
Iron Metabolism in Normal and Pathological Pregnancies and Fetal Consequences


Abstract
:1. Introduction
2. Systemic Iron Metabolism Outside of Pregnancy
3. Adaptation to Physiological Pregnancy
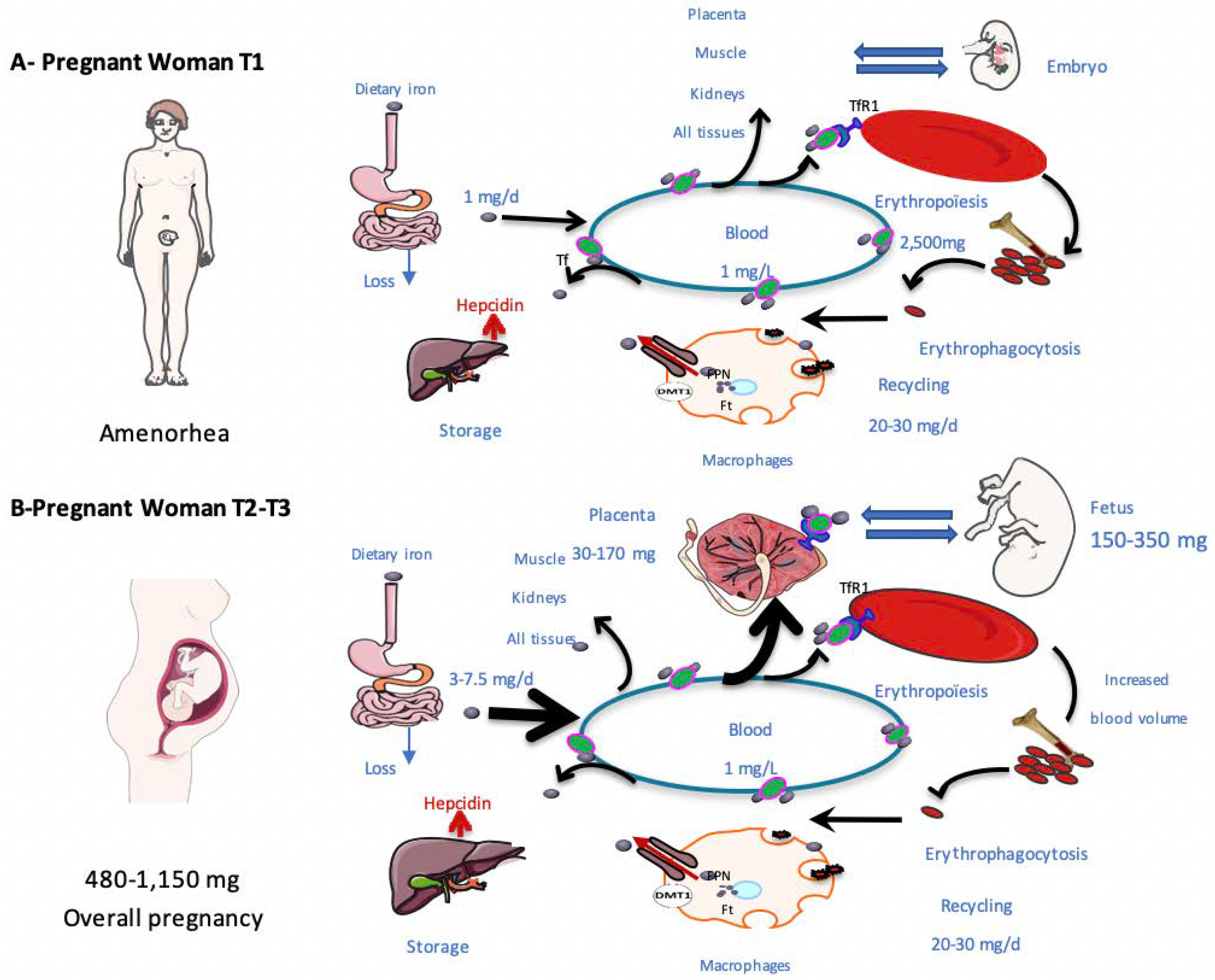
3.1. Iron Needs during Pregnancy
3.2. Adaptation of Iron Metabolism to Pregnancy
3.3. Sources of Placental Iron
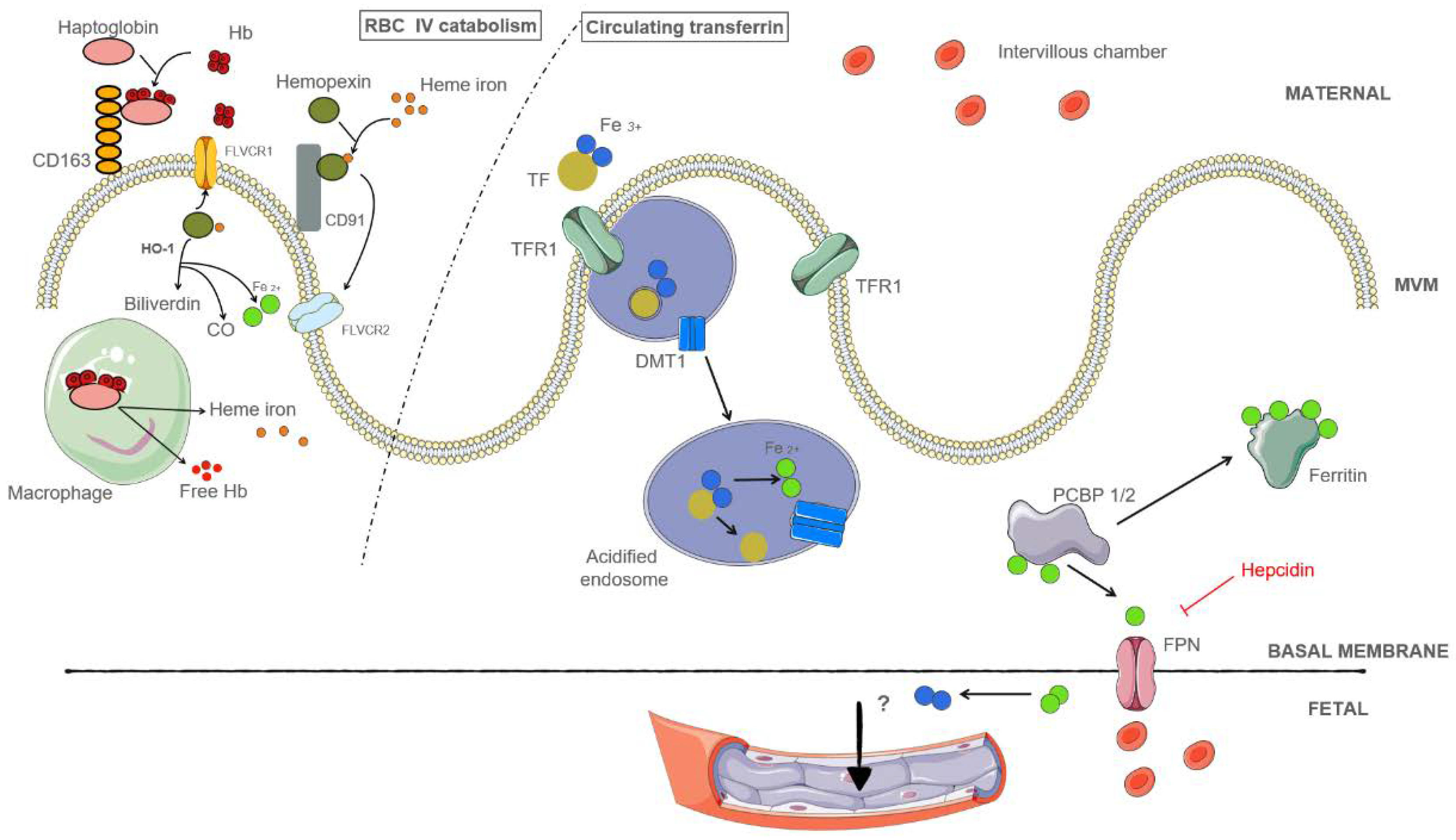
3.3.1. Placental Iron Is Obtained from Holo-Transferrin in the Maternal Circulation
3.3.2. Placental Iron May Be Obtained from Hemolysis
3.4. Regulation of Placental Iron Transport
3.5. The role of Heme Oxygenase during Pregnancy
3.6. Exploration of Iron Metabolism during Pregnancy
4. Iron Metabolism in Pathological Pregnancies
4.1. Preeclampsia
4.1.1. Early-Onset Preeclampsia (<34 WG)
4.1.2. Late-Onset Preeclampsia (≥34 WG)
4.2. The Impact of Toxoplasma and Plasmodium Infections on Iron Metabolism
5. Hemoglobin Diseases and Pregnancy
5.1. Sickle-Cell Disease
5.2. Thalassemia
6. Obesity, Bariatric Surgery, and Pregnancy
6.1. Obesity and Iron Deficiency
6.2. Iron Status after Bariatric Surgery
7. Long-Term Fetal Consequences of Changes in Iron Metabolism and Iron Deficiency
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daher, R.; Karim, Z. Iron metabolism: State of the art. Transfus. Clin. Biol. 2017, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Emerit, J.; Beaumont, C.; Trivin, F. Iron metabolism, free radicals, and oxidative injury. Biomed. Pharmacother. 2001, 55, 333–339. [Google Scholar] [CrossRef]
- Tuck, S.; Jensen, C.; Wonke, B.; Yardumian, A. Pregnancy management and outcomes in women with thalassaemia major. J. Pediatr. Endocrinol. Metab. JPEM 1998, 11 (Suppl. 3), 923–928. [Google Scholar]
- Boga, C.; Ozdogu, H. Pregnancy and sickle cell disease: A review of the current literature. Crit. Rev. Oncol. Hematol. 2016, 98, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Capelletti, M.M.; Manceau, H.; Puy, H.; Peoc’h, K. Ferroptosis in Liver Diseases: An Overview. Int. J. Mol. Sci. 2020, 21, 4908. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.L.; Sangkhae, V.; Balušíková, K.; Palaskas, N.J.; Ganz, T.; Nemeth, E. Iron-dependent apoptosis causes embryotoxicity in inflamed and obese pregnancy. Nat. Commun. 2021, 12, 4026. [Google Scholar] [CrossRef]
- Camaschella, C. Iron-Deficiency Anemia. N. Engl. J. Med. 2015, 372, 1832–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassebaum, N.J.; Jasrasaria, R.; Naghavi, M.; Wulf, S.K.; Johns, N.; Lozano, R.; Regan, M.; Weatherall, D.; Chou, D.P.; Eisele, T.P.; et al. A systematic analysis of global anemia burden from 1990 to 2010. Blood 2014, 123, 615–624. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists. Anemia in Pregnancy: ACOG Practice Bulletin, Number 233. Obstet. Gynecol. 2021, 138, e55–e64. [Google Scholar] [CrossRef] [PubMed]
- Pavord, S.; Myers, B.; Robinson, S.; Allard, S.; Strong, J.; Oppenheimer, C. UK guidelines on the management of iron deficiency in pregnancy. Br. J. Haematol. 2012, 156, 588–600. [Google Scholar] [CrossRef]
- Daru, J.; Zamora, J.; Fernández-Félix, B.M.; Vogel, J.; Oladapo, O.T.; Morisaki, N.; Tunçalp, Ö.; Torloni, M.R.; Mittal, S.; Jayaratne, K.; et al. Risk of maternal mortality in women with severe anaemia during pregnancy and post partum: A multilevel analysis. Lancet Glob. Health 2018, 6, e548–e554. [Google Scholar] [CrossRef] [Green Version]
- Scholl, T.O.; Reilly, T. Anemia, iron and pregnancy outcome. J. Nutr. 2000, 130 (Suppl. 2S), 443S–447S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, R.D.; Greer, F.R.; Committe on Nutrition American Academy of Pediatrics. Clinical report-Diagnosis and prevention of iron deficiency and iron-deficiency anemia in infants and young children (0-3 years of age). Pediatrics 2010, 126, 1040–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieff, M.K. Long-term brain and behavioral consequences of early iron deficiency. Nutr. Rev. 2011, 69 (Suppl. 1), S43–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegersma, A.M.; Dalman, C.; Lee, B.K.; Karlsson, H.; Gardner, R.M. Association of Prenatal Maternal Anemia with Neurodevelopmental Disorders. JAMA Psychiatry 2019, 76, 1294–1304. [Google Scholar] [CrossRef]
- Weinberg, J.; Levine, S.; Dallman, P.R. Long-term consequences of early iron deficiency in the rat. Pharmacol. Biochem. Behav. 1979, 11, 631–638. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J.; Buckett, P.D.; Böhlke, M.; Maher, T.J.; Wessling-Resnick, M. Severe Postnatal Iron Deficiency Alters Emotional Behavior and Dopamine Levels in the Prefrontal Cortex of Young Male Rats. J. Nutr. 2011, 141, 2133–2138. [Google Scholar] [CrossRef] [Green Version]
- Skolmowska, D.; Głąbska, D. Analysis of Heme and Non-Heme Iron Intake and Iron Dietary Sources in Adolescent Menstruating Females in a National Polish Sample. Nutrients 2019, 11, 1049. [Google Scholar] [CrossRef] [Green Version]
- The Nutrition monitoring division Human nutrition information service. Nationwide Food Consumption Survey. Nutr. Today 1986, 21, 31. [Google Scholar] [CrossRef]
- Krause’s Food & the Nutrition Care Process-14th Edition. Available online: https://www.elsevier.com/books/krauses-food-and-the-nutrition-care-process/mahan/978-0-323-34075-5 (accessed on 9 June 2021).
- West, A.R.; Oates, P.S. Mechanisms of heme iron absorption: Current questions and controversies. World J. Gastroenterol. 2008, 14, 4101–4110. [Google Scholar] [CrossRef]
- Olivares, M.; Figueroa, C.; Pizarro, F. Acute Copper and Ascorbic Acid Supplementation Inhibits Non-heme Iron Absorption in Humans. Biol. Trace Elem. Res. 2016, 172, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Hill, M.; Johnson, A.; Latunde-Dada, G.O. Modulation of Dcytb (Cybrd 1) expression and function by iron, dehydroascorbate and Hif-2α in cultured cells. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 106–112. [Google Scholar] [CrossRef] [PubMed]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef] [PubMed]
- Vlachodimitropoulou, E.; Naftalin, R.J.; Sharp, P.A. Quercetin is a substrate for the transmembrane oxidoreductase Dcytb. Free Radic. Biol. Med. 2010, 48, 1366–1369. [Google Scholar] [CrossRef]
- Cao, C.; O’Brien, K.O. Pregnancy and iron homeostasis: An update. Nutr. Rev. 2013, 71, 35–51. [Google Scholar] [CrossRef]
- Winter, W.; Bazydlo, L.; Harris, N. The molecular biology of human iron metabolism. Lab. Med. 2014, 45, 92–102. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Ruwe, T.A.; Vieth, K.R.; Casu, C.; Rivella, S.; Jormakka, M.; et al. Structure-function analysis of ferroportin defines the binding site and an alternative mechanism of action of hepcidin. Blood 2018, 131, 899–910. [Google Scholar] [CrossRef]
- Gkouvatsos, K.; Papanikolaou, G.; Pantopoulos, K. Regulation of iron transport and the role of transferrin. Biochim. Biophys. Acta 2012, 1820, 188–202. [Google Scholar] [CrossRef]
- Shayeghi, M.; Latunde-Dada, G.O.; Oakhill, J.S.; Laftah, A.H.; Takeuchi, K.; Halliday, N.; Khan, Y.; Warley, A.; McCann, F.E.; Hider, R.C.; et al. Identification of an intestinal heme transporter. Cell 2005, 122, 789–801. [Google Scholar] [CrossRef] [Green Version]
- Qiu, A.; Jansen, M.; Sakaris, A.; Min, S.H.; Chattopadhyay, S.; Tsai, E. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell 2006, 127, 917–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopal, A.; Rao, A.U.; Amigo, J.; Tian, M.; Upadhyay, S.K.; Hall, C.; Uhm, S.; Mathew, M.K.; Fleming, M.D.; Paw, B.H.; et al. Haem homeostasis is regulated by the conserved and concerted functions of HRG-1 proteins. Nature 2008, 453, 1127–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Protchenko, O.; Philpott, C.C.; Hamza, I. Topologically conserved residues direct heme transport in HRG-1-related proteins. J. Biol. Chem. 2012, 287, 4914–4924. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ortiz De Montellano, P.R. Reaction intermediates and single turnover rate constants for the oxidation of heme by human heme oxygenase-1. J. Biol. Chem. 2000, 275, 5297–5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulec, S.; Anderson, G.; Collins, J. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G397–G409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A.; Quigley, J.G. Heme and FLVCR-related transporter families SLC48 and SLC49. Mol. Asp. Med. 2013, 34, 669. [Google Scholar] [CrossRef] [Green Version]
- Cacoub, P.; Vandewalle, C.; Peoc’h, K. Using transferrin saturation as a diagnostic criterion for iron deficiency: A systematic review. Crit. Rev. Clin. Lab. Sci. 2019, 56, 526–532. [Google Scholar] [CrossRef]
- Elsayed, M.E.; Sharif, M.U.; Stack, A.G. Transferrin Saturation: A Body Iron Biomarker. Adv. Clin. Chem. 2016, 75, 71–97. [Google Scholar] [CrossRef]
- Gozzelino, R.; Arosio, P. Iron homeostasis in health and disease. Int. J. Mol. Sci. 2016, 17, 130. [Google Scholar] [CrossRef] [Green Version]
- Koenig, M.D.; Tussing-Humphreys, L.; Day, J.; Cadwell, B.; Nemeth, E. Hepcidin and iron homeostasis during pregnancy. Nutrients 2014, 6, 3062–3083. [Google Scholar] [CrossRef]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganz, T. Hepcidin and Its Role in Regulating Systemic Iron Metabolism. Hematol. Am. Soc. Hematol. Am. Soc. Hematol. Educ. Program 2006, 2006, 29–35. [Google Scholar] [CrossRef]
- Zhang, D.L.; Wu, J.; Shah, B.N.; Greutélaers, K.C.; Ghosh, M.C.; Ollivierre, H.; Su, X.Z.; Thuma, P.E.; Bedu-Addo, G.; Mockenhaupt, F.P.; et al. Erythrocytic ferroportin reduces intracellular iron accumulation, hemolysis, and malaria risk. Science 2018, 359, 1520–1523. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, G.; Chauvet, C.; Viatte, L.; Danan, J.L.; Bigard, X.; Devaux, I.; Beaumont, C.; Kahn, A.; Vaulont, S. The gene encoding the iron regulatory peptide hepcidin is regulated by anemia, hypoxia, and inflammation. J. Clin. Investig. 2002, 110, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, G.; Bennoun, M.; Porteu, A.; Mativet, S.; Beaumont, C.; Grandchamp, B.; Sirito, M.; Sawadogo, M.; Kahn, A.; Vaulont, S. Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc. Natl. Acad. Sci. USA 2002, 99, 4596–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosano, E.; Fagoonee, S.; Morello, N.; Vinchi, F.; Fiorito, V. Heme scavenging and the other facets of hemopexin. Antioxid. Redox Signal. 2010, 12, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; McCulloh, R.J. Hemopexin and haptoglobin: Allies against heme toxicity from hemoglobin not contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef]
- Viteri, F.E. The consequences of iron deficiency and anemia in pregnancy. Adv. Exp. Med. Biol. 1994, 352, 127–139. [Google Scholar] [CrossRef]
- Fisher, A.L.; Nemeth, E. Iron homeostasis during pregnancy. Am. J. Clin. Nutr. 2017, 106 (Suppl. 6), 1567S–1574S. [Google Scholar] [CrossRef]
- Georgieff, M.K. Iron Deficiency in Pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 516–524. [Google Scholar] [CrossRef]
- Barrett, J.; Whittaker, P.; Williams, J.; Lind, T. Absorption of non-haem iron from food during normal pregnancy. BMJ (Clin. Res. Ed.) 1994, 309, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milan, N. Iron and pregnancy--a delicate balance. Ann. Hematol. 2006, 85, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Young, M.F.; Griffin, I.; Pressman, E.; McIntyre, A.W.; Cooper, E.; McNanley, T.; Harris, Z.L.; Westerman, M.; O’Brien, K.O. Maternal Hepcidin Is Associated with Placental Transfer of Iron Derived from Dietary Heme and Nonheme Sources. J. Nutr. 2012, 142, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bah, A.; Pasricha, S.R.; Jallow, M.W.; Sise, E.A.; Wegmuller, R.; Armitage, A.E.; Drakesmith, H.; Moore, S.E.; Prentice, A.M. Serum Hepcidin Concentrations Decline during Pregnancy and May Identify Iron Deficiency: Analysis of a Longitudinal Pregnancy Cohort in the Gambia. J. Nutr. 2017, 147, 1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millard, K.; Frazer, D.; Wilkins, S.; Anderson, G. Changes in the expression of intestinal iron transport and hepatic regulatory molecules explain the enhanced iron absorption associated with pregnancy in the rat. Gut 2004, 53, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Van Santen, S.; Kroot, J.J.C.; Zijderveld, G.; Wiegerinck, E.T.; Spaanderman, M.E.A.; Swinkels, D.W. The iron regulatory hormone hepcidin is decreased in pregnancy: A prospective longitudinal study. Clin. Chem. Lab. Med. 2013, 51, 1395–1401. [Google Scholar] [CrossRef]
- Gambling, L.; Danzeisen, R.; Gair, S.; Lea, R.G.; Charania, Z.; Solanky, N.; Joory, K.D.; Srai, S.K.; McArdle, H.J. Effect of iron deficiency on placental transfer of iron and expression of iron transport proteins in vivo and in vitro. Biochem. J. 2001, 356 Pt 3, 883–889. [Google Scholar] [CrossRef]
- Mazgaj, R.; Lipiński, P.; Edison, E.S.; Bednarz, A.; Staroń, R.; Haberkiewicz, O.; Lenartowicz, M.; Smuda, E.; Jonczy, A.; Starzynski, R.R. Marginally reduced maternal hepatic and splenic ferroportin under severe nutritional iron deficiency in pregnancy maintains systemic iron supply. Am. J. Hematol. 2021, 96, 659. [Google Scholar] [CrossRef]
- Kämmerer, L.; Mohammad, G.; Wolna, M.; Robbins, P.A.; Lakhal-Littleton, S. Fetal liver hepcidin secures iron stores in utero. Blood 2020, 136, 1549–1557. [Google Scholar] [CrossRef]
- Sangkhae, V.; Nemeth, E. Placental iron transport: The mechanism and regulatory circuits. Free Radic. Biol. Med. 2019, 133, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, M.; Graversen, J.; Jacobsen, C.; Sonne, O.; Hoffman, H.; Law, S.; Moestrup, S.K. Identification of the haemoglobin scavenger receptor. Nature 2001, 409, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Seligman, P.; Schleicher, R.B.; Allen, R. Isolation and characterization of the transferrin receptor from human placenta. J. Biol. Chem. 1979, 25, 9943–9946. [Google Scholar] [CrossRef]
- Levy, J.E.; Jin, O.; Fujiwara, Y.; Kuo, F.; Andrews, N.C. Transferrin receptor is necessary for development of erythrocytes and the nervous system. Nat. Genet. 1999, 21, 396–399. [Google Scholar] [CrossRef]
- Srai, S.K.S.; Bomford, A.; McArdle, H.J. Iron transport across cell membranes: Molecular understanding of duodenal and placental iron uptake. Best Pract. Res. Clin. Haematol. 2002, 15, 243–259. [Google Scholar] [CrossRef]
- Chong, W.; Kwan, P.; Chan, L.; Chiu, P.; Cheung, T.; Lau, T. Expression of divalent metal transporter 1 (DMT1) isoforms in first trimester human placenta and embryonic tissues. Hum. Reprod. 2005, 20, 3532–3538. [Google Scholar] [CrossRef] [Green Version]
- Venkata Surekha, M.; Sujatha, T.; Gadhiraju, S.; Uday Kumar, P.; Siva Prasad, M.; Sailaja, G.; Bhaskar, V.; Srinivas, T. Expression of iron transport protein Divalent metal transporter 1 (DMT1) increases in response to maternal iron deficiency anemia in near term to term placenta. J. Matern. Fetal Neonatal Med. 2020, 1–9. [Google Scholar] [CrossRef]
- Begum, N.A.; Kobayashi, M.; Moriwaki, Y.; Matsumoto, M.; Toyoshima, K.; Seya, T. Mycobacterium bovis BCG cell wall and lipopolysaccharide induce a novel gene, BIGM103, encoding a 7-TM protein: Identification of a new protein family having Zn-transporter and Zn-metalloprotease signatures. Genomics 2002, 80, 630–645. [Google Scholar] [CrossRef]
- Wang, C.Y.; Jenkitkasemwong, S.; Duarte, S.; Sparkman, B.K.; Shawki, A.; Mackenzie, B.; Knutson, M.D. ZIP8 is an iron and zinc transporter whose cell-surface expression is up-regulated by cellular iron loading. J. Biol. Chem. 2012, 287, 34032–34043. [Google Scholar] [CrossRef] [Green Version]
- Beker Aydemir, T.; Chang, S.M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.S.; Karabiyik, A.; Cousins, R.J. Zinc transporter ZIP14 functions in hepatic zinc, iron and glucose homeostasis during the innate immune response (endotoxemia). PLoS ONE 2012, 7, e48679. [Google Scholar] [CrossRef] [Green Version]
- Pavličev, M.; Wagner, G.; Chavan, A.; Owens, K.; Maziarz, J.; Dunn-Fletcher, C.; Kallapur, S.G.; Muglia, L.; Jones, H. Single-cell transcriptomics of the human placenta: Inferring the cell communication network of the maternal-fetal interface. Genome Res. 2017, 27, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Bastin, J.; Drakesmith, H.; Rees, M.; Sargent, I.; Townsend, A. Localisation of proteins of iron metabolism in the human placenta and liver. Br. J. Haematol. 2006, 134, 532–543. [Google Scholar] [CrossRef]
- Georgieff, M.K.; Wobken, J.K.; Welle, J.; Burdo, J.R.; Connor, J.R. Identification and Localization of Divalent Metal Transporter-1 (DMT-1) in Term Human Placenta. Placenta 2000, 21, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Fleming, M.D. The placenta: The forgotten essential organ of iron transport. Nutr. Rev. 2016, 74, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elad, D.; Levkovitz, R.; Jaffa, A.; Desoye, G.; Hod, M. Have we neglected the role of fetal endothelium in transplacental transport? Traffic 2014, 15, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vashchenko, G.; MacGillivray, R.T.A. Multi-Copper Oxidases and Human Iron Metabolism. Nutrients 2013, 5, 2289–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hvidberg, V.; Maniecki, M.; Jacobsen, C.; Højrup, P.; Møller, H.; Moestrup, S. Identification of the receptor scavenging hemopexin-heme complexes. Blood 2005, 106, 2572–2579. [Google Scholar] [CrossRef]
- Lipiński, P.; Styś, A.; Starzyński, R. Molecular insights into the regulation of iron metabolism during the prenatal and early postnatal periods. Cell. Mol. Life Sci. CMLS 2013, 70, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.; Leibold, E.; Harris, Z.; Wobken, J.; Clarke, S.; Zumbrennen, K.; Eisenstein, R.S.; Georgieff, M.K. Influence of gestational age and fetal iron status on IRP activity and iron transporter protein expression in third-trimester human placenta. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis-an update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef] [Green Version]
- Selezneva, A.I.; Walden, W.E.; Volz, K.W. Nucleotide-specific recognition of iron-responsive elements by iron regulatory protein 1. J. Mol. Biol. 2013, 425, 3301–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peoc’h, K.; Puy, V.; Fournier, T. Haem oxygenases play a pivotal role in placental physiology and pathology. Hum. Reprod. Update 2020, 26, 634–649. [Google Scholar] [CrossRef] [PubMed]
- Ozen, M.; Zhao, H.; Lewis, D.B.; Wong, R.J.; Stevenson, D.K. Heme oxygenase and the immune system in normal and pathological pregnancies. Front. Pharmacol. 2015, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Rahman, M.; Zhang, X.; Acevedo, C.H.; Nijjar, S.; Rushton, I.; Bussolati, B.; St John, J. Induction of placental heme oxygenase-1 is protective against TNFalpha-induced cytotoxicity and promotes vessel relaxation. Mol. Med. 2000, 6, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wong, R.J.; Kalish, F.S.; Nayak, N.R.; Stevenson, D.K. Effect of heme oxygenase-1 deficiency on placental development. Placenta 2009, 30, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Cudmore, M.; Ahmad, S.; Al-Ani, B.; Fujisawa, T.; Coxall, H.; Chudasama, K.; Devey, L.R.; Wigmore, S.J.; Abbas, A.; Hewett, P.W.; et al. Negative regulation of soluble Flt-1 and soluble endoglin release by heme oxygenase-1. Circulation 2007, 115, 1789–1797. [Google Scholar] [CrossRef] [Green Version]
- HAS. Suivi et Orientation des Femmes Enceintes en Fonction des Situations à Risque Identifiées. 2016. Available online: https://www.has-sante.fr/jcms/c_567727/fr/suivi-et-orientation-des-femmes-enceintes-en-fonction-des-situations-a-risque-identifiees (accessed on 16 December 2021).
- ACOG. ACOG Practice Bulletin No. 95: Anemia in pregnancy. Obstet. Gynecol. 2008, 112, 201–207. [Google Scholar] [CrossRef]
- WHO. Haemoglobin concentrations for the diagnosis of anaemia and assessment of severity. In Vitamin and Mineral Nutrition Information System Geneva; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Peyrin-Biroulet, L.; Williet, N.; Cacoub, P. Guidelines on the diagnosis and treatment of iron deficiency across indications: A systematic review. Am. J. Clin. Nutr. 2015, 102, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Van Den Broek, N.R.; Letsky, E.A.; White, S.A.; Shenkin, A. Iron status in pregnant women: Which measurements are valid? Br. J. Haematol. 1998, 103, 817–824. [Google Scholar] [CrossRef]
- Short, M.; Domagalski, J. Iron deficiency anemia: Evaluation and management. Am. Fam. Physician 2013, 87, 98–104. [Google Scholar] [PubMed]
- Harmon, A.C.; Cornelius, D.C.; Amaral, L.M.; Faulkner, J.L.; Cunningham, M.W.; Wallace, K.; LaMarca, B. The role of inflammation in the pathology of preeclampsia. Clin. Sci. 2016, 130, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, W.; Giussani, D.A. Preeclampsia link to gestational hypoxia. J. Dev. Orig. Health Dis. 2019, 10, 322–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, L.C.; Shennan, A.; Hyett, J.A.; Kapur, A.; Hadar, E.; Divakar, H.; McAuliffe, F.; da Silva Costa, F.; von Dadelszen, P.; McIntyre, H.D.; et al. The International Federation of Gynecology and Obstetrics (FIGO) initiative on pre-eclampsia: A pragmatic guide for first-trimester screening and prevention. Int. J. Gynecol. Obstet. 2019, 145, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, S.H.; Cheng, P.J.; Chung, T.T.; Kuo, C.F.; Wu, H.M.; Chu, P.H. Population-based trends and risk factors of early-and late-onset preeclampsia in Taiwan 2001–2014. BMC Pregnancy Childbirth 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Brunacci, F.; Rocha, V.S.; De Carli, E.; Espósito, B.P.; Ruano, R.; Colli, C. Increased serum iron in preeclamptic women is likely due to low hepcidin levels. Nutr. Res. 2018, 53, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Khatun, R.; Wu, Y.; Kanenishi, K.; Ueno, M.; Tanaka, S.; Hata, T. Immunohistochemical study of transferrin receptor expression in the placenta of pre-eclamptic pregnancy. Placenta 2003, 24, 870–876. [Google Scholar] [CrossRef]
- Gómez-Gutiérrez, A.M.; Parra-Sosa, B.E.; Bueno-Sánchez, J.C. Glycosylation Profile of the Transferrin Receptor in Gestational Iron Deficiency and Early-Onset Severe Preeclampsia. J. Pregnancy 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, M.; Schiff, E.; Kreiser, D.; Dennery, P.A.; Stevenson, D.K.; Rosenthal, T. End-tidal carbon monoxide measurements in women with pregnancy-induced hypertension and preeclampsia. Am. J. Obstet. Gynecol. 2000, 183, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Levytska, K.; Kingdom, J.; Baczyk, D.; Drewlo, S. Heme oxygenase-1 in placental development and pathology. Placenta 2013, 34, 291–298. [Google Scholar] [CrossRef]
- Conde-Agudelo, A.; Althabe, F.; Belizán, J.M.; Kafury-Goeta, A.C. Cigarette smoking during pregnancy and risk of preeclampsia: A systematic review. Am. J. Obstet. Gynecol. 1999, 181, 1026–1035. [Google Scholar] [CrossRef]
- Sidle, E.H.; Casselman, R.; Smith, G.N. Effect of cigarette smoke on placental antioxidant enzyme expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R754–R758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.X.; Chen, D.; Li, M.X.; Hua, Y. Increased serum iron levels in pregnant women with preeclampsia: A meta-analysis of observational studies. J. Obstet. Gynaecol. 2019, 39, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Shaji Geetha, N.; Bobby, Z.; Dorairajan, G.; Jacob, S.E. Increased hepcidin levels in preeclampsia: A protective mechanism against iron overload mediated oxidative stress? J. Matern. Fetal Neonatal Med. 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cardaropoli, S.; Todros, T.; Nuzzo, A.M.; Rolfo, A. Maternal serum levels and placental expression of hepcidin in preeclampsia. Pregnancy Hypertens. 2018, 11, 47–53. [Google Scholar] [CrossRef]
- Peyron, F.; L’ollivier, C.; Mandelbrot, L.; Wallon, M.; Piarroux, R.; Kieffer, F.; Hadjadj, E.; Paris, L.; Garcia-Meric, P. Maternal and Congenital Toxoplasmosis: Diagnosis and Treatment Recommendations of a French Multidisciplinary Working Group. Pathogens 2019, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Nowakowska, D.; Stray-Pedersen, B.; Spiewak, E.; Sobala, W.; Małafiej, E.; Wilczyński, J. Prevalence and estimated incidence of Toxoplasma infection among pregnant women in Poland: A decreasing trend in the younger population. Clin. Microbiol. Infect. 2006, 12, 913–917. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C.F.; Murray, H.W.; Wlebe, I.E.; Rubin, B.Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 1983, 158, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfefferkorn, E.R. Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proc. Natl. Acad. Sci. USA 1984, 81, 908–912. [Google Scholar] [CrossRef] [Green Version]
- Dimier, I.H.; Bout, D.T. Interferon-gamma-activated primary enterocytes inhibit Toxoplasma gondii replication: A role for intracellular iron. Immunology 1998, 94, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Ter Kuile, F.O.; Nosten, F.; McGready, R.; Asamoa, K.; Brabin, B. Epidemiology and burden of malaria in pregnancy. Lancet Infect. Dis. 2007, 7, 93–104. [Google Scholar] [CrossRef]
- Prentice, A.; Ghattas, H.; Doherty, C.; Cox, S. Iron metabolism and malaria. Food Nutr. Bull. 2007, 28 (Suppl. 4). [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mast, Q.; Syafruddin, D.; Keijmel, S.; Riekerink, T.O.; Deky, O.; Asih, P.B.; Swinkels, D.W.; van der Ven, A.J. Increased serum hepcidin and alterations in blood iron parameters associated with asymptomatic P. falciparum and P. vivax malaria. Haematologica 2010, 95, 1068–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mast, Q.; Nadjm, B.; Reyburn, H.; Kemna, E.; Amos, B.; Laarakkers, C.; Silalye, S.; Verhoef, H.; Sauerwein, R.W.; Swinkels, D.W.; et al. Assessment of urinary concentrations of hepcidin provides novel insight into disturbances in iron homeostasis during malarial infection. J. Infect. Dis. 2009, 199, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Accrombessi, M.; Yovo, E.; Fievet, N.; Cottrell, G.; Agbota, G.; Gartner, A.; Martin-Prevel, Y.; Vianou, B.; Sossou, D.; Fanou-Fogny, N.; et al. Effects of Malaria in the First Trimester of Pregnancy on Poor Maternal and Birth Outcomes in Benin. Clin. Infect. Dis. 2019, 69, 1385–1393. [Google Scholar] [CrossRef]
- Sangaré, L.; van Eijk, A.M.; Ter Kuile, F.O.; Walson, J.; Stergachis, A. The Association between Malaria and Iron Status or Supplementation in Pregnancy: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e87743. [Google Scholar] [CrossRef]
- Friedman, J.F.; Kurtis, J.D.; Kabyemela, E.R.; Fried, M.; Duffy, P.E. The iron trap: Iron, malaria and anemia at the mother–child interface. Microbes Infect. 2009, 11, 460–466. [Google Scholar] [CrossRef]
- Rogerson, S.J.; Desai, M.; Mayor, A.; Sicuri, E.; Taylor, S.M.; van Eijk, A.M. Burden, pathology, and costs of malaria in pregnancy: New developments for an old problem. Lancet Infect. Dis. 2018, 18, e107–e118. [Google Scholar] [CrossRef]
- Mwangi, M.N.; Roth, J.M.; Smit, M.R.; Trijsburg, L.; Mwangi, A.M.; Demir, A.Y.; Wielders, J.P.; Mens, P.F.; Verweij, J.J.; Cox, S.E.; et al. Effect of Daily Antenatal Iron Supplementation on Plasmodium Infection in Kenyan Women: A Randomized Clinical Trial. JAMA 2015, 314, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Etheredge, A.J.; Premji, Z.; Gunaratna, N.S.; Abioye, A.I.; Aboud, S.; Duggan, C.; Mongi, R.; Meloney, L.; Spiegelman, D.; Roberts, D.; et al. Iron supplementation in iron-replete and nonanemic pregnant women in Tanzania: A randomized clinical trial. JAMA Pediatr. 2015, 169, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Briand, V.; Saal, J.; Ghafari, C.; Huynh, B.T.; Fievet, N.; Schmiegelow, C.; Massougbodji, A.; Deloron, P.; Zeitlin, J.; Cot, M. Fetal Growth Restriction Is Associated With Malaria in Pregnancy: A Prospective Longitudinal Study in Benin. J. Infect. Dis. 2016, 214, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Santen, S.; De Mast, Q.; Luty, A.J.F.; Wiegerinck, E.T.; Van Der Ven, A.J.A.M.; Swinkels, D.W. Iron homeostasis in mother and child during placental malaria infection. Am. J. Trop. Med. Hyg. 2011, 84, 148–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteng-Ntim, E.; Ayensah, B.; Knight, M.; Howard, J. Pregnancy outcome in patients with sickle cell disease in the UK-a national cohort study comparing sickle cell anaemia (HbSS) with HbSC disease. Br. J. Haematol. 2015, 169, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Lesage, N.; Deneux Tharaux, C.; Saucedo, M.; Habibi, A.; Galacteros, F.; Girot, R.; Bouvier Coll, M.H.; Kayem, G. Maternal mortality among women with sickle-cell disease in France, 1996-2009. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 194, 183–188. [Google Scholar] [CrossRef]
- Costa, M.F.H.; Torres, L.C.; da Matta, M.C.; Araújo, A.d.S.; Souza, A.I. Interleukin-6 in pregnancy with sickle cell disease. Hematol. Transfus. Cell Ther. 2019, 41, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Wrighting, D.M.; Andrews, N.C. Interleukin-6 induces hepcidin expression through STAT3. Blood 2006, 108, 3204. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Peng, H.; Gelbart, T.; Wang, L.; Beutler, E. Regulation of hepcidin transcription by interleukin-1 and interleukin-6. Proc. Natl. Acad. Sci. USA 2005, 102, 1906–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, E.; Rivera, S.; Gabayan, V.; Keller, C.; Taudorf, S.; Pedersen, B.K.; Ganz, T. IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. J. Clin. Investig. 2004, 113, 1271–1276. [Google Scholar] [CrossRef] [Green Version]
- Aroke, D.; Kadia, B.M.; Njim, T. Iron stores in pregnant women with sickle cell disease: A systematic review. BMC Pregnancy Childbirth 2020, 20, 627. [Google Scholar] [CrossRef]
- Origa, R.; Piga, A.; Quarta, G.; Forni, G.L.; Longo, F.; Melpignano, A.; Galanello, R. Pregnancy and β-thalassemia: An Italian multicenter experience. Haematologica 2010, 95, 376–381. [Google Scholar] [CrossRef]
- Sanchaisuriya, K.; Fucharoen, S.; Ratanasiri, T.; Sanchaisuriya, P.; Fucharoen, G.; Dietz, E.; Schelp, F.P. Thalassemia and hemoglobinopathies rather than iron deficiency are major causes of pregnancy-related anemia in northeast Thailand. Blood Cells Mol. Dis. 2006, 37, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Jamnok, J.; Sanchaisuriya, K.; Sanchaisuriya, P.; Fucharoen, G.; Fucharoen, S.; Ahmed, F. Factors associated with anaemia and iron deficiency among women of reproductive age in Northeast Thailand: A cross-sectional study. BMC Public Health 2020, 20, 102. [Google Scholar] [CrossRef] [PubMed]
- Takhviji, V.; Zibara, K.; Azarkeivan, A.; Mehrvar, N.; Mehrvar, N.; Mezginejad, F.; Khosravi, A. Fertility and pregnancy in Iranian thalassemia patients: An update on transfusion complications. Transfus. Med. 2020, 30, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Devlieger, R.; Benhalima, K.; Damm, P.; Van Assche, A.; Mathieu, C.; Mahmood, T.; Dunne, F.; Bogaerts, A. Maternal obesity in Europe: Where do we stand and how to move forward?: A scientific paper commissioned by the European Board and College of Obstetrics and Gynaecology (EBCOG). Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 201, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Mayasari, N.R.; Hu, T.Y.; Chao, J.C.J.; Bai, C.H.; Chen, Y.C.; Huang, Y.L.; Chang, C.C.; Wang, F.F.; Hadi, H.; Nurwanti, E.; et al. Associations of the pre-pregnancy weight status with anaemia and the erythropoiesis-related micronutrient status. Public Health Nutr. 2021, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.D.; Zhao, G.; Jiang, Y.P.; Zhou, M.; Xu, G.; Kaciroti, N.; Zhang, Z.; Lozoff, B. Maternal obesity during pregnancy is negatively associated with maternal and neonatal iron status. Eur. J. Clin. Nutr. 2016, 70, 918–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, T.; Coupaye, M.; Esposito-Farèse, M.; Gault, N.; Talbi, N.; Quintin, C.; Schmitt, C.; Bekri, S.; Bado, A.; Puy, H.; et al. Hepcidin and Iron Deficiency in Women One Year after Sleeve Gastrectomy: A Prospective Cohort Study. Nutrients 2021, 13, 2516. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, B. Adipose tissue, inflammation and atherosclerosis. J. Atheroscler. Thromb. 2010, 17, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawer, A.A.; Hodyl, N.A.; Fairweather-Tait, S.; Froessler, B. Are Pregnant Women Who Are Living with Overweight or Obesity at Greater Risk of Developing Iron Deficiency/Anaemia? Nutrients 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. AMS 2017, 13, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Tzanavari, T.; Giannogonas, P.; Karalis, K. TNF-alpha and obesity. Curr. Dir. Autoimmun. 2010, 11, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Sen, S.; Iyer, C.; Klebenov, D.; Meydani, S.N. Obesity during pregnancy and fetal iron status: Is Hepcidin the link? J. Perinatol. Off. J. Calif. Perinat. Assoc. 2013, 33, 177–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tussing-Humphreys, L.; Nemeth, E.; Fantuzzi, G.; Freels, S.; Holterman, A.; Galvani, C.; Ayloo, S.; Vitello, J.; Braunschweig, C. Decreased serum hepcidin and improved functional iron status 6 months after restrictive bariatric surgery. Obesity 2010, 18, 2010–2016. [Google Scholar] [CrossRef] [Green Version]
- Devlieger, R.; Guelinckx, I.; Jans, G.; Voets, W.; Vanholsbeke, C.; Vansant, G. Micronutrient levels and supplement intake in pregnancy after bariatric surgery: A prospective cohort study. PLoS ONE 2014, 9, e114192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galazis, N.; Docheva, N.; Simillis, C.; Nicolaides, K.H. Maternal and neonatal outcomes in women undergoing bariatric surgery: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 181, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Weng, T.; Chang, C.; Dong, Y.; Chang, Y.; Chuang, L. Anaemia and related nutrient deficiencies after Roux-en-Y gastric bypass surgery: A systematic review and meta-analysis. BMJ Open 2015, 5, e006964. [Google Scholar] [CrossRef] [PubMed]
- Kemppinen, L.; Mattila, M.; Ekholm, E.; Pallasmaa, N.; Törmä, A.; Varakas, L.; Mäkikallio, K. Gestational iron deficiency anemia is associated with preterm birth, fetal growth restriction, and postpartum infections. J. Perinat. Med. 2021, 49, 431–438. [Google Scholar] [CrossRef]
- Sun, C.F.; Liu, H.; Hao, Y.H.; Hu, H.T.; Zhou, Z.Y.; Zou, K.X.; Liu, X.M.; Sheng, J.Z.; Ding, G.L.; Huang, H.F. Association between gestational anemia in different trimesters and neonatal outcomes: A retrospective longitudinal cohort study. World J. Pediatr. 2021, 42, 504–518. [Google Scholar] [CrossRef]
- Lao, T.T.; Tam, K.F.; Chan, L.Y. Third trimester iron status and pregnancy outcome in non-anaemic women; pregnancy unfavourably affected by maternal iron excess. Hum. Reprod. 2000, 15, 1843–1848. [Google Scholar] [CrossRef] [Green Version]
- Khambalia, A.; Collins, C.; Roberts, C.; Morris, J.; Powell, K.; Tasevski, V.; Nasar, N. High maternal serum ferritin in early pregnancy and risk of spontaneous preterm birth. Br. J. Nutr. 2015, 114, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sangkhae, V.; Fisher, A.L.; Wong, S.; Koenig, M.D.; Tussing-Humphreys, L.; Chu, A.; Lelic, M.; Ganz, T.; Nemeth, E. Effects of maternal iron status on placental and fetal iron homeostasis. J. Clin. Investig. 2020, 130, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Preziosi, P.; Prual, A.; Galan, P.; Daouda, H.; Boureima, H.; Hercberg, S. Effect of iron supplementation on the iron status of pregnant women: Consequences for newborns. Am. J. Clin. Nutr. 1997, 66, 1178–1182. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Georgieff, M.K. Iron Deficiency and Brain Development. Semin. Pediatr. Neurol. 2006, 13, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Goldenberg, R.L.; Hou, J.; Johnston, K.E.; Cliver, S.P.; Ramey, S.L.; Nelson, K.G. Cord Serum Ferritin Concentrations and Mental and Psychomotor Development of Children at Five Years of Age. Obstet. Gynecol. Surv. 2002, 57, 493–494. [Google Scholar] [CrossRef]
- Hajianfar, H.; Abbasi, K.; Azadbakht, L.; Esmaeilzadeh, A.; Mollaghasemi, N.; Arab, A. The Association between Maternal Dietary Iron Intake during the First Trimester of Pregnancy with Pregnancy Outcomes and Pregnancy-Related Complications. Clin. Nutr. Res. 2020, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cheng, Y.; Pei, L.; Jiang, Y.; Lei, F.; Zeng, L.; Wang, Q.; Li, Q.; Kang, Y.; Shen, Y.; et al. Maternal iron intake during pregnancy and birth outcomes: A cross-sectional study in Northwest China. Br. J. Nutr. 2017, 117, 862–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabe, H.; Gyte, G.M.; Díaz-Rossello, J.L.; Duley, L. Effect of timing of umbilical cord clamping and other strategies to influence placental transfusion at preterm birth on maternal and infant outcomes. Cochrane Database Syst. Rev. 2019, 2019. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| UK | ACOG | |||
|---|---|---|---|---|
| Laboratory Thresholds | Iron Dose | Laboratory Thresholds | Iron Dose | |
| Supplementation for anemic pregnant women |
11 g/dL (1st trimester)
10.5 g/dL (2nd and 3rd trimesters) | 100–200 mg iron / day |
11 g/dL (1st and 3rd trimesters)
| Full supplementation |
| Supplementation for non-anemic women with iron deficiency |
| 65 mg iron / day | - | - |
| Systematic supplementation for all women | - | Not recommended | - | Low dose iron in the first trimester |
| T1 | T2 | T3 | |
|---|---|---|---|
| Blood Hepcidin (nmol/L) | 1.85 [1,2,3,4] | <0.5 * | <0.5 * |
| TSAT (%) | 25 [15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35] | 20 [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] * | 10 [7.5–15] * |
| Blood Iron (µmol/L) | 15 [11,12,13,14,15,16,17,18,19,20,21,22,23,24] | 15 [11,12,13,14,15,16,17,18,19,20,21,22,23,24] | 9 [8,9,10,11,12,13,14,15] * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mégier, C.; Peoc’h, K.; Puy, V.; Cordier, A.-G. Iron Metabolism in Normal and Pathological Pregnancies and Fetal Consequences. Metabolites 2022, 12, 129. https://doi.org/10.3390/metabo12020129
Mégier C, Peoc’h K, Puy V, Cordier A-G. Iron Metabolism in Normal and Pathological Pregnancies and Fetal Consequences. Metabolites. 2022; 12(2):129. https://doi.org/10.3390/metabo12020129
Chicago/Turabian StyleMégier, Charles, Katell Peoc’h, Vincent Puy, and Anne-Gaël Cordier. 2022. "Iron Metabolism in Normal and Pathological Pregnancies and Fetal Consequences" Metabolites 12, no. 2: 129. https://doi.org/10.3390/metabo12020129
APA StyleMégier, C., Peoc’h, K., Puy, V., & Cordier, A. -G. (2022). Iron Metabolism in Normal and Pathological Pregnancies and Fetal Consequences. Metabolites, 12(2), 129. https://doi.org/10.3390/metabo12020129