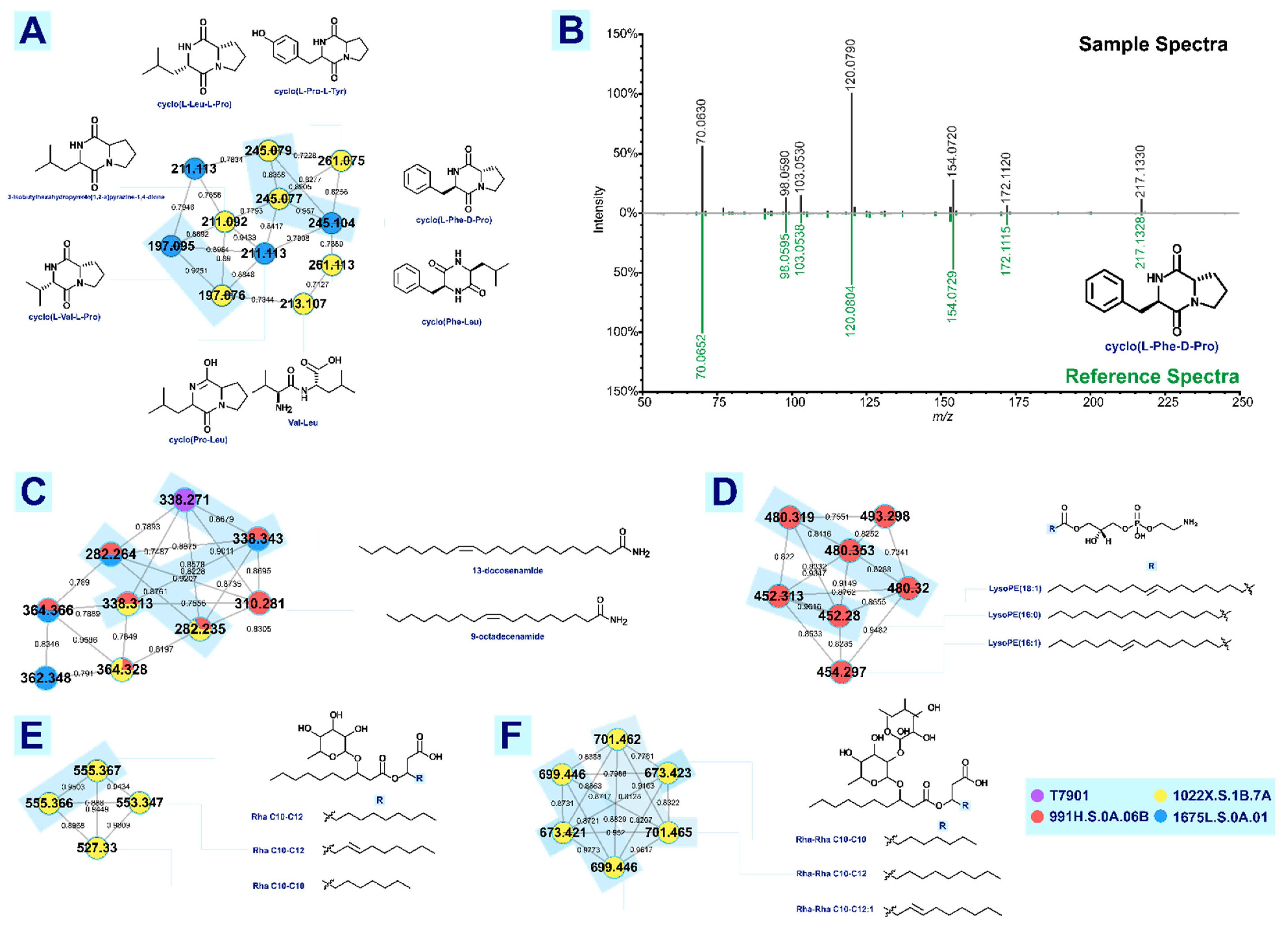
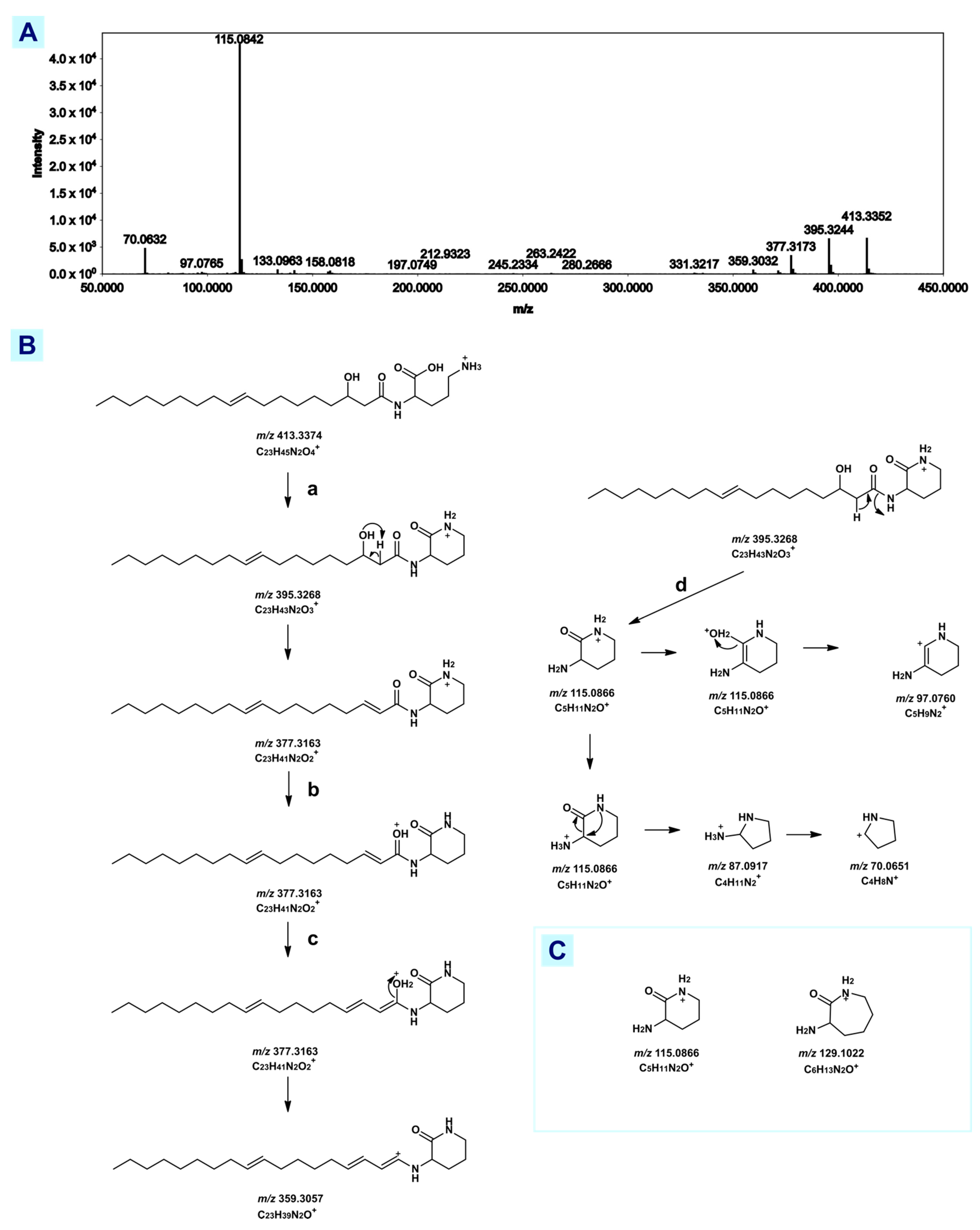
3.2.1. Fatty Acid Amide
3.2.2. Cyclic Dipeptides
3.2.3. Lysophosphatidylethanolamine
3.2.4. Rhamnolipids
3.3. Dereplication Based on Manual Dissociation Analysis
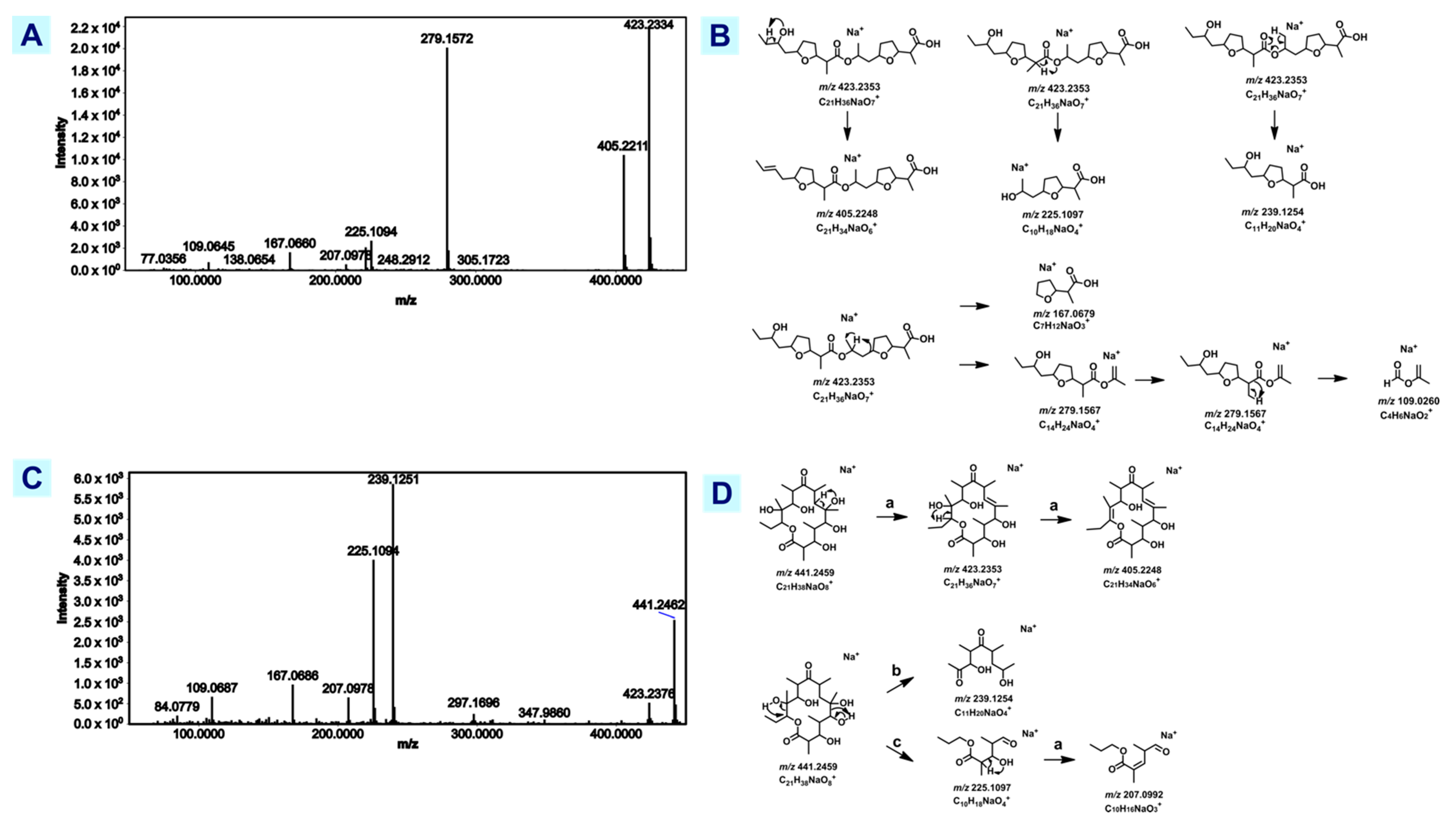
3.3.1. Antibiotic Precursors Erythronolide A and Nonactyl Homononactoate
3.3.2. Lyso-Ornithine Lipids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LC-MS | Liquid Chromatography-Mass Spectrometry |
| UHPLC-HRMS-QTOF | Ultrahigh-performance liquid chromatography with high-resolution mass spectrometry quadrupole time-of-flight |
| GNPS | Global Natural Products Social molecular networking |
| DNP | Dictionary of Natural Products® |
| CMNPD | Comprehensive Marine Natural Products Database |
| LPEs | lysophosphatidylethanolamine |
| CDPs | cyclic dipeptides |
| QS | quorum sensing |
| ESI | Electrospray Ionization |
| MALDI | Matrix-assisted laser desorption/ionization |
| RLs | Rhamnolipids |
| PKS | Polyketide synthase |
| LOLs | Lyso-ornithine lipids |
| SBM | Shipworm basal medium |
| OCF | Open column fraction |
| CID | Collision-induced dissociation |
| RP-HPLC | Reversed-phase High Performance Liquid Chromatography |
| MeOH | Methanol |
| H2O | Water |
| CH3CN | Acetonitrile |
| BGC | Biosynthetic Gene Cluster |
References
- Paknia, O.; Rajaei, S.; Koch, A. Lack of Well-maintained Natural History Collections and Taxonomists in Megadiverse Developing Countries Hampers Global Biodiversity Exploration. Org. Divers. Evol. 2015, 15, 619–629. [Google Scholar] [CrossRef]
- Distel, D.L.; Altamia, M.A.; Lin, Z.; Shipway, J.R.; Han, A.; Forteza, I.; Antemano, R.; Limbaco, M.G.J.P.; Tebo, A.G.; Dechavez, R.; et al. Discovery of Chemoautotrophic Symbiosis in the Giant Shipworm Kuphus polythalamia (Bivalvia: Teredinidae) extends wooden-steps theory. Proc. Natl. Acad. Sci. USA 2017, 114, E3652–E3658. [Google Scholar] [CrossRef] [Green Version]
- Waterbury, J.B.; Calloway, C.B.; Turner, R.D. A Cellulolytic Nitrogen-Fixing Bacterium Cultured from the Gland of Deshayes in Shipworms (Bivalvia: Teredinidae). Science 1983, 221, 1401–1403. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Madupu, R.; Durkin, A.S.; Ekborg, N.A.; Pedamallu, C.S.; Hostetler, J.B.; Radune, D.; Toms, B.S.; Henrissat, B.; Coutinho, P.M.; et al. The Complete Genome of Teredinibacter turnerae T7901: An Intracellular Endosymbiont of Marine Wood-Boring Bivalves (Shipworms). PLoS ONE 2009, 4, e6085. [Google Scholar] [CrossRef]
- Altamia, M.A.; Lin, Z.; Trindade-Silva, A.E.; Uy, I.D.; Shipway, J.R.; Wilke, D.V.; Concepcion, G.P.; Distel, D.L.; Schmidt, E.W.; Haygood, M.G. Secondary Metabolism in the Gill Microbiota of Shipworms (Teredinidae) as Revealed by Comparison of Metagenomes and Nearly Complete Symbiont Genomes. MSystems 2020, 5, e00261-20. [Google Scholar] [CrossRef] [PubMed]
- Han, A.W.; Sandy, M.; Fishman, B.; Trindade-Silva, A.E.; Soares, C.A.G.; Distel, D.L.; Butler, A.; Haygood, M.G. Turnerbactin, a Novel Triscatecholate Siderophore from the Shipworm Endosymbiont Teredinibacter turnerae T7901. PLoS ONE 2013, 8, e76151. [Google Scholar] [CrossRef] [Green Version]
- Elshahawi, S.I.; Trindade-Silva, A.E.; Hanora, A.; Han, A.W.; Flores, M.S.; Vizzoni, V.; Schrago, C.G.; Soares, C.A.; Concepcion, G.P.; Distel, D.L.; et al. Boronated Tartrolon Antibiotic Produced by Symbiotic Cellulose-Degrading Bacteria in Shipworm Gills. Proc. Natl. Acad. Sci. USA 2013, 110, E295–E304. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.W.; Lim, A.L.; Lin, Z.; Bailey, J.; Aoyagi, K.L.; Fisher, M.A.; Barrows, L.R.; Manoil, C.; Schmidt, E.W.; Haygood, M.G. Shipworm Symbiosis Ecology-Guided Discovery of an Antibiotic that Kills Colistin-Resistant Acinetobacter. Cell Chem. Biol. 2021, 28, 1628–1637.e4. [Google Scholar] [CrossRef]
- Miller, B.W.; Schmidt, E.W.; Concepcion, G.P.; Haygood, M.G. Halogenated Metal-Binding Compounds from Shipworm Symbionts. J. Nat. Prod. 2022, 85, 479–484. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete Genome Sequence of the Model Actinomycete Streptomyces coelicolor A3. Nature 2002, 417, 6885. [Google Scholar] [CrossRef]
- Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Shinose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Ōmura, S. Complete Genome Sequence and Comparative Analysis of the Industrial Microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.-L.; Marti, G.; Queiroz, E.F. Advances in Techniques for Profiling Crude Extracts and for the Rapid Identificationof Natural Products: Dereplication, Quality Control and Metabolomics. Curr. Org. Chem. 2010, 14, 1808–1832. [Google Scholar] [CrossRef]
- Yuliana, N.D.; Khatib, A.; Choi, Y.H.; Verpoorte, R. Metabolomics for Bioactivity Assessment of Natural Products. Phytotherapy Res. 2011, 25, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Robes, J.M.D.; Altamia, M.A.; Murdock, E.G.; Concepcion, G.P.; Haygood, M.G.; Puri, A.W. A Conserved Biosynthetic Gene Cluster Is Regulated by Quorum Sensing in a Shipworm Symbiont. Appl. Environ. Microbiol. 2022, 88, e0027022. [Google Scholar] [CrossRef] [PubMed]
- Distel, D.L.; Morrill, W.; MacLaren-Toussaint, N.; Franks, D.; Waterbury, J. Teredinibacter turnerae gen. nov., sp. nov., a Dinitrogen-Fixing, Cellulolytic, Endosymbiotic Gamma-Proteobacterium Isolated from the Gills of Wood-Boring Molluscs (Bivalvia: Teredinidae). Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 6, 2261–2269. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 8. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible Molecular Networking of Untargeted Mass Spectrometry Data Using GNPS. Nat. Protoc. 2020, 15, 6. [Google Scholar] [CrossRef]
- Huber, F.; Ridder, L.; Verhoeven, S.; Spaaks, J.H.; Diblen, F.; Rogers, S.; van der Hooft, J.J.J. Spec2Vec: Improved Mass Spectral Similarity Scoring Through Learning of Structural Relationships. PLOS Comput. Biol. 2021, 17, e1008724. [Google Scholar] [CrossRef]
- Lyu, C.; Chen, T.; Qiang, B.; Liu, N.; Wang, H.; Zhang, L.; Liu, Z. CMNPD: A Comprehensive Marine Natural Products Database Towards Facilitating Drug Discovery from the Ocean. Nucleic Acids Res. 2021, 49, D509–D515. [Google Scholar] [CrossRef]
- Pérez, M.; Crespo, C.; Schleissner, C.; Rodríguez, P.; Zúñiga, P.; Reyes, F. Tartrolon D, a Cytotoxic Macrodiolide from the Marine-Derived Actinomycete Streptomyces sp. MDG-04-17-069. J. Nat. Prod. 2009, 72, 2192–2194. [Google Scholar] [CrossRef] [PubMed]
- Demarque, D.P.; Crotti, A.E.M.; Vessecchi, R.; Lopes, J.L.C.; Lopes, N.P. Fragmentation Reactions Using Electrospray Ionization Mass Spectrometry: An Important Tool for the Structural Elucidation and Characterization of Synthetic and Natural Products. Nat. Prod. Rep. 2016, 33, 432–455. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.; Bandeira, N.; Shen, Z.; Tanner, S.; Briggs, S.; Smith, R.; Pevzner, P. Clustering Millions of Tandem Mass Spectra. J. Proteome Res. 2007, 7, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Beer, I.; Barnea, E.; Ziv, T.; Admon, A. Improving Large-Scale Proteomics by Clustering of Mass Spectrometry Data. PROTEOMICS 2004, 4, 950–960. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.W.; Lim, A.L.; Haygood, M.G.; Schmidt, E.W. Initial Efficacy Determination and Resistance Profile of Anti-Acinetobacter antibiotics, turnercyclamycins. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lacerna, N.M.; Ramones, C.M.V.; Robes, J.M.D.; Picart, M.R.D.; Tun, J.O.; Miller, B.W.; Haygood, M.G.; Schmidt, E.W.; Salvador-Reyes, L.A.; Concepcion, G.P. Inhibition of Biofilm Formation by Modified Oxylipins from the Shipworm Symbiont Teredinibacter turnerae. Mar. Drugs 2020, 18, 656. [Google Scholar] [CrossRef]
- Divito, E.B.; Davic, A.P.; Johnson, M.E.; Cascio, M. Electrospray Ionization and Collision Induced Dissociation Mass Spectrometry of Primary Fatty Acid Amides. Anal. Chem. 2012, 84, 2388–2394. [Google Scholar] [CrossRef]
- Bergé, J.-P.; Barnathan, G. Fatty acids from Lipids of Marine Organisms: Molecular Biodiversity, Roles as Biomarkers, Biologically Active Compounds, and Economical Aspects. Adv. Biochem. Eng./Biotechnol. 2005, 96, 49–125. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, K.; Masaki, T.; Itoh, F.; Kondo, K.; Sudo, K. Isolation of Fatty Acid Amide as an Angiogenic Principle from Bovine Mesentery. Biochem. Biophys. Res. Commun. 1990, 168, 423–429. [Google Scholar] [CrossRef]
- Sun, L.; Lu, Y.; Kronzucker, H.J.; Shi, W. Quantification and Enzyme Targets of Fatty Acid Amides from Duckweed Root Exudates Involved in the Stimulation of Denitrification. J. Plant Physiol. 2016, 198, 81–88. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, Y.; Nakai, S.; Hosomi, M.; Zhang, H.; Kronzucker, H.J.; Shi, W. Stimulation of Nitrogen Removal in the Rhizosphere of Aquatic Duckweed by Root Exudate Components. Planta 2014, 239, 591–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamilmani, E.; Radhakrishnan, R.; Sankaran, K. 13-Docosenamide Release by Bacteria in Response to Glucose During Growth—Fluorescein Quenching and Clinical Application. Appl. Microbiol. Biotechnol. 2018, 102, 6673–6685. [Google Scholar] [CrossRef]
- Cravatt, B.F.; Lerner, R.A.; Boger, D.L. Structure Determination of an Endogenous Sleep-Inducing Lipid, cis-9-Octadecenamide (Oleamide): A Synthetic Approach to the Chemical Analysis of Trace Quantities of a Natural Product. J. Am. Chem. Soc. 1996, 118, 580–590. [Google Scholar] [CrossRef]
- Selvin, J.; Shanmughapriya, S.; Gandhimathi, R.; Seghal Kiran, G.; Rajeetha Ravji, T.; Natarajaseenivasan, K.; Hema, T.A. Optimization and Production of Novel Antimicrobial Agents from Sponge Associated Marine Actinomycetes Nocardiopsis dassonvillei MAD08. Appl. Microbiol. Biotechnol. 2009, 83, 435–445. [Google Scholar] [CrossRef] [PubMed]
- El-Kassas, H.Y.; El-Sheekh, M.M. Induction of the Synthesis of Bioactive Compounds of the Marine Alga Tetraselmis tetrathele (West) Butcher Grown Under Salinity Stress. Egypt. J. Aquat. Res. 2016, 42, 385–391. [Google Scholar] [CrossRef]
- Furtado, N.A.J.C.; Vessecchi, R.; Tomaz, J.C.; Galembeck, S.E.; Bastos, J.K.; Lopes, N.P.; Crotti, A.E.M. Fragmentation of Diketopiperazines from Aspergillus fumigatus by Electrospray Ionization Tandem Mass Spectrometry (ESI-MS/MS). J. Mass Spectrom. 2007, 42, 1279–1286. [Google Scholar] [CrossRef]
- Buroni, S.; Scoffone, V.C.; Fumagalli, M.; Makarov, V.; Cagnone, M.; Trespidi, G.; De Rossi, E.; Forneris, F.; Riccardi, G.; Chiarelli, L.R. Investigating the Mechanism of Action of Diketopiperazines Inhibitors of the Burkholderia cenocepacia Quorum Sensing Synthase CepI: A Site-Directed Mutagenesis Study. Front. Pharmacol. 2018, 9, 836. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, Y.; Ge, Y.; Zhu, X.; Pan, J. Regulatory Mechanisms and Promising Applications of Quorum Sensing-Inhibiting Agents in Control of Bacterial Biofilm Formation. Front. Microbiol. 2020, 11, 589640. [Google Scholar] [CrossRef]
- Zhao, K.; Xing, R.; Yan, X. Cyclic Dipeptides: Biological Activities and Self-Assembled Materials. Pept. Sci. 2021, 113, e24202. [Google Scholar] [CrossRef]
- Balachandra, C.; Padhi, D.; Govindaraju, T. Cyclic Dipeptide: A Privileged Molecular Scaffold to Derive Structural Diversity and Functional Utility. ChemMedChem 2021, 16, 2558–2587. [Google Scholar] [CrossRef]
- Kumar, N.; Mohandas, C.; Nambisan, B.; Kumar, D.R.S.; Lankalapalli, R.S. Isolation of Proline-Based Cyclic Dipeptides from Bacillus sp. N strain Associated with Rhabitid Entomopathogenic Nematode and its Antimicrobial Properties. World J. Microbiol. Biotechnol. 2013, 29, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Mishkind, M. Phosphatidylethanolamine—In a Pinch. Trends Cell Biol. 2000, 10, 368. [Google Scholar] [CrossRef]
- Nishina, A.; Kimura, H.; Sekiguchi, A.; Fukumoto, R.; Nakajima, S.; Furukawa, S. Lysophosphatidylethanolamine in Grifola frondosa as a Neurotrophic Activator via Activation of MAPK. J. Lipid Res. 2006, 47, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meylaers, K.; Clynen, E.; Daloze, D.; DeLoof, A.; Schoofs, L. Identification of 1-lysophosphatidylethanolamine (C(16:1)) as an Antimicrobial Compound in the Housefly, Musca domestica. Insect Biochem. Mol. Biol. 2004, 34, 43–49. [Google Scholar] [CrossRef]
- Farag, K.M.; Palta, J.P. Use of Lysophosphatidylethanolamine, a Natural Lipid, to Retard Tomato Leaf and Fruit Senescence. Physiol. Plant. 1993, 87, 515–521. [Google Scholar] [CrossRef]
- Ryu, S.B.; Karlsson, B.H.; Özgen, M.; Palta, J.P. Inhibition of Phospholipase D by Lysophosphatidylethanolamine, a Lipid-Derived Senescence Retardant. Proc. Natl. Acad. Sci. USA 1997, 94, 12717–12721. [Google Scholar] [CrossRef] [Green Version]
- Watrous, J.D.; Phelan, V.V.; Hsu, C.-C.; Moree, W.J.; Duggan, B.M.; Alexandrov, T.; Dorrestein, P.C. Microbial Metabolic Exchange in 3D. ISME J. 2013, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of Structures, Microbial Origins and Roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, L.; Nitschke, M. Antimicrobial Activity of Rhamnolipids Against Listeria monocytogenes and Their Synergistic Interaction with Nisin. Food Control 2013, 29, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Chong, H.; Li, Q. Microbial Production of Rhamnolipids: Opportunities, Challenges and Strategies. Microb. Cell Factories 2017, 16, 137. [Google Scholar] [CrossRef]
- Roddis, M.; Gates, P.; Roddis, Y.; Staunton, J. Structural Elucidation Studies on 14- and 16-Membered Macrolide Aglycones by Accurate-Mass Electrospray Sequential Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2002, 13, 862–874. [Google Scholar] [CrossRef] [Green Version]
- Ashley, G.W.; Carney, J.R. API-Mass Spectrometry of Polyketides. II. Fragmentation Analysis of 6-Deoxyerythronolide B Analogs. J. Antibiot. 2004, 57, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, T.L.; Campos, A.B.; von Meijenfeldt, F.A.B.; Daniel, J.P.; Ribeiro, G.B.; Silva, G.G.Z.; Wilke, D.V.; de Moraes, D.T.; Dutilh, B.E.; Meirelles, P.M.; et al. The Gill-Associated Microbiome is the Main Source of Wood Plant Polysaccharide Hydrolases and Secondary Metabolite Gene Clusters in the Mangrove Shipworm Neoteredo reynei. PLoS ONE 2018, 13, e0200437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crevelin, E.J.; Crotti, A.E.M.; Zucchi, T.D.; Melo, I.S.; Moraes, L.A.B. Dereplication of Streptomyces sp. AMC 23 Polyether Ionophore Antibiotics by Accurate-Mass Electrospray Tandem Mass Spectrometry. J. Mass Spectrom. 2014, 49, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Q.; Sattler, I.; Thiericke, R.; Grabley, S.; Feng, X.-Z. Feigrisolides A, B, C and D, New Lactones with Antibacterial Activities from Streptomyces griseus. J. Antibiot. 2000, 53, 934–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Sánchez-Andrea, I.; Villanueva, L.; Wienk, H.; Schoutsen, F.; Stams, A.J.M.; Sinninghe Damsté, J.S. Lysine and Novel Hydroxylysine Lipids in Soil Bacteria: Amino Acid Membrane Lipid Response to Temperature and pH in Pseudopedobacter saltans. Front. Microbiol. 2015, 6, 637. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Chan, W.; Ang, I.L.; Wei, R.; Lam, M.M.T.; Lei, K.M.K.; Poon, T.C.W. Revisiting Fragmentation Reactions of Protonated α-Amino Acids by High-Resolution Electrospray Ionization Tandem Mass Spectrometry with Collision-Induced Dissociation. Sci. Rep. 2019, 9, 6453. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ferguson-Miller, S.M.; Reid, G.E. Characterization of Ornithine and Glutamine Lipids Extracted from Cell Membranes of Rhodobacter sphaeroides. J. Am. Soc. Mass Spectrom. 2009, 20, 198–212. [Google Scholar] [CrossRef] [Green Version]
- Williams, W.; Kunorozva, L.; Klaiber, I.; Henkel, M.; Pfannstiel, J.; Van Zyl, L.J.; Hausmann, R.; Burger, A.; Trindade, M. Novel Metagenome-Derived Ornithine Lipids Identified by Functional Screening for Biosurfactants. Appl. Microbiol. Biotechnol. 2019, 103, 4429–4441. [Google Scholar] [CrossRef]
- Jawad, J. Isolation, Structure Elucidation and Bioactivity Profiling of Lyso-Ornithine Lipids from the Marine Bacterium Lacinutrix sp. Master’s Thesis, UiT Norges Arktiske Universitet, Tromsø, Norway, 2021. [Google Scholar]
- Kristoffersen, V.; Jenssen, M.; Jawad, H.R.; Isaksson, J.; Hansen, E.H.; Rämä, T.; Hansen, K.Ø.; Andersen, J.H. Two Novel Lyso-Ornithine Lipids Isolated from an Arctic Marine Lacinutrix sp. Bacterium. Molecules 2021, 26, 5295. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Sampling Site | Date Collected | Host Organism | Part |
|---|---|---|---|---|
| 991H.S.0A.06B | Brgy Danao, Panglao Bohol Philippines | 4 May 2010 | Lyrodus pedicellatus | gill/ctenidium |
| 1675L.S.0A.01 | Brgy Sta Ana, Kalamansig, Sultan Kudarat Philippines | 24 November 2010 | Kuphus polythamia | gill/ctenidium |
| 1022X.S.1B.7A | Cataban; Camotes Sea, Talibon, Bohol Philippines | 1 May 2010 | Lyrodus massa | caecum |
| T7901 (reference strain) | Newport Estuary, Beaufort North Carolina, USA | 1979 by John Waterbury | Bankia gouldi | gill/ctenidium |
| Compound Name | Major Ion | Monoisotopic Mass | Accurate Mass | Mass Error (ppm) | Cosine Score | Strains |
|---|---|---|---|---|---|---|
| Tartrolon D | [M+Na]+ | 843.450 | 843.451 | 1.19 | MN/FA * | T7901, 991H.S.0A.06B, 1022X.S.1B.7A |
| Turnercyclamycin A | [M+H]+ | 786.958 | 786.955 | 3.18 | MN/FA | 1022X.S.1B.7A |
| Turnercyclamycin B | [M+H]+ | 799.966 | 799.969 | 4.13 | MN/FA | 1022X.S.1B.7A |
| Teredinibactin A | [M+H]+ | 331.016 | 331.016 | 4.23 | MN/FA | 991H.S.0A.06B, 1022X.S.1B.7A |
| Dechloroteredinibactin | [M+H]+ | 297.055 | 297.055 | 3.37 | MN/FA | 1022X.S.1B.7A |
| Brominated teredinibactin | [M+H]+ | 374.964 | 374.964 | 0.80 | MN/FA | 1022X.S.1B.7A |
| Teredinibactin A + (CH3) | [M+H]+ | 345.031 | 345.032 | 4.06 | MN/FA | 1022X.S.1B.7A |
| Turneroic Acid | [M+Na]+ | 337.235 | 337.236 | 1.48 | MN/FA | 991H.S.0A.06B |
| 11-oxooctadec-12-enoic acid | [M+H-H2O]+ | 279.232 | 279.233 | 2.86 | MN/FA | 991H.S.0A.06B |
| (13Z)-docosenamide | [M+H]+ | 338.342 | 338.342 | 0.30 | 0.81 | T7901, 991H.S.0A.06B, 1022X.S.1B.7A, 1675L.S.0A.01 |
| (9Z)-octadecenamide | [M+H]+ | 282.279 | 282.278 | 3.90 | 0.87 | 991H.S.0A.06B, 1022X.S.1B.7A, 1675L.S.0A.01 |
| cyclo(L-Val-L-Pro) | [M+H]+ | 197.128 | 197.129 | 5.07 | 0.88 | 1022X.S.1B.7A, 1675L.S.0A.01 |
| cyclo(L-Phe-D-Pro) | [M+H]+ | 245.128 | 245.128 | 3.26 | 0.93 | 1022X.S.1B.7A, 1675L.S.0A.01 |
| cyclo(Pro-Leu) | [M+H]+ | 211.144 | 211.145 | 2.84 | 0.87 | 1675L.S.0A.01 |
| 3-(2-methylpropyl)-2,3,6,7,8,8a-hexahydropyrrolo[1,2-a]pyrazine-1,4-dione | [M+H]+ | 211.144 | 211.145 | 2.84 | 0.83 | 1022X.S.1B.7A |
| Val-Leu | [M+H-H2O]+ | 213.156 | 213.159 | 1.88 | 0.86 | 1022X.S.1B.7A |
| 1-palmitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamine | [M+H]+ | 454.293 | 454.294 | 3.30 | 0.94 | 991H.S.0A.06B |
| 1-(9Z-octadecenoyl)-sn-glycero-3-phosphoethanolamine | [M+H]+ | 480.309 | 480.308 | 1.67 | 0.88 | 991H.S.0A.06B |
| LysoPE (16:1) | [M+H]+ | 452.277 | 452.277 | 0.44 | MN/FA | 991H.S.0A.06B |
| Rha Rha C10-C10 | [M+Na]+ | 673.377 | 673.377 | 0.74 | 0.87 | 1022X.S.1B.7A |
| Rha Rha C10-C12:1 | [M+Na]+ | 699.393 | 699.395 | 3.29 | MN/FA | 1022X.S.1B.7A |
| Rha Rha C10-C12 | [M+Na]+ | 701.408 | 701.409 | 1.28 | 0.75 | 1022X.S.1B.7A |
| Rha C10-C10 | [M+Na]+ | 527.319 | 527.318 | 2.84 | MN/FA | 1022X.S.1B.7A |
| Rha C10-C12:1 | [M+Na]+ | 553.335 | 553.336 | 2.17 | MN/FA | 1022X.S.1B.7A |
| Rha C10-C12 | [M+Na]+ | 555.350 | 555.349 | 1.62 | MN/FA | 1022X.S.1B.7A |
| Erythronolide A | [M+Na]+ | 441.246 | 441.246 | 0.68 | MN/FA | T7901 |
| Nonactyl homononactoate | [M+Na]+ | 423.235 | 423.236 | 0.95 | MN/FA | 991H.S.0A.06B, 1022X.S.1B.7A, 1675L.S.0A.01 |
| C21H42N2O4 | [M+H]+ | 387.322 | 387.323 | 2.58 | MN/FA | 991H.S.0A.06B, 1022X.S.1B.7A, 1675L.S.0A.01 |
| C23H44N2O4 | [M+H]+ | 413.337 | 413.339 | 4.60 | MN/FA | T7901, 991H.S.0A.06B, 1022X.S.1B.7A, 1675L.S.0A.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villacorta, J.B.; Rodriguez, C.V.; Peran, J.E.; Batucan, J.D.; Concepcion, G.P.; Salvador-Reyes, L.A.; Junio, H.A. Mining Small Molecules from Teredinibacter turnerae Strains Isolated from Philippine Teredinidae. Metabolites 2022, 12, 1152. https://doi.org/10.3390/metabo12111152
Villacorta JB, Rodriguez CV, Peran JE, Batucan JD, Concepcion GP, Salvador-Reyes LA, Junio HA. Mining Small Molecules from Teredinibacter turnerae Strains Isolated from Philippine Teredinidae. Metabolites. 2022; 12(11):1152. https://doi.org/10.3390/metabo12111152
Chicago/Turabian StyleVillacorta, Jamaine B., Camille V. Rodriguez, Jacquelyn E. Peran, Jeremiah D. Batucan, Gisela P. Concepcion, Lilibeth A. Salvador-Reyes, and Hiyas A. Junio. 2022. "Mining Small Molecules from Teredinibacter turnerae Strains Isolated from Philippine Teredinidae" Metabolites 12, no. 11: 1152. https://doi.org/10.3390/metabo12111152
APA StyleVillacorta, J. B., Rodriguez, C. V., Peran, J. E., Batucan, J. D., Concepcion, G. P., Salvador-Reyes, L. A., & Junio, H. A. (2022). Mining Small Molecules from Teredinibacter turnerae Strains Isolated from Philippine Teredinidae. Metabolites, 12(11), 1152. https://doi.org/10.3390/metabo12111152