
Metabolites Associated with Memory and Gait: A Systematic Review


Abstract
:1. Introduction
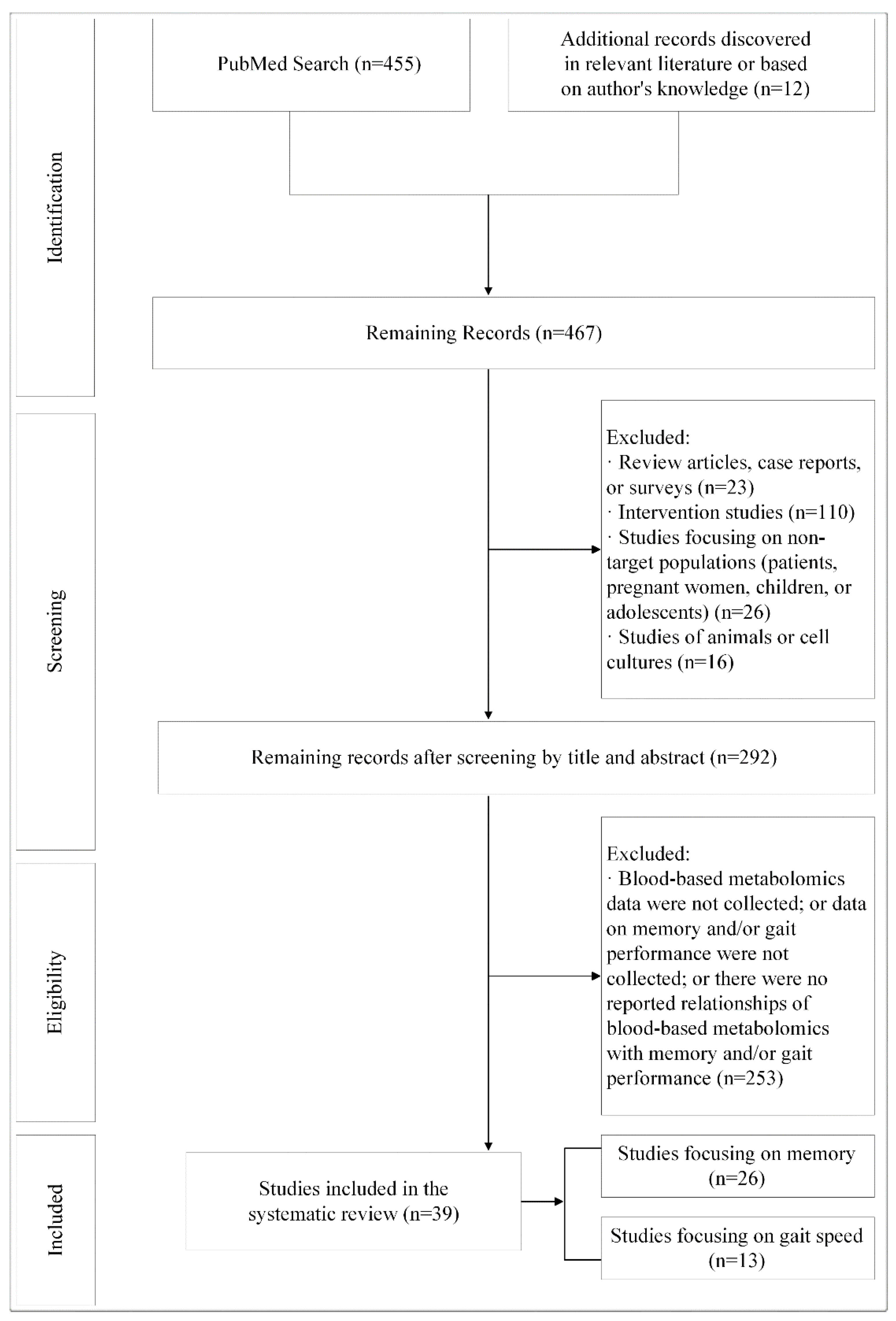
2. Materials and Methods
2.1. Literature Search and Study Selection
2.2. Analysis
3. Results
3.1. Overview
3.2. Metabolites and Memory Performance
3.3. Metabolites and Gait
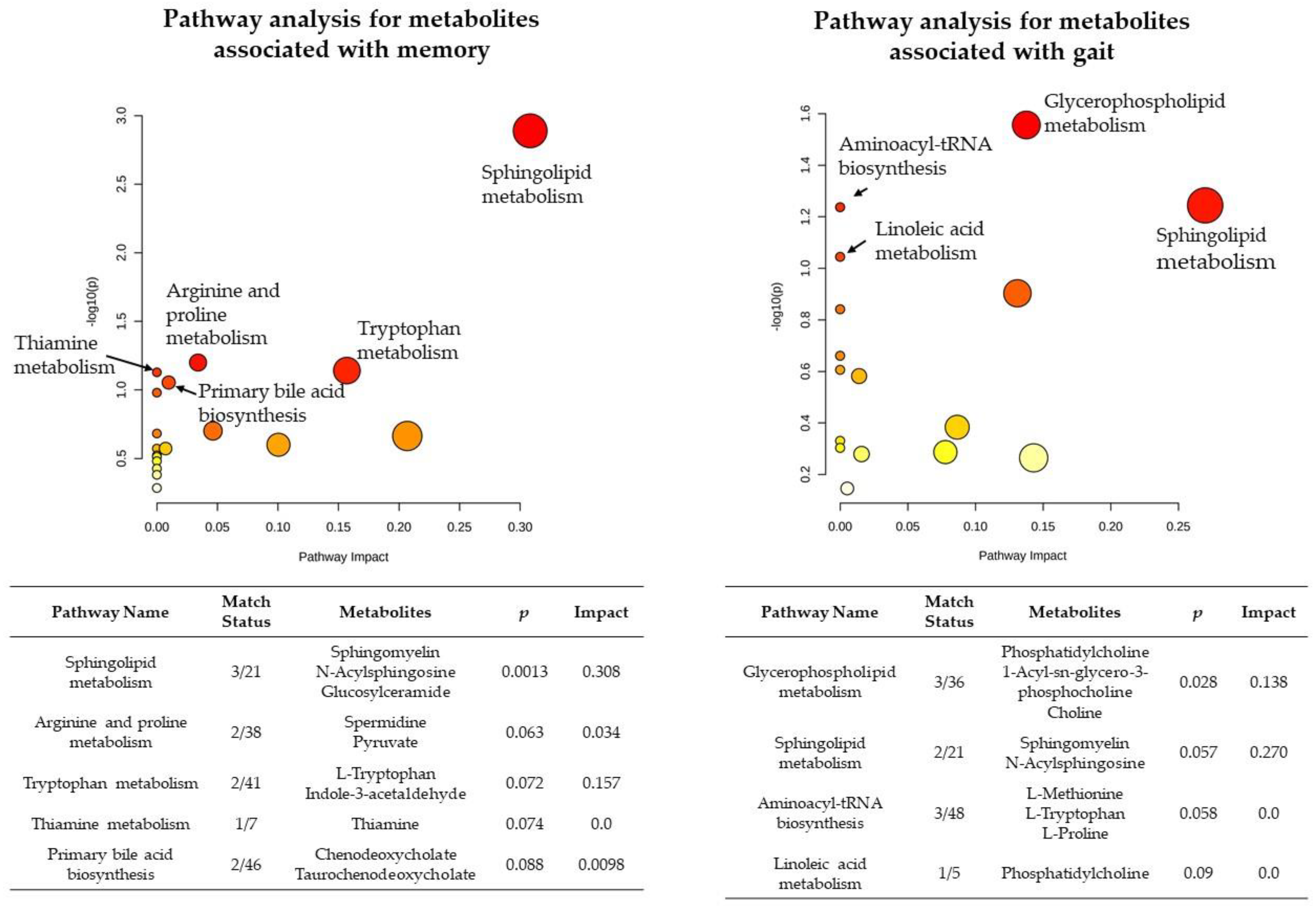
3.4. Pathway Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montero-Odasso, M.; Speechley, M.; Muir-Hunter, S.W.; Sarquis-Adamson, Y.; Sposato, L.A.; Hachinski, V.; Borrie, M.; Wells, J.; Black, A.; Sejdic, E.; et al. Motor and Cognitive Trajectories Before Dementia: Results from Gait and Brain Study. J. Am. Geriatr. Soc. 2018, 66, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Resnick, S.M.; Mielke, M.M.; Yaffe, K.; Launer, L.J.; Jonsson, P.V.; Grande, G.; Welmer, A.K.; Laukka, E.J.; Bandinelli, S.; et al. Association of Dual Decline in Memory and Gait Speed With Risk for Dementia Among Adults Older Than 60 Years: A Multicohort Individual-Level Meta-analysis. JAMA Netw. Open 2020, 3, e1921636. [Google Scholar] [CrossRef] [PubMed]
- Trushina, E.; Mielke, M.M. Recent advances in the application of metabolomics to Alzheimer’s Disease. Biochim. Biophys. Acta 2014, 1842, 1232–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, J.M.; Trushina, E. Application of Metabolomics in Alzheimer’s Disease. Front. Neurol. 2017, 8, 719. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.F.; Chevallier, O.P.; Elliott, C.T.; Holscher, C.; Johnston, J.; McGuinness, B.; Kehoe, P.G.; Passmore, A.P.; Green, B.D. Untargeted metabolomic analysis of human plasma indicates differentially affected polyamine and L-arginine metabolism in mild cognitive impairment subjects converting to Alzheimer’s disease. PLoS ONE 2015, 10, e0119452. [Google Scholar] [CrossRef]
- Pagani, M.; Nobili, F.; Morbelli, S.; Arnaldi, D.; Giuliani, A.; Oberg, J.; Girtler, N.; Brugnolo, A.; Picco, A.; Bauckneht, M.; et al. Early identification of MCI converting to AD: A FDG PET study. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 2042–2052. [Google Scholar] [CrossRef]
- Choi, S.; Reiter, D.A.; Shardell, M.; Simonsick, E.M.; Studenski, S.; Spencer, R.G.; Fishbein, K.W.; Ferrucci, L. 31P Magnetic Resonance Spectroscopy Assessment of Muscle Bioenergetics as a Predictor of Gait Speed in the Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1638–1645. [Google Scholar] [CrossRef] [Green Version]
- Coen, P.M.; Jubrias, S.A.; Distefano, G.; Amati, F.; Mackey, D.C.; Glynn, N.W.; Manini, T.M.; Wohlgemuth, S.E.; Leeuwenburgh, C.; Cummings, S.R.; et al. Skeletal muscle mitochondrial energetics are associated with maximal aerobic capacity and walking speed in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 447–455. [Google Scholar] [CrossRef]
- Gonzalez-Freire, M.; Scalzo, P.; D’Agostino, J.; Moore, Z.A.; Diaz-Ruiz, A.; Fabbri, E.; Zane, A.; Chen, B.; Becker, K.G.; Lehrmann, E.; et al. Skeletal muscle ex vivo mitochondrial respiration parallels decline in vivo oxidative capacity, cardiorespiratory fitness, and muscle strength: The Baltimore Longitudinal Study of Aging. Aging Cell 2018, 17, e12725. [Google Scholar] [CrossRef] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Bazzano, L.A.; He, J.; Gu, X.; Li, C.; Li, S.; Yaffe, K.; Kinchen, J.M.; Stuchlik, P.; Mi, X.; et al. Novel serum metabolites associate with cognition phenotypes among Bogalusa Heart Study participants. Aging 2019, 11, 5124–5139. [Google Scholar] [CrossRef] [PubMed]
- Darst, B.F.; Huo, Z.; Jonaitis, E.M.; Koscik, R.L.; Clark, L.R.; Lu, Q.; Kremen, W.S.; Franz, C.E.; Rana, B.; Lyons, M.J.; et al. Metabolites Associated with Early Cognitive Changes Implicated in Alzheimer’s Disease. J. Alzheimers Dis. 2021, 79, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Snowden, S.; Suvitaival, T.; Ali, A.; Merkler, D.J.; Ahmad, T.; Westwood, S.; Baird, A.; Proitsi, P.; Nevado-Holgado, A.; et al. Primary fatty amides in plasma associated with brain amyloid burden, hippocampal volume, and memory in the European Medical Information Framework for Alzheimer’s Disease biomarker discovery cohort. Alzheimers Dement. 2019, 15, 817–827. [Google Scholar] [CrossRef]
- Du, Y.; Wei, J.; Yang, X.; Dou, Y.; Zhao, L.; Qi, X.; Yu, X.; Guo, W.; Wang, Q.; Deng, W.; et al. Plasma metabolites were associated with spatial working memory in major depressive disorder. Medicine 2021, 100, e24581. [Google Scholar] [CrossRef]
- Kurella Tamura, M.; Chertow, G.M.; Depner, T.A.; Nissenson, A.R.; Schiller, B.; Mehta, R.L.; Liu, S.; Sirich, T.L.; Study, F.H.N. Metabolic Profiling of Impaired Cognitive Function in Patients Receiving Dialysis. J. Am. Soc. Nephrol. 2016, 27, 3780–3787. [Google Scholar] [CrossRef]
- Proitsi, P.; Kuh, D.; Wong, A.; Maddock, J.; Bendayan, R.; Wulaningsih, W.; Hardy, R.; Richards, M. Lifetime cognition and late midlife blood metabolites: Findings from a British birth cohort. Transl. Psychiatry 2018, 8, 203. [Google Scholar] [CrossRef] [Green Version]
- Bressler, J.; Yu, B.; Mosley, T.H.; Knopman, D.S.; Gottesman, R.F.; Alonso, A.; Sharrett, A.R.; Wruck, L.M.; Boerwinkle, E. Metabolomics and cognition in African American adults in midlife: The atherosclerosis risk in communities study. Transl. Psychiatry 2017, 7, e1173. [Google Scholar] [CrossRef]
- Mapstone, M.; Cheema, A.K.; Fiandaca, M.S.; Zhong, X.; Mhyre, T.R.; MacArthur, L.H.; Hall, W.J.; Fisher, S.G.; Peterson, D.R.; Haley, J.M.; et al. Plasma phospholipids identify antecedent memory impairment in older adults. Nat. Med. 2014, 20, 415–418. [Google Scholar] [CrossRef]
- Li, D.; Misialek, J.R.; Boerwinkle, E.; Gottesman, R.F.; Sharrett, A.R.; Mosley, T.H.; Coresh, J.; Wruck, L.M.; Knopman, D.S.; Alonso, A. Plasma phospholipids and prevalence of mild cognitive impairment and/or dementia in the ARIC Neurocognitive Study (ARIC-NCS). Alzheimers Dement. 2016, 3, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Varma, V.R.; Oommen, A.M.; Varma, S.; Casanova, R.; An, Y.; Andrews, R.M.; O’Brien, R.; Pletnikova, O.; Troncoso, J.C.; Toledo, J.; et al. Brain and blood metabolite signatures of pathology and progression in Alzheimer disease: A targeted metabolomics study. PLoS Med. 2018, 15, e1002482. [Google Scholar] [CrossRef] [PubMed]
- Huo, Z.; Yu, L.; Yang, J.; Zhu, Y.; Bennett, D.A.; Zhao, J. Brain and blood metabolome for Alzheimer’s dementia: Findings from a targeted metabolomics analysis. Neurobiol. Aging 2020, 86, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre, D.A.; Slupsky, C.M.; Aviv, R.I.; Swardfager, W.; Taha, A.Y. Untargeted metabolomic analysis of plasma from relapsing-remitting multiple sclerosis patients reveals changes in metabolites associated with structural changes in brain. Brain Res. 2020, 1732, 146589. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Hennebelle, M.; Sahlas, D.J.; Ramirez, J.; Gao, F.; Masellis, M.; Cogo-Moreira, H.; Swartz, R.H.; Herrmann, N.; Chan, P.C.; et al. Soluble Epoxide Hydrolase-Derived Linoleic Acid Oxylipins in Serum Are Associated with Periventricular White Matter Hyperintensities and Vascular Cognitive Impairment. Transl. Stroke Res. 2019, 10, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Solvang, S.H.; Nordrehaug, J.E.; Tell, G.S.; Nygard, O.; McCann, A.; Ueland, P.M.; Midttun, O.; Meyer, K.; Vedeler, C.A.; Aarsland, D.; et al. The kynurenine pathway and cognitive performance in community-dwelling older adults. The Hordaland Health Study. Brain Behav. Immun. 2019, 75, 155–162. [Google Scholar] [CrossRef]
- Mielke, M.M.; Bandaru, V.V.; Haughey, N.J.; Rabins, P.V.; Lyketsos, C.G.; Carlson, M.C. Serum sphingomyelins and ceramides are early predictors of memory impairment. Neurobiol. Aging 2010, 31, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Arnoriaga-Rodriguez, M.; Mayneris-Perxachs, J.; Burokas, A.; Contreras-Rodriguez, O.; Blasco, G.; Coll, C.; Biarnes, C.; Miranda-Olivos, R.; Latorre, J.; Moreno-Navarrete, J.M.; et al. Obesity Impairs Short-Term and Working Memory through Gut Microbial Metabolism of Aromatic Amino Acids. Cell Metab. 2020, 32, 548–560.e7. [Google Scholar] [CrossRef]
- Kindler, J.; Lim, C.K.; Weickert, C.S.; Boerrigter, D.; Galletly, C.; Liu, D.; Jacobs, K.R.; Balzan, R.; Bruggemann, J.; O’Donnell, M.; et al. Dysregulation of kynurenine metabolism is related to proinflammatory cytokines, attention, and prefrontal cortex volume in schizophrenia. Mol. Psychiatry 2020, 25, 2860–2872. [Google Scholar] [CrossRef] [Green Version]
- Borkowski, K.; Taha, A.Y.; Pedersen, T.L.; De Jager, P.L.; Bennett, D.A.; Arnold, M.; Kaddurah-Daouk, R.; Newman, J.W. Serum metabolomic biomarkers of perceptual speed in cognitively normal and mildly impaired subjects with fasting state stratification. Sci. Rep. 2021, 11, 18964. [Google Scholar] [CrossRef]
- Mielke, M.M.; Haughey, N.J.; Bandaru, V.V.; Schech, S.; Carrick, R.; Carlson, M.C.; Mori, S.; Miller, M.I.; Ceritoglu, C.; Brown, T.; et al. Plasma ceramides are altered in mild cognitive impairment and predict cognitive decline and hippocampal volume loss. Alzheimers Dement. 2010, 6, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.; Saleem, M.; Herrmann, N.; Mielke, M.M.; Haughey, N.J.; Oh, P.I.; Kiss, A.; Lanctot, K.L. Ceramide Accumulation Is Associated with Declining Verbal Memory in Coronary Artery Disease Patients: An Observational Study. J. Alzheimers Dis. 2018, 64, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuster, O.C.; Laptinskaya, D.; Fissler, P.; Schnack, C.; Zugel, M.; Nold, V.; Thurm, F.; Pleiner, S.; Karabatsiakis, A.; von Einem, B.; et al. Novel Blood-Based Biomarkers of Cognition, Stress, and Physical or Cognitive Training in Older Adults at Risk of Dementia: Preliminary Evidence for a Role of BDNF, Irisin, and the Kynurenine Pathway. J. Alzheimers Dis. 2017, 59, 1097–1111. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.N.; Kim, M.; Chuang, Y.F.; Beason-Held, L.; Kitner-Triolo, M.; Kraut, M.; Lirette, S.T.; Windham, B.G.; Griswold, M.E.; Legido-Quigley, C.; et al. Blood metabolite markers of cognitive performance and brain function in aging. J. Cereb. Blood Flow Metab. 2016, 36, 1212–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, M.M.; Xue, Q.L.; Zhou, J.; Chaves, P.H.; Fried, L.P.; Carlson, M.C. Baseline serum cholesterol is selectively associated with motor speed and not rates of cognitive decline: The Women’s Health and Aging Study II. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Becklen, M.; Orhan, F.; Piehl, F.; Cervenka, S.; Sellgren, C.M.; Flyckt, L.; Erhardt, S.; Fatouros-Bergman, H. Plasma bilirubin levels are reduced in first-episode psychosis patients and associates to working memory and duration of untreated psychosis. Sci. Rep. 2021, 11, 7527. [Google Scholar] [CrossRef]
- Wang, L.J.; Lin, P.Y.; Lee, Y.; Huang, Y.C.; Wu, C.C.; Hsu, S.T.; Chen, C.C.; Chong, M.Y.; Lin, C.H.; Hung, C.F. Increased serum levels of cysteine in patients with schizophrenia: A potential marker of cognitive function preservation. Schizophr. Res. 2018, 192, 391–397. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Canas, J.A.; Dore, G.A.; Beydoun, H.A.; Rostant, O.S.; Fanelli-Kuczmarski, M.T.; Evans, M.K.; Zonderman, A.B. Serum Uric Acid and Its Association with Longitudinal Cognitive Change Among Urban Adults. J. Alzheimers Dis. 2016, 52, 1415–1430. [Google Scholar] [CrossRef] [Green Version]
- Nierenberg, J.L.; He, J.; Li, C.; Gu, X.; Shi, M.; Razavi, A.C.; Mi, X.; Li, S.; Bazzano, L.A.; Anderson, A.H.; et al. Serum metabolites associate with physical performance among middle-aged adults: Evidence from the Bogalusa Heart Study. Aging 2020, 12, 11914–11941. [Google Scholar] [CrossRef]
- Marron, M.M.; Wendell, S.G.; Boudreau, R.M.; Clish, C.B.; Santanasto, A.J.; Tseng, G.C.; Zmuda, J.M.; Newman, A.B. Metabolites Associated with Walking Ability Among the Oldest Old from the CHS All Stars Study. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 2371–2378. [Google Scholar] [CrossRef]
- Murphy, R.A.; Moore, S.; Playdon, M.; Kritchevsky, S.; Newman, A.B.; Satterfield, S.; Ayonayon, H.; Clish, C.; Gerszten, R.; Harris, T.B. Metabolites Associated With Risk of Developing Mobility Disability in the Health, Aging and Body Composition Study. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Freire, M.; Moaddel, R.; Sun, K.; Fabbri, E.; Zhang, P.; Khadeer, M.; Salem, N., Jr.; Ferrucci, L.; Semba, R.D. Targeted Metabolomics Shows Low Plasma Lysophosphatidylcholine 18:2 Predicts Greater Decline of Gait Speed in Older Adults: The Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Misialek, J.R.; Huang, F.; Windham, G.B.; Yu, F.; Alonso, A. Independent Association of Plasma Hydroxysphingomyelins With Physical Function in the Atherosclerosis Risk in Communities (ARIC) Study. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Kameda, M.; Teruya, T.; Yanagida, M.; Kondoh, H. Frailty markers comprise blood metabolites involved in antioxidation, cognition, and mobility. Proc. Natl. Acad. Sci. USA 2020, 117, 9483–9489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lum, H.; Sloane, R.; Huffman, K.M.; Kraus, V.B.; Thompson, D.K.; Kraus, W.E.; Bain, J.R.; Stevens, R.; Pieper, C.F.; Taylor, G.A.; et al. Plasma acylcarnitines are associated with physical performance in elderly men. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 548–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Karagounis, L.G.; Ng, T.P.; Carre, C.; Narang, V.; Wong, G.; Tan, C.T.Y.; Zin Nyunt, M.S.; Gao, Q.; Abel, B.; et al. Systemic and Metabolic Signature of Sarcopenia in Community-Dwelling Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 309–317. [Google Scholar] [CrossRef]
- Meng, L.; Yang, R.; Wang, D.; Wu, W.; Shi, J.; Shen, J.; Dang, Y.; Fan, G.; Shi, H.; Dong, J.; et al. Specific lysophosphatidylcholine and acylcarnitine related to sarcopenia and its components in older men. BMC Geriatr. 2022, 22, 249. [Google Scholar] [CrossRef]
- Wennberg, A.M.V.; Schafer, M.J.; LeBrasseur, N.K.; Savica, R.; Bui, H.H.; Hagen, C.E.; Hollman, J.H.; Petersen, R.C.; Mielke, M.M. Plasma Sphingolipids are Associated With Gait Parameters in the Mayo Clinic Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Frison, E.; Boirie, Y.; Peuchant, E.; Tabue-Teguo, M.; Barberger-Gateau, P.; Feart, C. Plasma fatty acid biomarkers are associated with gait speed in community-dwelling older adults: The Three-City-Bordeaux study. Clin. Nutr. 2017, 36, 416–422. [Google Scholar] [CrossRef]
- Jang, I.Y.; Park, J.H.; Kim, J.H.; Lee, S.; Lee, E.; Lee, J.Y.; Park, S.J.; Kim, D.A.; Hamrick, M.W.; Kim, B.J. The association of circulating kynurenine, a tryptophan metabolite, with frailty in older adults. Aging 2020, 12, 22253–22265. [Google Scholar] [CrossRef]
- He, W.; Luo, Y.; Liu, J.P.; Sun, N.; Guo, D.; Cui, L.L.; Zheng, P.P.; Yao, S.M.; Yang, J.F.; Wang, H. Trimethylamine N-Oxide, a Gut Microbiota-Dependent Metabolite, is Associated with Frailty in Older Adults with Cardiovascular Disease. Clin. Interv. Aging 2020, 15, 1809–1820. [Google Scholar] [CrossRef]
- Trayssac, M.; Hannun, Y.A.; Obeid, L.M. Role of sphingolipids in senescence: Implication in aging and age-related diseases. J. Clin. Investig. 2018, 128, 2702–2712. [Google Scholar] [CrossRef] [PubMed]
- De Larichaudy, J.; Zufferli, A.; Serra, F.; Isidori, A.M.; Naro, F.; Dessalle, K.; Desgeorges, M.; Piraud, M.; Cheillan, D.; Vidal, H.; et al. TNF-alpha- and tumor-induced skeletal muscle atrophy involves sphingolipid metabolism. Skelet. Muscle 2012, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, D.; Wegner, M.S.; Wanger, R.A.; Ferreiros, N.; Schreiber, Y.; Lucks, J.; Schiffmann, S.; Geisslinger, G.; Grosch, S. The equilibrium between long and very long chain ceramides is important for the fate of the cell and can be influenced by co-expression of CerS. Int. J. Biochem. Cell Biol. 2013, 45, 1195–1203. [Google Scholar] [CrossRef]
- Stiban, J.; Perera, M. Very long chain ceramides interfere with C16-ceramide-induced channel formation: A plausible mechanism for regulating the initiation of intrinsic apoptosis. Biochim. Biophys. Acta 2015, 1848, 561–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaurasia, B.; Talbot, C.L.; Summers, S.A. Adipocyte Ceramides-The Nexus of Inflammation and Metabolic Disease. Front. Immunol. 2020, 11, 576347. [Google Scholar] [CrossRef]
- Boon, J.; Hoy, A.J.; Stark, R.; Brown, R.D.; Meex, R.C.; Henstridge, D.C.; Schenk, S.; Meikle, P.J.; Horowitz, J.F.; Kingwell, B.A.; et al. Ceramides contained in LDL are elevated in type 2 diabetes and promote inflammation and skeletal muscle insulin resistance. Diabetes 2013, 62, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Pralhada Rao, R.; Vaidyanathan, N.; Rengasamy, M.; Mammen Oommen, A.; Somaiya, N.; Jagannath, M.R. Sphingolipid metabolic pathway: An overview of major roles played in human diseases. J. Lipids 2013, 2013, 178910. [Google Scholar] [CrossRef] [Green Version]
- Le Stunff, H.; Veret, J.; Kassis, N.; Denom, J.; Meneyrol, K.; Paul, J.L.; Cruciani-Guglielmacci, C.; Magnan, C.; Janel, N. Deciphering the Link Between Hyperhomocysteinemia and Ceramide Metabolism in Alzheimer-Type Neurodegeneration. Front. Neurol. 2019, 10, 807. [Google Scholar] [CrossRef] [Green Version]
- Del Campo, N.; Payoux, P.; Djilali, A.; Delrieu, J.; Hoogendijk, E.O.; Rolland, Y.; Cesari, M.; Weiner, M.W.; Andrieu, S.; Vellas, B.; et al. Relationship of regional brain beta-amyloid to gait speed. Neurology 2016, 86, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Wennberg, A.M.V.; Lesnick, T.G.; Schwarz, C.G.; Savica, R.; Hagen, C.E.; Roberts, R.O.; Knopman, D.S.; Hollman, J.H.; Vemuri, P.; Jack, C.R., Jr.; et al. Longitudinal Association Between Brain Amyloid-Beta and Gait in the Mayo Clinic Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1244–1250. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; Resnick, S.M.; Bilgel, M.; Wong, D.F.; Ferrucci, L.; Studenski, S.A. beta-Amyloid Burden Predicts Lower Extremity Performance Decline in Cognitively Unimpaired Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 716–723. [Google Scholar] [PubMed] [Green Version]
- Freitas, H.R.; Isaac, A.R.; Malcher-Lopes, R.; Diaz, B.L.; Trevenzoli, I.H.; De Melo Reis, R.A. Polyunsaturated fatty acids and endocannabinoids in health and disease. Nutr. Neurosci. 2018, 21, 695–714. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Frisardi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Vendemiale, G.; Capurso, A.; Panza, F. Dietary fatty acids and predementia syndromes. Sci. World J. 2009, 9, 792–810. [Google Scholar] [CrossRef] [PubMed]
- Samieri, C.; Feart, C.; Letenneur, L.; Dartigues, J.F.; Peres, K.; Auriacombe, S.; Peuchant, E.; Delcourt, C.; Barberger-Gateau, P. Low plasma eicosapentaenoic acid and depressive symptomatology are independent predictors of dementia risk. Am. J. Clin. Nutr. 2008, 88, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotrich, F.E.; Sears, B.; McNamara, R.K. Elevated ratio of arachidonic acid to long-chain omega-3 fatty acids predicts depression development following interferon-alpha treatment: Relationship with interleukin-6. Brain Behav. Immun. 2013, 31, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [Green Version]
- Mingam, R.; Moranis, A.; Bluthe, R.M.; De Smedt-Peyrusse, V.; Kelley, K.W.; Guesnet, P.; Lavialle, M.; Dantzer, R.; Laye, S. Uncoupling of interleukin-6 from its signalling pathway by dietary n-3-polyunsaturated fatty acid deprivation alters sickness behaviour in mice. Eur. J. Neurosci. 2008, 28, 1877–1886. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Li, X.; Leonard, B.E.; Horrobin, D.F. Effects of dietary n-3 or n-6 fatty acids on interleukin-1beta-induced anxiety, stress, and inflammatory responses in rats. J. Lipid Res. 2003, 44, 1984–1991. [Google Scholar] [CrossRef] [Green Version]
- Fernstrom, J.D. Branched-chain amino acids and brain function. J. Nutr. 2005, 135, 1539S–1546S. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.; Riddle, E.; Barre, L. Protein and amino acids for skeletal muscle health in aging. Adv. Food Nutr. Res. 2020, 91, 29–64. [Google Scholar]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357, eaaf9794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, A.; Tankiewicz-Kwedlo, A.; Krupa, A.; Pawlak, D. Role of Kynurenine Pathway in Oxidative Stress during Neurodegenerative Disorders. Cells 2021, 10, 1603. [Google Scholar] [CrossRef] [PubMed]
- Sas, K.; Szabo, E.; Vecsei, L. Mitochondria, Oxidative Stress and the Kynurenine System, with a Focus on Ageing and Neuroprotection. Molecules 2018, 23, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Study Name (First Author, Year) | n (Women%) Age, Mean (SD), Median (IRQ), Range, Cognitive Status | Race/Ethnicity (%) | Memory Assessment | Metabolomics Technique | Sample Type; Number of Metabolites Analyzed (Classes) | Threshold for Statistical Significance |
|---|---|---|---|---|---|---|
| Bogalusa Heart Study (Shi et al., 2019) * [12] | n = 1177, 59.7% 48.11 (5.26) | White (65%) and Black (35%) | WAIS-IV for working memory and WMS-IV for verbal memory | UPLC-MS/MS (Metabolon Inc., Durham, NC, USA) | Serum 1466 (1202 analyzed, including AAs, FAs, carbohydrates, and nucleotides) | FDR (Bonferroni correction); p < 4.16 × 10−5 (=0.05/1202) |
| WRAP (Darst et al., 2021) † [13] | n = 2324, 68.8% 62 (6.8), range 40–81 | White and non-Hispanic (95%) | Composite score for delayed recall from RAVLT, WMS-R LM, and BVMT-R | UPLC-MS/MS (Metabolon Inc., Durham, NC, USA) | Plasma 1097 (untargeted, including AAs, FAs, carbohydrates, and nucleotides) | FDR (Benjamini-Hochberg correction); q < 0.05 |
| EMIF-AD Multimodal Biomarker Discovery Study (Kim et al., 2019) * [14] | n = 593, 53% CN: 65.06 (7.93) MCI: 70.44 (7.86) AD: 69.55 (8.51) | Not specified (European, 100%) | AVLT for immediate and delayed verbal memory | UPLC-MS/MS (Metabolon Inc., Durham, NC, USA) | plasma 883 (648 analyzed, then focused on only 9, including AAs and FAs) | FDR (Bonferroni correction); p < 7.72 × 10−5 (=0.05/648) |
| Mental Health Center of West China Hospital, Sichuan University (Du et al., 2021) * [15] | n = 83 (controls); 62.7% 26.4 (8.62), range 18–60 | East Asian (Chinese, 100%) | Neuropsychological Tests Automated Battery for spatial working memory | LC-MS/MS | plasma 728 (296 analyzed, including AAs, acylcarnitines, biogenic amines, carbohydrates, LPCs, and PCs) | Spearman rank correlation p < 0.05 |
| Outpatient Dialysis Clinics in Northern California (Kurella Tamura et al., 2016) * [16] | n = 141, 36% 56.6 (14.6) | White (42.6%) | Controlled Oral Word Association for verbal memory and language and RAVLT for delayed recall. | GC & LC-MS/MS (Metabolon Inc., Durham, NC, USA) | plasma 562 (95 analyzed, including AA derivatives) | FDR (Benjamini-Hochberg correction) q < 0.05 |
| MRC NSHD British 1946 Birth Cohort (Proitsi et al., 2018) *,† [17] | n = 909; 52% range 60–64 | Not specified (British: English, Scottish, and Welsh, 100%) | Three-trial 15-item word list learning task for short-term verbal memory and an uncued delayed free recall trial. | NMR | serum 233 (including FAs, and AAs) | Multiple testing correction; p < 0.002 (=0.05/principal components) |
| Community-Dwelling African American Participants in the Biracial ARIC study (Bressler et al., 2017) † [18] | n = 1534 (n = 1393 without incident dementia); 63.6% 53.4 (5.8), range 45–64 | Black (African American, 100%) | DWRT for verbal memory | GC/MS and LC-MS (Metabolon Inc., Durham, NC, USA) | serum 204 (including AAs and FAs) | FDR (Dubey/Armitage-Parmar correction); p < 3.9 × 10−4 |
| Rochester/Orange County Aging Study (Mapstone et al., 2017) * [19] | n = 224, 62% superior memory: 83.2 (3.4) normal control: 82.3 (3.6) MCI/AD: 81.9 (4.4) | Not specified | RAVLT for verbal memory | Triple quadrupole MS, SID-MRM-MS, and FIA MS/MS (Biocrates, Innsbruck, Austria, p180) | plasma 188 (185 analyzed, then focused on only 12, including AAs, acylcarnitines, PCs, LPCs, SLs, and biogenic amines) | p < 0.05 |
| ARIC study (Li et al., 2016) * [20] | n = 441, 54.42% CN: 77.6 (5.5) MCI: 76.5 (5.6) Dementia: 79.7 (5.1) | Black (African American, 85.1%) | Delayed word recall, logical memory test part A and B, and incidental learning | triple-quadrupole MS (Biocrates, Innsbruck, Austria, p180) | Plasma 188 (main analysis focused on 9 metabolites including PCs and LPCs; additional analyses explored 151) | p < 0.05 for 9 metabolites in main analysis. FDR (Bonferroni correction) for 151 metabolites in exploratory analysis p < 0.00033 (=0.05/151) |
| BLSA (Varma et al., 2018) *,† [21] | n = 207, 51.69% 78.68 (7.23) | White (83.09%) | CVLT for learning and immediate and long delay free recall | FIA-MS/MS and HPLC-MS/MS (Biocrates, Innsbruck, Austria, p180) | serum 187 (20 analyzed including AAs, SLs, PCs, acylcarnitines, and biogenic amines) | p < 0.05 |
| ROS and MAP (Huo et al., 2020) † [22] | n = 530, 78.5% 82 (7.4) | White (European origin, 100%) | episode, working, and semantic memory | FIA-MS/MS and UHPLC-MS/MS (Biocrates, Innsbruck, Austria, p180) | serum 182 (including AAs, biogenic amines, acylcarnitines, PCs, and SLs) | FDR (Benjamini-Hochberg correction); q ≤ 10% |
| Sunnybrook Hospital (Sylvestre et al., 2020) * [23] | n = 18 (controls); 66.7% 48.7 (7.2) | Not specified (Canadian, 100%) | BVMT-R for visuospatial memory | 1H-NMR spectroscopy | plasma 56 (9 analyzed, mostly AAs) | Spearman’s rank correlation, p < 0.05; post-hoc FDR (Bonferroni correction) |
| Stroke Prevention Clinic (Yu et al., 2019) * [24] | n = 25 (healthy controls with minimal SIVD); 54% 71.7 (7.9), range 50–85 | Not specified (Canadian, 100%) | CVLT-II for verbal memory (short delayed free recall, long-delayed recall, and recall discriminability) | UPLC-MS/MS | serum Not specified (24 analyzed, oxylipins only) | FDR (Bonferroni correction) |
| Hordaland Health Study (Solvang et al., 2019) * [25] | n = 2174, 55.2% median 71, range 70–72 | Not specified (Norwegian, 100%) | KOLT for immediate recall and COWAT for verbal memory | LC-MS/MS | plasma 12 (targeted, AAs and biogenic amines only) | FDR (Bonferroni correction); p < 0.0042 (=0.05/12) |
| WHAS II (Mielke, Bandaru et al., 2010) *,† [26] | n = 100 (100%) 74 (2.5), 70–79 | Black (African American, 23%) | HVLT-R for verbal immediate and delayed recall | ESI/MS/MS | serum Not specified (12 analyzed, including SLs and cholesterols) | p < 0.05 |
| Josep Trueta University Hospital (Arnoriaga et al., 2020) * [27] | n = 116; 69.8% median 50.4, IQR: 41.8–58.5 | Not specified (Spanish, 100%) | CVLT for immediate and short delayed recall and TDS for working memory | LC-MS/MS (Scharlau, Barcelona, Spain) | plasma Not specified (untargeted, including AAs, FAs, Indoles, and Phenylpropanoic acids) | Variable importance measure from random forest algorithm |
| Living Cohort (Kindler et al., 2020) * [28] | n = 81 (healthy controls); 50.6% 31.7 (8.5) | Not specified (Australian, 100%) | WAIS-III LNS for working memory and WMS-R LM for verbal memory | UHPLC and GC-MS (Agilent, Santa Clara, CA, USA) | plasma Not specified (targeted, kynurenine pathway metabolites only) | p < 0.05 |
| ROS and MAP (Borkowski et al., 2021) * [29] | n = 198 (59 fasted); 88% 78.2 (7.2) | White and non-Hispanic (95%) | Global measures of episodic, semantic, and working memory from 17 tests | LC-MS/MS | serum Not specified (targeted, lipid mediators only) | Spearman’s rank correlation, p < 0.05 |
| Community-Dwelling Volunteers Recruited From the Clinical Core of the Johns Hopkins Alzheimer’s Disease Research Center (Mielke, Haughey et al., 2010) * [30] | n = 63; 39.7%; CN: 74.4 (7.0) MCI: 74.5 (5.6) AD: 74.8 (7.0) All 55+ | White (96%) | CVLT for verbal memory and Logical Memory Story A from the Wechsler Memory Scale for immediate and delayed recall. | HPLC/MS/MS | plasma 8 SLs (2 analyzed, Cer only) | p < 0.05 |
Cardiac Rehab Program at the Rumsey Centre of University Health Network Toronto Rehab Institute (Chan et al., 2018) † [31] | n = 60, 16.7% 64.6 (6), range 50–75 46 CN and 14 with possible MVND (sMMSE <24 excluded); all had CAD | White (79.7%) | CVLT-II for verbal memory and BVMT-R for visuospatial memory. | LC/MS/MS | plasma Not specified (5 analyzed, including SLs) | p ≤ 0.05 |
| Sensory-cognitive and Physical Fitness Training in Mild Cognitive Impairment Study (Küster et al., 2017) *,† [32] | n = 47, 57.4% 71.2 (6), range 60–88 | Not specified (German, 100%) | German CVLT for verbal memory and Everyday Cognition Battery for working memory | Enzyme-linked Immunosorbent Assay kit (Promega Corporation, Madison, WI, USA), spectrophotometer, and LC-MS/MS | serum 6 (targeted, mostly kynurenine pathway metabolites) | p < 0.05 |
| BLSA (Simpson et al., 2016) *,† [33] | n = 107, 39.25% 72.92 (7.61) | Not specified | CVLT for verbal memory in short and delayed recall tests. BVRT for visual memory. | UPLC-MS | plasma 3 (targeted, PCs only) | p < 0.005 |
| WHAS II (Mielke et al., 2008) † [34] | n = 426, 100% 74.5 (2.8), range 70–79 | Black (African American, 19%) | HVLT-R for verbal immediate and delayed memory | Total/HDL cholesterol levels were calculated using standard enzymatic techniques. LDL calculated using Friedewald equation. | serum Not specified (3 analyzed, including FAs and cholesterols) | p < 0.05 |
| Karolinska Schizophrenia Project (Becklén et al., 2021) * [35] | n = 22 (healthy controls); 50% median 25, IQR: 22–28 | Not specified (Swedish, 100%) | WMSIII for working memory: Spatial Span and Letter-Number Span | Colorimetry (Roche Diagnostics, Basel, Switzerland) | plasma 1 (targeted, bilirubin only) | Spearman’s rank correlation, p < 0.05 |
| Kaohsiung Chang Gung Memorial Hospital (Wang et al., 2018) * [36] | n = 65 (healthy controls); 44.6% 40.1 (12), range 18–65 | East Asian (Chinese, 100%) | List Learning Test for verbal memory and Digit Sequencing Task for working memory | MicroMolar Cysteine Assay Kit (ProFoldin, Hudson, MA, USA) | serum 1 (targeted, cysteine only) | p < 0.05 |
| HANDLS Study (Beydoun et al., 2016) *,† [37] | n = 2630, 56.6% 47 (0.3), range 30–64 | Not specified | CVLT for immediate and delayed free recall and BVRT for visual memory. | Spectrophotometry (Quest Diagnostics, Secaucus, NJ, USA) | serum Not specified (1 analyzed, uric acid only) | FDR (Bonferroni correction for multiple cognitive tests); p < 0.004 (=0.05/11) |
| Study Name (First Author, Year) | n (Women%) age, Mean (SD), Median (IRQ), Range, Cognitive Status | Race/Ethnicity (%) | Gait Assessment | Metabolomics Technique | Sample Type Number of Metabolites Analyzed (Classes) | Threshold of Significance |
|---|---|---|---|---|---|---|
| Bogalusa Heart Study (Nierenberg et al., 2020) *,† [38] | n = 1239; 58.92% 48.2 (5.3) | White (65.5%) | 6-minute walk | UPLC-MS/MS (Metabolon Inc. Durham, NC, USA) | serum 1466 (1202 analyzed, including AAs, carbohydrates, FAs, LPCs and SLs) | p < 0.05 |
| CHS All Stars Study (Marron et al., 2020) * [39] | n = 120, 60% 85(2.9), range 79–95 | White (90%) | 15 ft walk | LC-MS | plasma 605 (569 analyzed, including AAs and FAs) | p < 0.05 and FDR (Benjamini-Hochberg correction); q ≤ 30% |
| Health ABC Study (Murphy et al., 2019) * [40] | n = 313, 0% 74.6(2.8), range 70–79 | Black (African American, 100%) | 20 m usual walking speed | LC-MS (Broad Institute of MIT and Harvard, Cambridge, MA, USA) | plasma 350 (including FAs, AAs, SLs, PCs) | p ≤ 0.01 and q ≤ 0.3 |
| BLSA (Gonzalez-Freire et al., 2019) *,† [41] | n = 504, 49% 70.7 (9.9), all 50+ | Not specified | 6 m walk | LC-MS/MS (Biocrates, Innsbruck, Austria, p180) | plasma 188 (148 analyzed, including AAs, SLs, PCs, acylcarnitines, biogenic amines, and LPCs) | Spearman rank correlations, p < 0.05 and FDR (multiple testing correction); q < 0.05 |
| ARIC Study (Li et al., 2018) * [42] | n = 383, 52.5% 77.5 (5.5) | White (75%) | 4 m walk | triple-quadrupole mass spectrometer (Biocrates, Innsbruck, Austria, p180) | plasma 188 (12 analyzed, including PCs and SLs) | p < 0.05 |
| Kyoto University Hospital (Kameda et al., 2020) [43] | n = 19 (10 non-frail); 63.2% 84.2 (6.9) | East Asian (Japanese, 100%) | TUG | LC-MS/MS (Thermo Fisher Scientific, Waltham, MA, USA) | whole blood 131 (untargeted, including AAs, acylcarnitines, and lactones) | p < 0.05 |
| U.S. Veterans LIFE Study (Lum et al., 2011) * [44] | n = 77, 0% 79.2 (4.8), all 70+ | Not specified | 8 ft walk 400 m walk | MS | plasma 45 (Acylcarnitines only; PCA score) | p < 0.05 |
| Singapore Longitudinal Ageing Study Wave 2 (Lu et al., 2020) * [45] | n = 189; 63% Sarcopenia:73.9 (5.3), No sarcopenia:72.5 (5.3) Range 65–90 | East Asian (Chinese, 100%) | 6 m walk | N/A (Bevital Lab, Bergen, Norway) | plasma Not specified (27 analyzed, including AAs) | p < 0.05 |
| Geriatric Medicine Department of Beijing Hospital (Meng et al., 2022) * [46] | n = 246; 0% Sarcopenia: 80.9 (8.5) Nonsarcopenia: 78.6 (7.4) Range 61–100 | East Asian (Chinese, 100%) | 6 m walk | LC-MS/MS (Sciex and Agilent, Santa Clara, CA, USA) | Serum Not specified (targeted, including AAs, acylcarnitines, and LPCs) | p < 0.05 |
| Mayo Clinic Study of Aging (Wennberg et al., 2018) * [47] | n = 340, 38.2% median 80.3, IQR:77.2–83.7 range 70–95 | Not specified | GAITRite-5.6 m electronic walk-way | LC/ESI/MS/MS (Sciex, Agilent, Santa Clara, CA, USA) | plasma Not specified (12 analyzed, including SLs) | p ≤ 0.05 |
| Bordeaux Centre of the Three-City Study (Frison et al., 2017) * [48] | n = 982, 59.1% Low gait speed: 75.5 (4.7) Not low gait:73.6 (4.8) All 65+ | Not specified (French, 100%) | 6 m walk | GC | plasma Not specified (12 analyzed, FAs only) | p < 0.005 |
| Division of Geriatrics of the Department of Internal Medicine of the Asan Medical Center in Seoul, South Korea (Jang et al., 2020) * [49] | n = 73, 56.2% robust: 67.6 (6.8) pre-frail: 69.8 (5.9) frail: 70.8 (5.0) | East Asian (South Korean, 100%) | 4 m walk | LC-MS/MS | serum 3 (targeted, Kynurenine, tryptophan, and ratio of the two) | p < 0.05 |
| National Center of Gerontology (He et al., 2020) * [50] | n = 451 (316 non-frail), 47% 75.2 (6.6), all 65+ | East Asian (Chinese, 100%) | 15 ft walk time for slowness | UPLC-MS/MS (Waters Corp, Milford, MA USA) | plasma 1 (targeted, Trimethylamine N-Oxide only) | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Q.; Mitchell, B.A.; Corkum, A.E.; Moaddel, R.; Ferrucci, L. Metabolites Associated with Memory and Gait: A Systematic Review. Metabolites 2022, 12, 356. https://doi.org/10.3390/metabo12040356
Tian Q, Mitchell BA, Corkum AE, Moaddel R, Ferrucci L. Metabolites Associated with Memory and Gait: A Systematic Review. Metabolites. 2022; 12(4):356. https://doi.org/10.3390/metabo12040356
Chicago/Turabian StyleTian, Qu, Brendan A. Mitchell, Abigail E. Corkum, Ruin Moaddel, and Luigi Ferrucci. 2022. "Metabolites Associated with Memory and Gait: A Systematic Review" Metabolites 12, no. 4: 356. https://doi.org/10.3390/metabo12040356
APA StyleTian, Q., Mitchell, B. A., Corkum, A. E., Moaddel, R., & Ferrucci, L. (2022). Metabolites Associated with Memory and Gait: A Systematic Review. Metabolites, 12(4), 356. https://doi.org/10.3390/metabo12040356