
Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses

Abstract
:1. Introduction
2. Results
2.1. Identification of Metabolites
2.2. Principal Component Analysis (PCA) of Metabolites in Xylem and Phloem Sap of Cucumber under Phosphorus Stress
2.3. PLS-DA Analysis of Metabolites in Xylem and Phloem Sap of Cucumber under Phosphorus Stress
2.4. Screening of Differential Metabolites
2.5. Cluster Analysis of Differential Metabolites
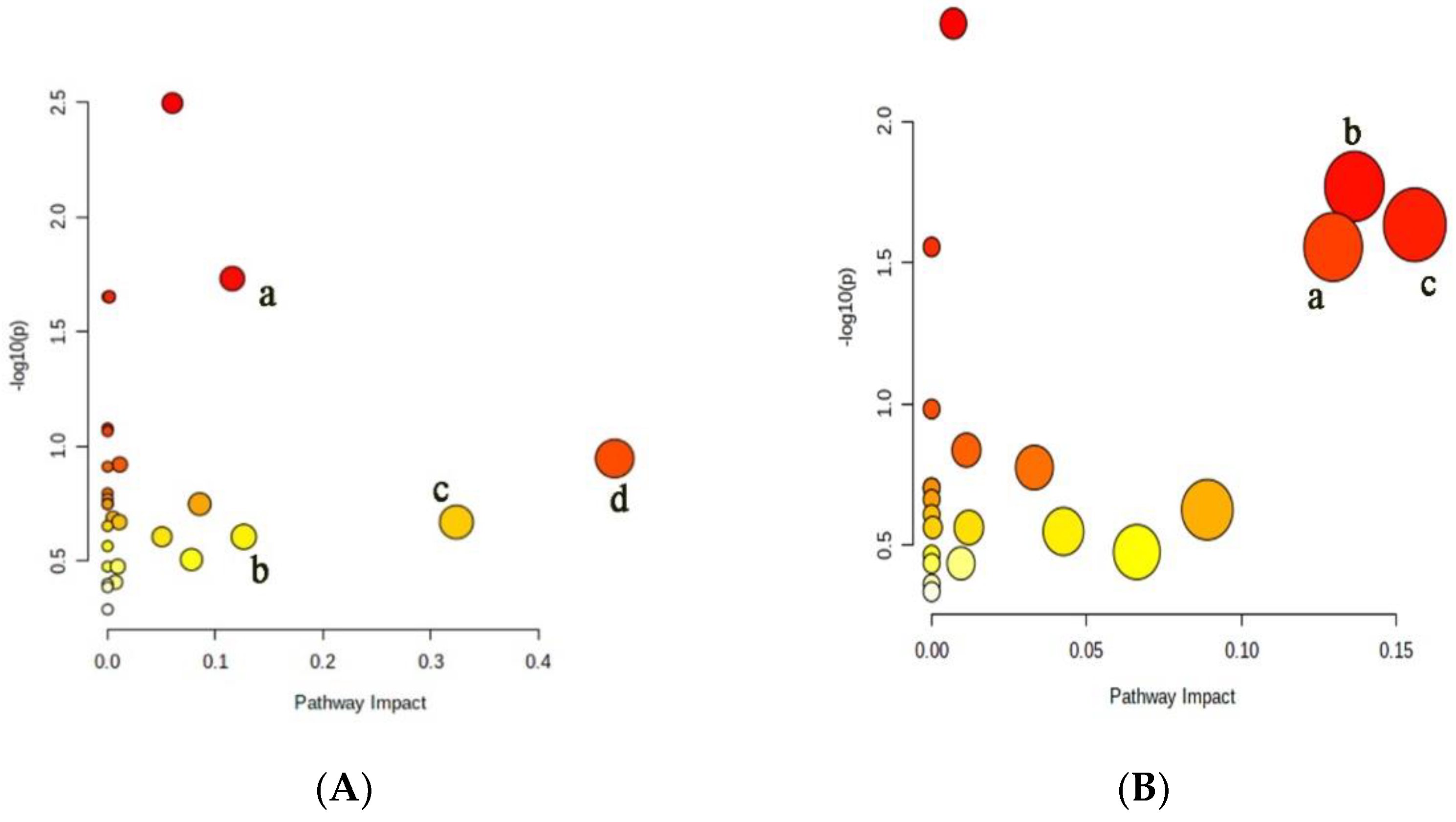
2.6. Metabolic Pathway Analysis of Differential Metabolites
3. Discussion
4. Materials and Methods
4.1. Experimental Instruments and Materials
4.2. Plant Sample Cultivation
4.3. Sampling and Measurement
4.3.1. Sampling of Xylem and Phloem Sap of Cucumber
4.3.2. GC-MS Metabolite Detection
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, J.; Meng, F.; Yu, D.; Qiu, Q.; Zhang, M.; Rao, D.; Cong, B.; Zhang, W.; Yan, X. Response of agronomic traits and P/Fe utilization efficiency to P application with different Pefficiency in soybean. Acta. Agron. Sin. 2021, 47, 1824–1833. [Google Scholar]
- Lin, Z.; Chen, R.; Guo, S. Research Progress on Physiological Adaptability of Plants to Phosphorus Deficiency. Crops 2010, 5, 5–9. [Google Scholar]
- Peng, X. Analysis of Metabolic and Phosphatase Gene Expression Profile in Cajanus Cajan Roots under Low Phosphorus Stress. Master’s Thesis, Hainan University, Haikou, China, 2019. [Google Scholar]
- Chen, L.; Huang, S.; Sun, Y.; Zhu, E.; Wang, K. Rapid Identification of Potassium Nutrition Stress in Rice Based on Machine Vision and Object-Oriented Segmentation. J. Spectrosc. 2019, 2019, 4623545. [Google Scholar] [CrossRef]
- Gao, H.; Mao, H.; Ullah, I. Analysis of metabolomic changes in lettuce leaves under low nitrogen and phosphorus deficiencies stresses. Agriculture 2020, 10, 406. [Google Scholar] [CrossRef]
- Guo, R.; Zhou, J.; Yang, F.; Li, F. Metabolic responses of wheat roots to alkaline stress. Chin. J. Plant Ecol. 2017, 41, 683–692. [Google Scholar]
- Li, Q.; Li, L.; Hu, C.; Tan, Q.; Sun, X. Metabolic response and molybdenum tolerance mechanism of winter wheat to excess molybdenum stress. J. Huazhong Agric. Univ. 2021, 4, 36–43. [Google Scholar]
- Yang, M.; Yang, J.; Su, L. Metabolic profile analysis and identification of key metabolites during rice seed germination under low-temperature stress. Plant Sci. 2019, 289, 110282. [Google Scholar] [CrossRef]
- Yu, X.; Wang, J.; Zhu, M.; Jiang, J. Molecular Mechanism of Carbohydrate Metabolism Participation in Tomato Response to Low Phosphorus Stress. Mol. Plant Breed. 2015, 12, 2833–2842. [Google Scholar]
- Duan, X.; Zhao, Y.; Zhang, J. Characteristics of the root exudate release system of typical plants in plateau lakeside wetland under phosphorus stress conditions. Open Chem. 2020, 18, 808–821. [Google Scholar] [CrossRef]
- Muhammad, I.; Abdullah, S.; Saud, H.; Shaharuddin, N. The dynamic responses of oil palm leaf and root metabolome to phosphorus deficiency. Metabolites 2021, 11, 217. [Google Scholar] [CrossRef]
- Xie, C.; Cui, J.; Li, Y.; Zhang, J.; Liu, Q.; Gong, H.; Lei, J.; Wang, F. GC-MS Analysis of Metabolites and Gene Expression Characteristics of Dioscorea zingiberensis in Response to Low Phosphorus Stress. Chin. J. Exp. Tradit. 2022, 1–13. [Google Scholar]
- Ding, Z.; Jia, S.; Wang, Y.; Xiao, J.; Zhang, Y. Phosphate stresses affectionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 120, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Ceballos, L.; Grusak, M. Plant fluid proteomics: Delving into the Xylem Sap, Phloem Sap and Apoplastic Fluid Proteomes. BBA Proteins Proteom. 2016, 1864, 991–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, W.; Groover, A.; Lichtenberger, A. The plant vascular system: Evolution, Development and Functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef]
- Yang, C.; Tan, T.; Yu, J.; Liao, Q.; Zhang, X.; Zhang, Z.; Song, H.; Guan, C. Effects of atmospheric CO2 enrichment on phloem sap composition and root nitrogen accumulation in oilseed rape. Chin. J. Plant Ecol. 2014, 38, 776–784. [Google Scholar]
- Tang, F.; Teng, J.; Wei, B.; Huang, L.; Ning, Q. Evaluation of the influence of traditional fermentation on the composition of phenolic compounds in Hakka pickled mustard greens based on non-targeted metabolomics. Food Ferment. Ind. 2021, 47, 128–133. [Google Scholar]
- Wang, F. Study on Effect of Potassium on Quality and the Main Correlative Mechanism of Cucumber in Solar-Greenhouse. Master’s Thesis, Shandong Agriculture University, Taian, China, 2005. [Google Scholar]
- Li, Y. Study on Mechanism of Nano Potassium Silicate in Regulating Nutrition and Flavor Quality of Different Vegetables. Master’s Thesis, Southwest University, Chongqing, China, 2019. [Google Scholar]
- Chen, Z.; Chen, T.; Li, H.; Cui, X. Research Progress in Molecular Mechanism of Amino Acids Absorption and Transport by Plants. Mol. Plant Breed. 2017, 15, 5166–5171. [Google Scholar]
- Li, Y.; Hou, D.; Yue, H.; Zhang, D. Effect of Inoculating Fusarium oxysporum on Seedings Growth and Leaf Free Amino Acids of Resistant and Susceptible Cucumbers. J. Agric. Sci. Technol. 2019, 21, 94–102. [Google Scholar]
- Wang, C. Effect of Parching-Field on Yield and Physio-Ecological Characteristics of Early and Late Dual Purpose Super Rice in South China. Master’s Thesis, South China Agnicultural University, Guangzhou, China, 2016. [Google Scholar]
- Wang, J.; Tang, Z. The Regulation of Soluble Sugars in the Growth and Development of Plants. Bot. Res. 2014, 3, 71–76. [Google Scholar]
- Zhang, S.; Wang, C.; Feng, Y. Analysis of Metabolites in Rice Produced in Different Regions by GC-MS-based Metabonomics. Food Sci. 2019, 40, 17990. [Google Scholar]
- Hu, M.; Liu, H.; Pan, L.; Wang, T.; Li, X.; Wang, Y.; Fang, H. Analysis of Metabolic Differences of Goji Juice Fermented by Lactobacillus paracasei Based on Non-targeted Metabonomics. Food Sci. 2022, 1–12. [Google Scholar]
- Zhou, L.; Li, Y.; Wang, W.; Zhong, S. Research Progress in the Metabolomics for Plants Response to Temperature Stress. J. Shanxi Agric. Sci. 2017, 45, 317–320. [Google Scholar]
- Hu, L.; Xu, Q. Review of Current Progress in the Metabolomics for Plant Response to Abiotic Stress. Crop Res. 2014, 28, 428–434. [Google Scholar]
- Liu, C.; Xing, J.; Wei, F.; Dong, J.; Zhu, X.; Hong, L. Effects of exogenous nitrate nitrogen on glutamate accumulation and key enzymes activities in metabolism of Portulaca oleracea L.under salt stress. Jiangsu Agric. Sci. 2020, 48, 130–136. [Google Scholar]
- Qiu, F. Cloning of Phenylalanine Ammonia-Lyase from Caragana Korshinkii Kom, Construction of RNAi Vector and Anaylysis of its Function. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2006. [Google Scholar]
- Wang, G. Studies on Disease Resistance and Related Enzyme Activity of Different Corn. Master’s Thesis, Yanbian University, Yanji, China, 2010. [Google Scholar]
- Ye, W.; Yun, H.; Fan, L.; Guo, S. Effects of Compound Coating Antistaling Agent of Polysaccharide Treatment in Storage on POD, PPO and PAL Enzymatic Activity of Chinese Bayberry. Packag. Food. Mach. 2012, 30, 10–16. [Google Scholar]
- Lu, M. Effects of low phosphorus stress on the composition and content of free amino acids in soil orange and trifoliate orange. South China Fruits 2017, 46, 39–42. [Google Scholar]
- Chen, H.; Li, X.; Li, Z. Sugar Signaling and Its Role in Plant Response to Environmental Stress. Biotechnol. Bull. 2022, 38, 1–10. [Google Scholar]
- Jin, X.; Chen, H.; Shi, Y.; Bai, L.; Hou, L.; Zhang, Y. Effect of citric acid seed priming on the growth and physiological characteristics of tomato seedlings under low phosphorus stress. Chin. J. Eco. Agric. 2021, 29, 1159–1170. [Google Scholar]
- Jia, H.; Yin, G.; Huang, H.; Liu, W.; Liu, G.; Cui, H. Influence of Low Phosphorus Stress on Glucose Metabolism and Nutrition Accumulation in Tobacco Yunyan 87. J. Agric. Sci. Technol. 2014, 16, 36–41. [Google Scholar]
- Zhao, X.; Chen, M.; Zhao, Y.; Zha, L.; Yang, H.; Wu, Y. GC–MS-Based Nontargeted and Targeted Metabolic Profiling Identifies Changes in the Lentinula Edodes Mycelial Metabolome under High-Temperature Stress. Int. J. Mol. Sci. 2019, 20, 2330. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, M.; Song, X.; Li, Q.; Qi, X.; Wang, L. The Application of Metabonomics in Study of Plant under Heavy Metal Stress. Biol. Chem. Eng. 2020, 6, 128–132. [Google Scholar]
- Guo, L.; Shi, D.; Ma, C. Metabolic regulation and secretion of organic acids in plants in response to stress. J. Chang. Educ. Inst. 2005, 3, 19–24. [Google Scholar]
- Zhu, W.; Mao, H.; Zhou, Y.; Zhang, X. Hyperspectral imaging technology of nitrogen status diagnose for tomato leaves. J. Jiangsu Nat. Sci Ed. 2014, 35, 290–294. [Google Scholar]
- Zhu, W.; Mao, H.; Li, Q.; Liu, H.; Sun, J.; Zuo, Z.; Chen, Y. Study on the Polarized Reflectance-Hyperspectral Information Fusion Technology of Tomato Leaves Nutrient Diagnoses. Spectrosc. Spect. Anal. 2014, 34, 2500–2505. [Google Scholar]
- Nie, P.; Li, C.; Gao, Y.; Du, G.; Lv, D.; Guan, Q. Progress on phloem unloading pathway of photoassimilate. Plant Physiol. J. 2019, 55, 697–702. [Google Scholar]
- Ge, T.; Jiang, W.; Song, S. Influence of inorganic and amino acid nitrogen on mineral nutrient contents in xylem and phloem sap of different tomato (Solanum lycopersicum) cultivars. Acta Hortic. Sin. 2009, 36, 347–354. [Google Scholar]
- Fan, H.; Xu, Y.; Du, C. Phloem sap proteome studied by iTRAQ provides integrated insight into salinity response mechanisms in cucumber plants. J. Proteom. 2015, 125, 54–67. [Google Scholar] [CrossRef]
- Yue, Y.; Liu, S.; Yin, C.; Guo, R.; Yang, M.; Tian, S.; Hu, H. Study on the Flower Bud Metabolomics of Petunia hybrida Fantasy Grown Under Weak Light or Short Day Conditions. Acta Hortic. Sin. 2017, 44, 2391–2398. [Google Scholar]
- Yao, K. UItra High Performance Liquid Chromatography-Mass Spectrometry Based Metabolomics Research on Gastric Cancer Tissue. Master’s Thesis, Zhengzhou University, Zhengzhou, China, 2017. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Xylem | Phloem | ||||
|---|---|---|---|---|---|
| Metabolites | p | VIP | Metabolites | p | VIP |
| Stearic acid | *** | 3.12029 | Galactitol | *** | 3.33729 |
| Palmitic acid | *** | 2.89369 | Palmitic acid | *** | 3.12769 |
| Phenylalanine | *** | 2.51584 | Stearic acid | *** | 2.93922 |
| Ethanolamine | *** | 2.39481 | Galactaric acid | *** | 2.69905 |
| Malic acid | *** | 2.15182 | Pipecolic acid | *** | 2.64936 |
| 2-Monostearin | *** | 1.84387 | Trimethylsilyl ester | *** | 2.44461 |
| Beta-Sitosterol | *** | 1.67496 | Ethanolamine | *** | 2.28112 |
| Citric acid | *** | 1.61834 | D-Fructose | *** | 2.15505 |
| Silane | *** | 1.48680 | γ-aminobutyric acid | *** | 1.76774 |
| Glycerolmonostearate | *** | 1.45735 | Oxalic acid | *** | 1.63812 |
| Galactitol | *** | 1.44325 | 1-Monopalmitin | *** | 1.51261 |
| Phosphoenolpyruvate | *** | 1.34310 | Glycerolmonostearate | *** | 1.49872 |
| 1-Monopalmitin | *** | 1.31531 | Glucose | *** | 1.46972 |
| Glutamate | *** | 1.31233 | Mannitol | *** | 1.27220 |
| Oxalic acid | *** | 1.29589 | Benzene | ** | 1.26719 |
| Benzene | *** | 1.14935 | Sucrose | *** | 1.15488 |
| D-Xylitol | *** | 1.12737 | Homoserine | *** | 1.15416 |
| Trisiloxane | *** | 1.12260 | Malic acid | ** | 1.15056 |
| Galactaric acid | *** | 1.02495 | Dehydroabietic acid | *** | 1.13394 |
| Aminoadipic acid | *** | 1.00987 | Succinic acid | ** | 1.11712 |
| Heptasiloxane | *** | 1.00980 | Citric acid | ** | 1.05460 |
| Ethane | *** | 1.00030 | Pyroglutamic acid | *** | 1.00308 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Li, Q.; Xu, H.; Zhang, W. Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses. Metabolites 2022, 12, 361. https://doi.org/10.3390/metabo12040361
Sun J, Li Q, Xu H, Zhang W. Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses. Metabolites. 2022; 12(4):361. https://doi.org/10.3390/metabo12040361
Chicago/Turabian StyleSun, Jingjing, Qinglin Li, Hui Xu, and Wentao Zhang. 2022. "Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses" Metabolites 12, no. 4: 361. https://doi.org/10.3390/metabo12040361
APA StyleSun, J., Li, Q., Xu, H., & Zhang, W. (2022). Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses. Metabolites, 12(4), 361. https://doi.org/10.3390/metabo12040361