
A Spectroscopic Technique to Simultaneously Characterize Fatty Acid Uptake, Mitochondrial Activity, Vascularity, and Oxygen Saturation for Longitudinal Studies In Vivo

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
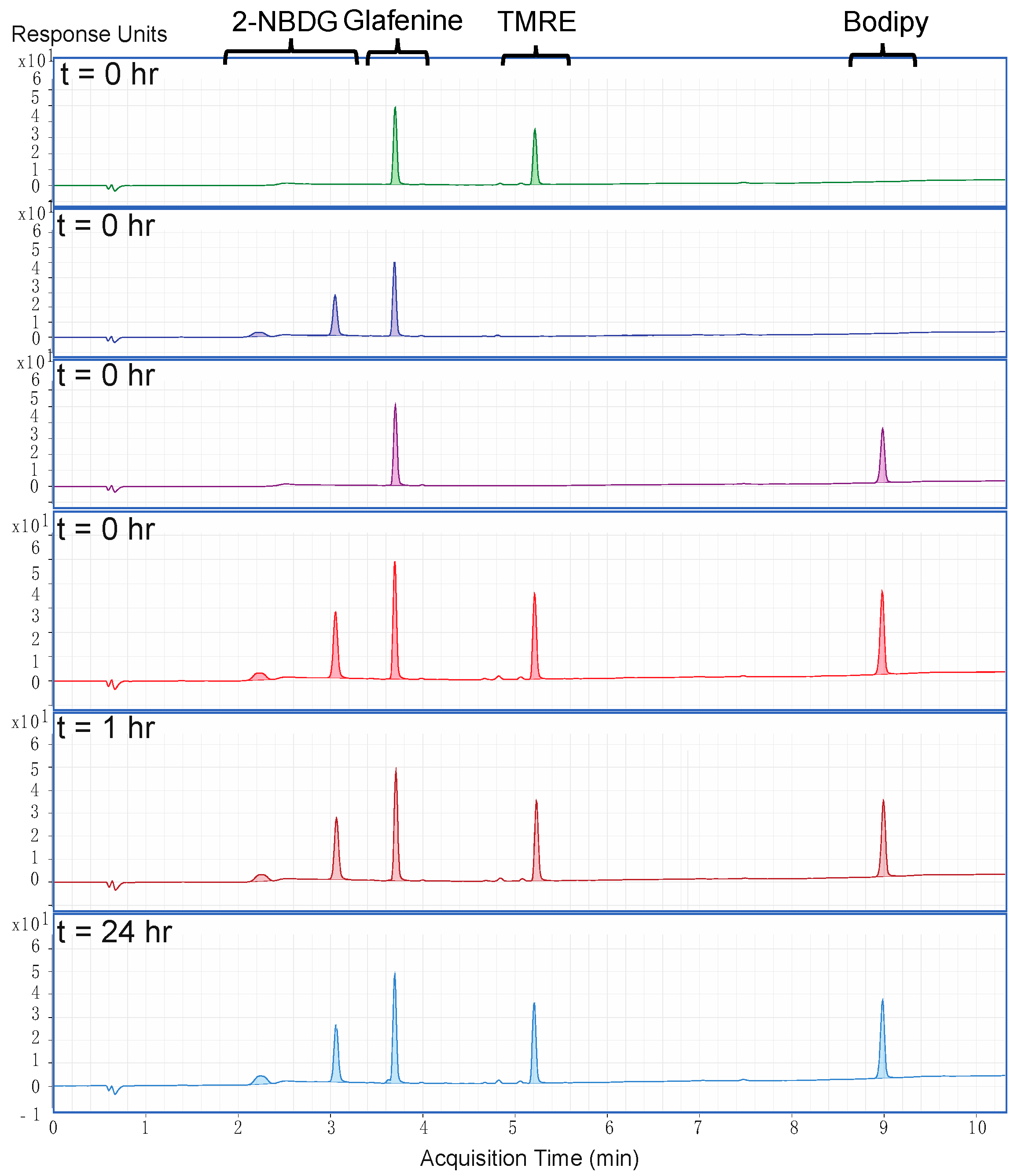
2.1. There Is No Chemical Crosstalk between TMRE and Bodipy FL C16
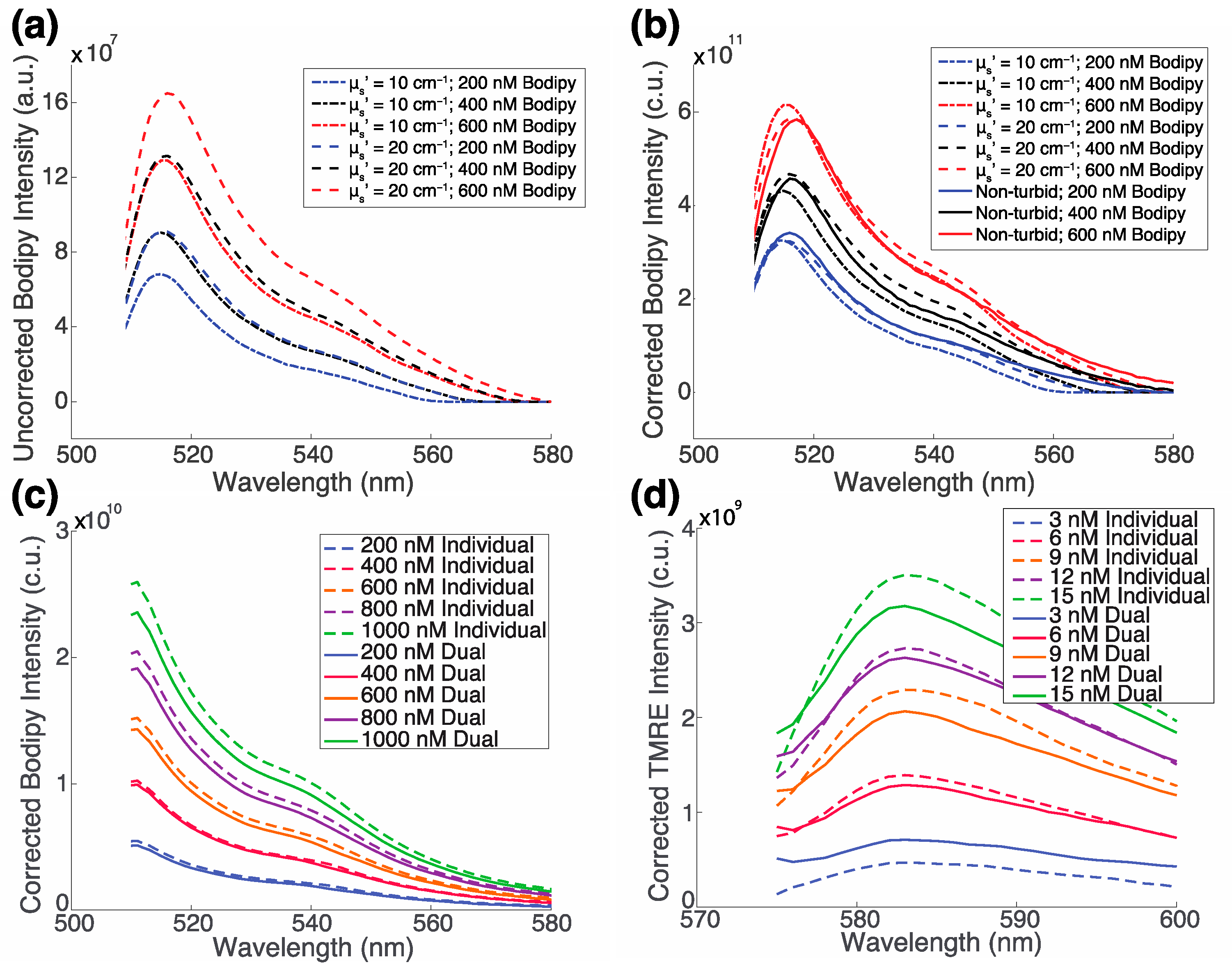
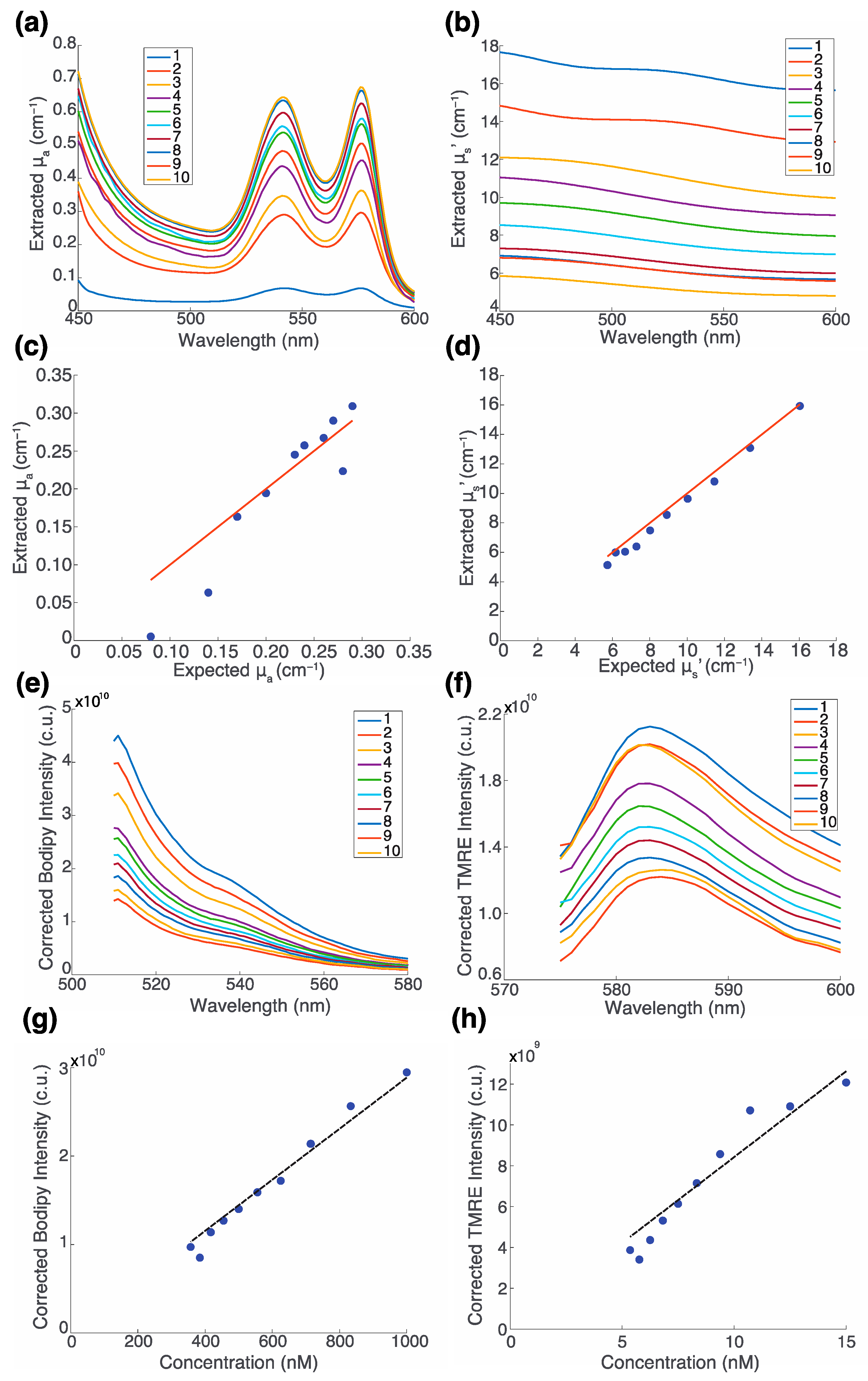
2.2. The Fluorescence of TMRE and Bodipy FL C16 along with Optical Properties Can Be Extracted from Turbid Phantoms with No Optical Crosstalk
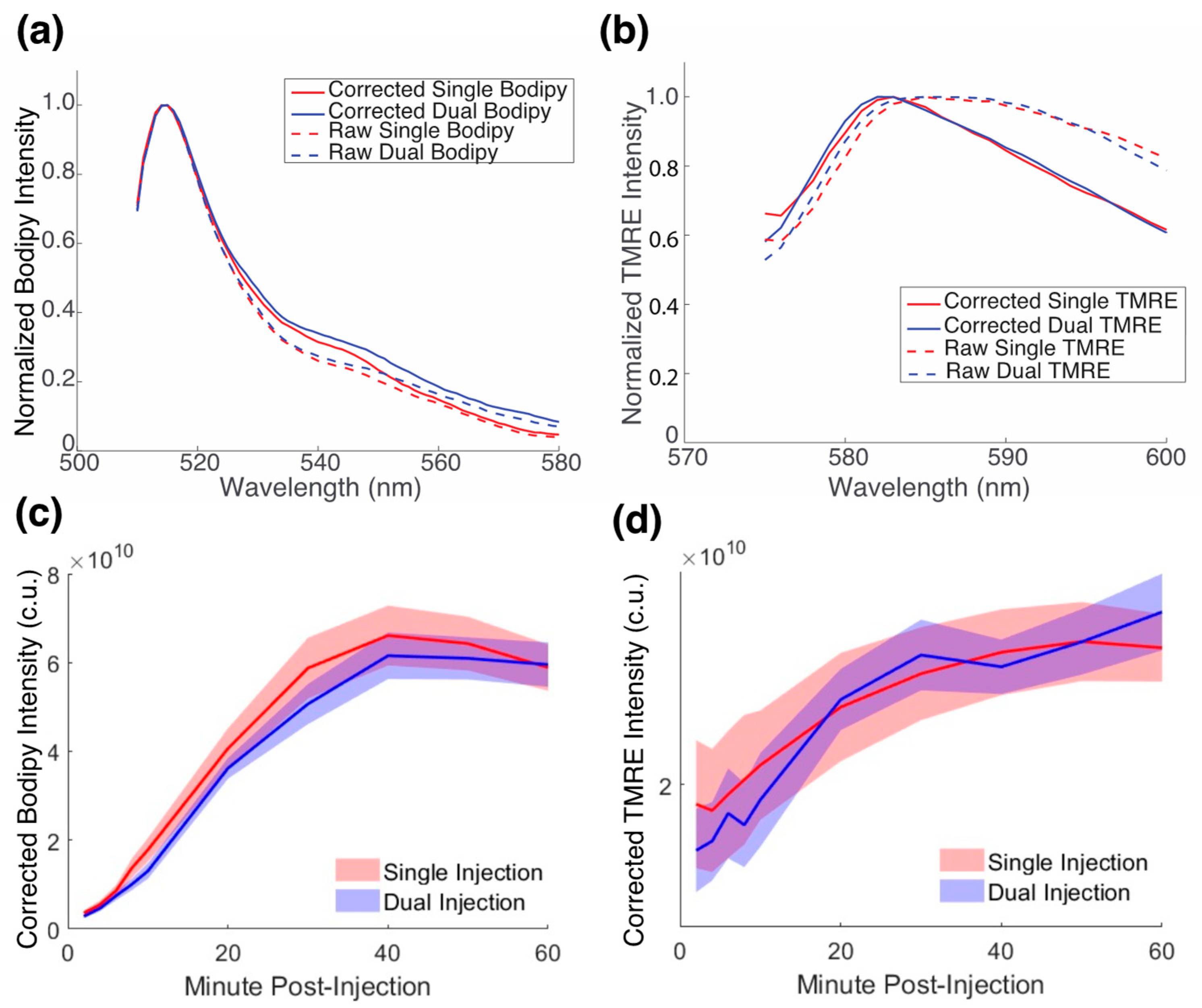
2.3. There Is No Biological Crosstalk between TMRE and Bodipy FL C16
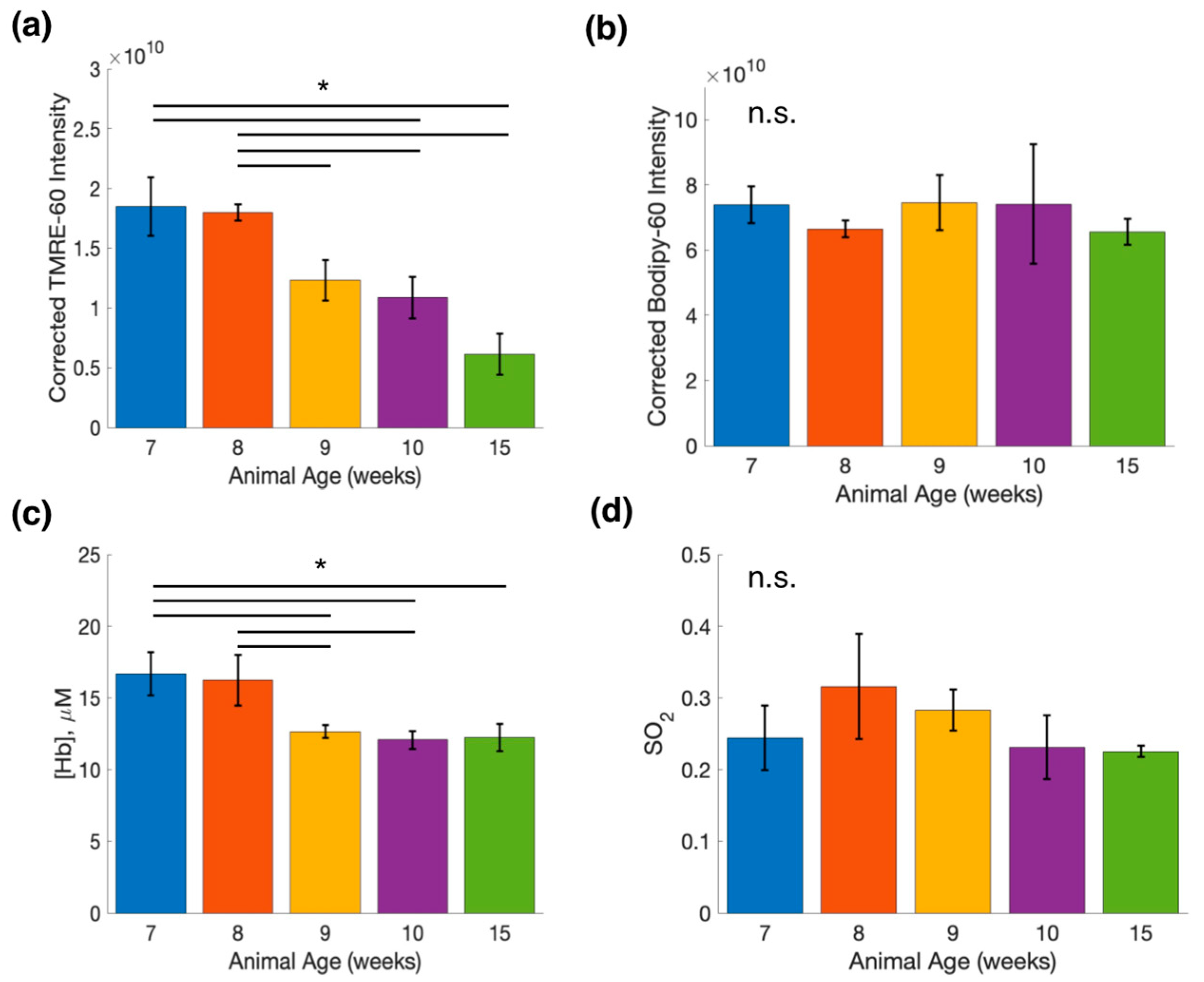
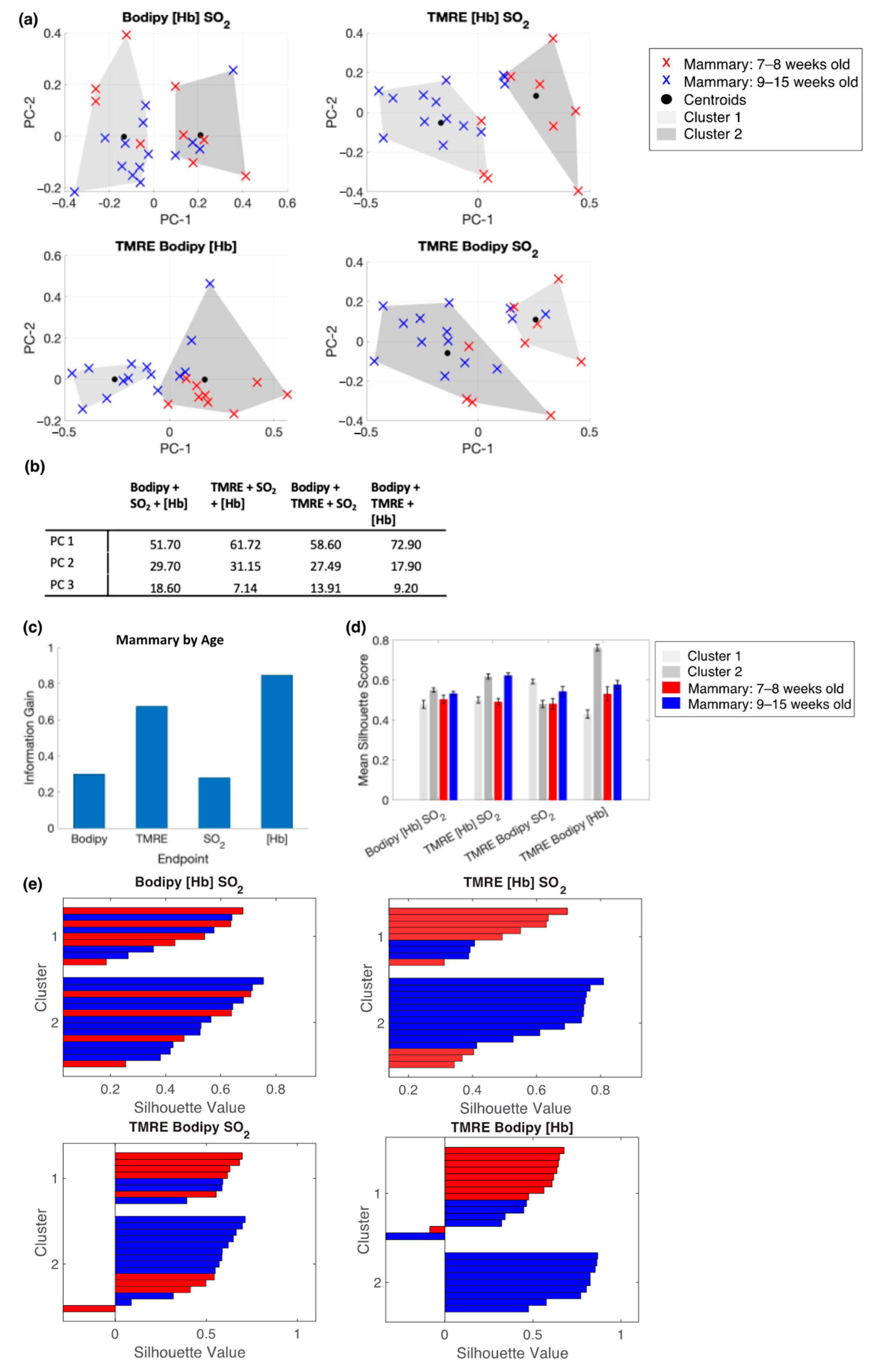
2.4. Bodipy FL C16-TMRE Spectroscopy Measurements Reveal Metabolic and Vascular Changes in Normal Mammary Glands throughout Development
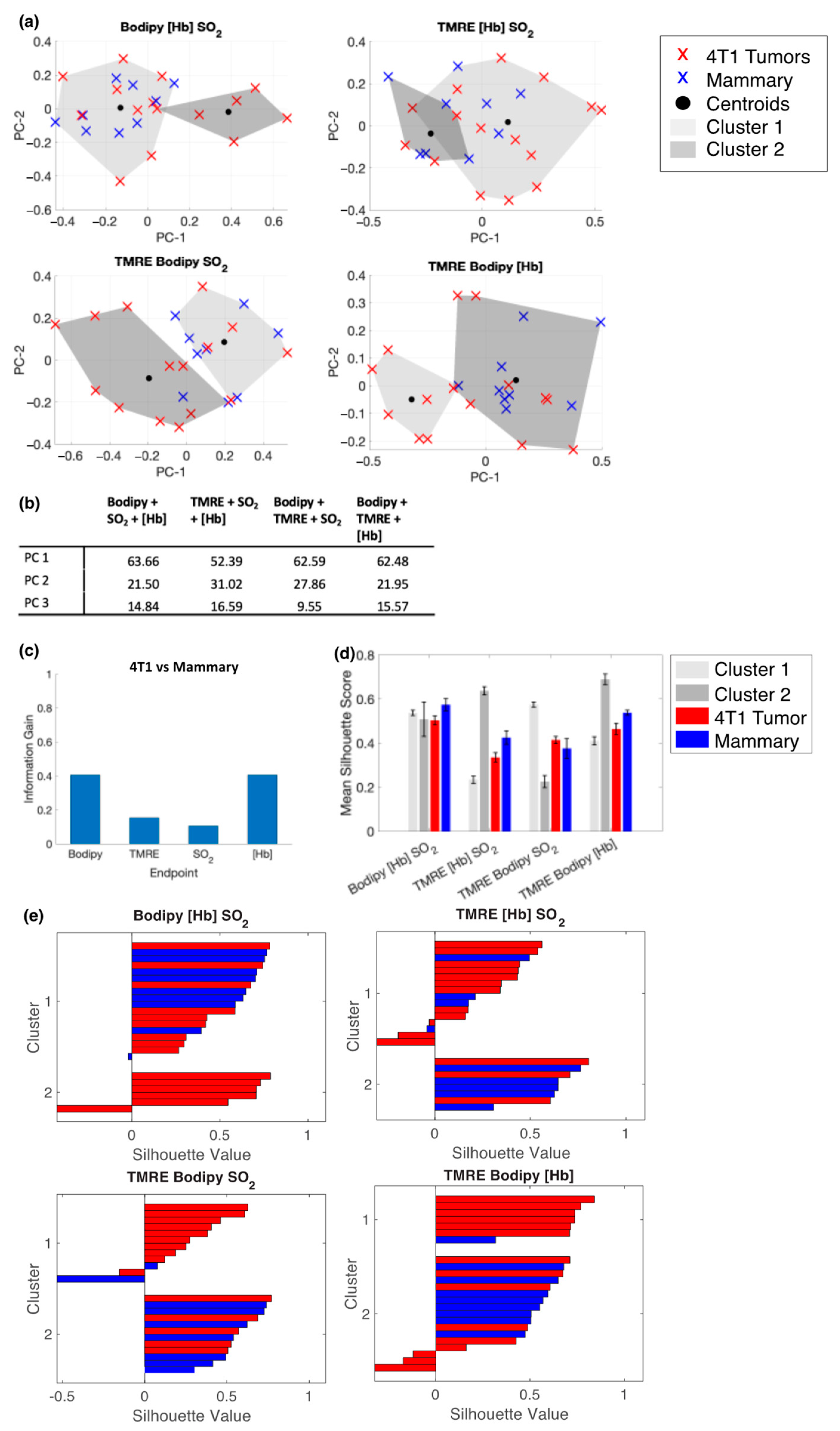
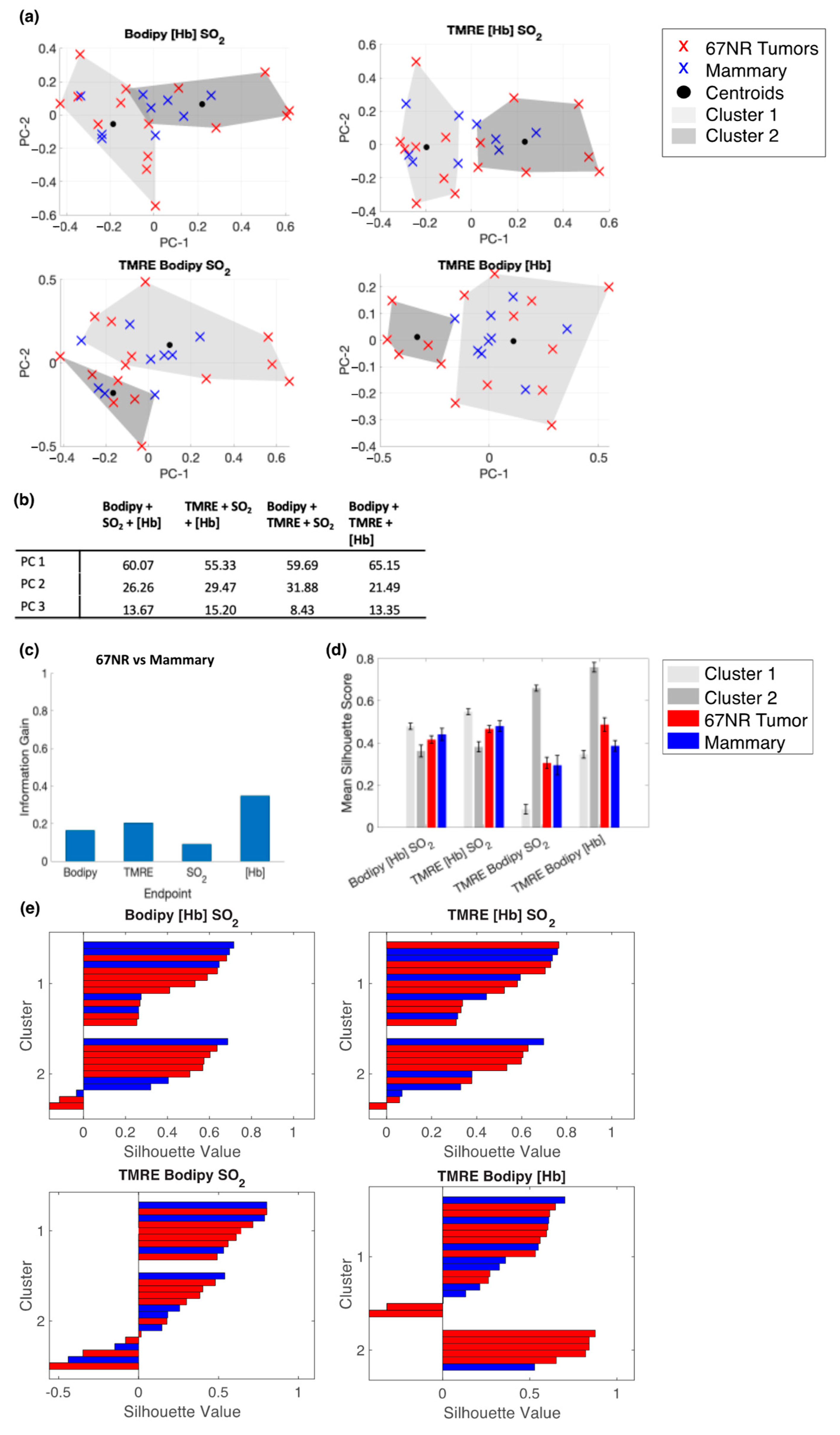
2.5. Bodipy FL C16 and TMRE Fluorescence Measurements Combined with Extracted Vasculature Parameters Show the Clustering of Tumors and Mammary Tissues in Age Matched Mice
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Liquid Chromatography-Mass Spectrometry of Fluorophore Samples
4.3. Optical Measurements
4.4. Inverse MC Models for Reflectance and Fluorescence
4.5. Tissue Phantoms
4.6. In Vivo Murine Breast Cancer Model Studies
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- de Heer, E.C.; Jalving, M.; Harris, A.L. HIFs, angiogenesis, and metabolism: Elusive enemies in breast cancer. J. Clin. Investig. 2020, 130, 5074–5087. [Google Scholar] [CrossRef] [PubMed]
- Kreuzaler, P.; Panina, Y.; Segal, J.; Yuneva, M. Adapt and conquer: Metabolic flexibility in cancer growth, invasion and evasion. Mol. Metab. 2020, 33, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, A.; Malvi, P.; Wajapeyee, N. Oncogene-directed alterations in cancer cell metabolism. Trends Cancer 2016, 2, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, M.M.; Chi, J.-T. Alternative fuels for cancer cells. Cancer J. 2015, 21, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Turdo, A.; Porcelli, G.; D’Accardo, C.; Di Franco, S.; Verona, F.; Forte, S.; Giuffrida, D.; Memeo, L.; Todaro, M.; Stassi, G. Metabolic Escape Routes of Cancer Stem Cells and Therapeutic Opportunities. Cancers 2020, 12, 1436. [Google Scholar] [CrossRef]
- Chen, E.I.; Hewel, J.; Krueger, J.S.; Tiraby, C.; Weber, M.R.; Kralli, A.; Becker, K.; Yates, J.R., 3rd; Felding-Habermann, B. Adaptation of energy metabolism in breast cancer brain metastases. Cancer Res. 2007, 67, 1472–1486. [Google Scholar] [CrossRef] [Green Version]
- Simões, R.V.; Serganova, I.S.; Kruchevsky, N.; Leftin, A.; Shestov, A.A.; Thaler, H.T.; Sukenick, G.; Locasale, J.W.; Blasberg, R.G.; Koutcher, J.A.; et al. Metabolic plasticity of metastatic breast cancer cells: Adaptation to changes in the microenvironment. Neoplasia 2015, 17, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Apple, S.; Zhao, H.; Song, J.; Lee, M.; Luo, W.; Wu, X.; Chung, D.; Pietras, R.J.; Chang, H.R. CD24 Expression and differential resistance to chemotherapy in triple-negative breast cancer. Oncotarget 2017, 8, 38294–38308. [Google Scholar] [CrossRef] [Green Version]
- Valcourt, J.R.; Lemons, J.M.; Haley, E.M.; Kojima, M.; Demuren, O.O.; Coller, H.A. Staying alive: Metabolic adaptations to quiescence. Cell Cycle 2012, 11, 1680–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sounni, N.E.; Cimino, J.; Blacher, S.; Primac, I.; Truong, A.; Mazzucchelli, G.; Paye, A.; Calligaris, D.; Debois, D.; De Tullio, P.; et al. Blocking lipid synthesis overcomes tumor regrowth and metastasis after antiangiogenic therapy withdrawal. Cell Metab. 2014, 20, 280–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havas, K.M.; Milchevskaya, V.; Radic, K.; Alladin, A.; Kafkia, E.; Garcia, M.; Stolte, J.; Klaus, B.; Rotmensz, N.; Gibson, T.J. Metabolic shifts in residual breast cancer drive tumor recurrence. J. Clin. Investig. 2017, 127, 2091–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarda, R.; Zhou, A.Y.; Kohnz, R.A.; Balakrishnan, S.; Mahieu, C.; Anderton, B.; Eyob, H.; Kajimura, S.; Tward, A.; Krings, G.; et al. Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer. Nat. Med. 2016, 22, 427–432. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Q.; Dong, C. Metabolic reprogramming in triple-negative breast cancer. Cancer Biol. Med. 2020, 17, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Madonna, M.C.; Duer, J.E.; Lee, J.V.; Williams, J.; Avsaroglu, B.; Zhu, C.; Deutsch, R.; Wang, R.; Crouch, B.T.; Hirschey, M.D.; et al. In Vivo Optical Metabolic Imaging of Long-Chain Fatty Acid Uptake in Orthotopic Models of Triple-Negative Breast Cancer. Cancers 2021, 13, 148. [Google Scholar] [CrossRef]
- Sirois, I.; Aguilar-Mahecha, A.; Lafleur, J.; Fowler, E.; Vu, V.; Scriver, M.; Buchanan, M.; Chabot, C.; Ramanathan, A.; Balachandran, B.; et al. A Unique Morphological Phenotype in Chemoresistant Triple-Negative Breast Cancer Reveals Metabolic Reprogramming and PLIN4 Expression as a Molecular Vulnerability. Mol. Cancer Res. 2019, 17, 2492. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Fahrmann, J.F.; Lee, H.; Li, Y.J.; Tripathi, S.C.; Yue, C.; Zhang, C.; Lifshitz, V.; Song, J.; Yuan, Y.; et al. JAK/STAT3-Regulated Fatty Acid β-Oxidation Is Critical for Breast Cancer Stem Cell Self-Renewal and Chemoresistance. Cell Metab. 2018, 27, 136–150.e135. [Google Scholar] [CrossRef] [Green Version]
- Camarda, R.; Williams, J.; Goga, A. In vivo Reprogramming of Cancer Metabolism by MYC. Front. Cell Dev. Biol. 2017, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Casciano, J.C.; Perry, C.; Cohen-Nowak, A.J.; Miller, K.D.; Voorde, J.V.; Zhang, Q.; Chalmers, S.; Sandison, M.E.; Liu, Q.; Hedley, A. MYC regulates fatty acid metabolism through a multigenic program in claudin-low triple negative breast cancer. Br. J. Cancer 2020, 122, 868–884. [Google Scholar] [CrossRef]
- Madonna, M.C.; Fox, D.B.; Crouch, B.T.; Lee, J.; Zhu, C.; Martinez, A.F.; Alvarez, J.V.; Ramanujam, N. Optical Imaging of Glucose Uptake and Mitochondrial Membrane Potential to Characterize Her2 Breast Tumor Metabolic Phenotypes. Mol. Cancer Res. 2019, 17, 1545–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Martin, H.L.; Crouch, B.T.; Martinez, A.F.; Li, M.; Palmer, G.M.; Dewhirst, M.W.; Ramanujam, N. Near-simultaneous quantification of glucose uptake, mitochondrial membrane potential, and vascular parameters in murine flank tumors using quantitative diffuse reflectance and fluorescence spectroscopy. Biomed. Opt. Express 2018, 9, 3399–3412. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Yu, Y.; Folick, A.; Currie, E.; Farese, R.V., Jr.; Tsai, T.-H.; Xie, X.S.; Wang, M.C. In vivo metabolic fingerprinting of neutral lipids with hyperspectral stimulated Raman scattering microscopy. J. Am. Chem. Soc. 2014, 136, 8820–8828. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Shen, Y.; Xu, F.; Hu, F.; Harrington, J.K.; Targoff, K.L.; Min, W. Imaging complex protein metabolism in live organisms by stimulated Raman scattering microscopy with isotope labeling. ACS Chem. Biol. 2015, 10, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, R.; Zhang, L.; Shi, L.; Shen, Y.; Hu, F.; Zeng, C.; Min, W. Two-color vibrational imaging of glucose metabolism using stimulated Raman scattering. Chem. Commun. 2018, 54, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Pouli, D.; Alonzo, C.A.; Varone, A.; Karaliota, S.; Quinn, K.P.; Münger, K.; Karalis, K.P.; Georgakoudi, I. Mapping metabolic changes by noninvasive, multiparametric, high-resolution imaging using endogenous contrast. Sci. Adv. 2018, 4, eaap9302. [Google Scholar] [CrossRef] [Green Version]
- Paidi, S.K.; Diaz, P.M.; Dadgar, S.; Jenkins, S.V.; Quick, C.M.; Griffin, R.J.; Dings, R.P.M.; Rajaram, N.; Barman, I. Label-Free Raman Spectroscopy Reveals Signatures of Radiation Resistance in the Tumor Microenvironment. Cancer Res. 2019, 79, 2054–2064. [Google Scholar] [CrossRef] [Green Version]
- Richert, M.M.; Schwertfeger, K.L.; Ryder, J.W.; Anderson, S.M. An atlas of mouse mammary gland development. J. Mammary Gland. Biol. Neoplasia 2000, 5, 227–241. [Google Scholar] [CrossRef]
- Colleluori, G.; Perugini, J.; Barbatelli, G.; Cinti, S. Mammary gland adipocytes in lactation cycle, obesity and breast cancer. Rev. Endocr. Metab. Disord. 2021, 22, 241–255. [Google Scholar] [CrossRef]
- Martinez, A.F.; McCachren, S.S., 3rd; Lee, M.; Murphy, H.A.; Zhu, C.; Crouch, B.T.; Martin, H.L.; Erkanli, A.; Rajaram, N.; Ashcraft, K.A.; et al. Metaboloptics: Visualization of the tumor functional landscape via metabolic and vascular imaging. Sci. Rep. 2018, 8, 4171. [Google Scholar] [CrossRef]
- Zhu, C.; Li, M.; Vincent, T.; Martin, H.L.; Crouch, B.T.; Martinez, A.F.; Madonna, M.C.; Palmer, G.M.; Dewhirst, M.W.; Ramanujam, N. Simultaneous in vivo optical quantification of key metabolic and vascular endpoints reveals tumor metabolic diversity in murine breast tumor models. J. Biophotonics 2019, 12, e201800372. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Martinez, A.F.; Martin, H.L.; Li, M.; Crouch, B.T.; Carlson, D.A.; Haystead, T.A.; Ramanujam, N. Near-simultaneous intravital microscopy of glucose uptake and mitochondrial membrane potential, key endpoints that reflect major metabolic axes in cancer. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pranzini, E.; Raugei, G.; Taddei, M.L. Metabolic Features of Tumor Dormancy: Possible Therapeutic Strategies. Cancers 2022, 14, 547. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Park, J.H.; Jung, K.H.; Levine, H.; Kaipparettu, B.A. Elucidating the metabolic plasticity of cancer: Mitochondrial reprogramming and hybrid metabolic states. Cells 2018, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, N.; Reesor, A.F.; Mulvey, C.S.; Frees, A.E.; Ramanujam, N. Non-invasive, simultaneous quantification of vascular oxygenation and glucose uptake in tissue. PLoS ONE 2015, 10, e0117132. [Google Scholar] [CrossRef]
- Bender, J.; Vishwanath, K.; Moore, L.; Brown, J.; Chang, V.; Palmer, G.; Ramanujam, N. A Robust Monte Carlo Model for the Extraction of Biological Absorption and Scattering In Vivo. IEEE Trans. Bio-Med. Eng. 2009, 56, 960–968. [Google Scholar] [CrossRef]
- Liu, C.; Rajaram, N.; Vishwanath, K.; Jiang, T.; Palmer, G.M.; Ramanujam, N. Experimental validation of an inverse fluorescence Monte Carlo model to extract concentrations of metabolically relevant fluorophores from turbid phantoms and a murine tumor model. J. Biomed. Opt. 2012, 17, 077012. [Google Scholar] [CrossRef] [Green Version]
- Palmer, G.M.; Ramanujam, N. Monte-Carlo-based model for the extraction of intrinsic fluorescence from turbid media. J. Biomed. Opt. 2008, 13, 024017. [Google Scholar] [CrossRef]
- Palmer, G.M.; Ramanujam, N. Monte Carlo-based inverse model for calculating tissue optical properties. Part I: Theory and validation on synthetic phantoms. Appl. Opt. 2006, 45, 1062–1071. [Google Scholar]
- Fu, Q.; Sun, W. Mie theory for light scattering by a spherical particle in an absorbing medium. Appl. Opt. 2001, 40, 1354–1361. [Google Scholar] [CrossRef]
- Thumser, A.E.; Storch, J. Characterization of a BODIPY-labeled fluorescent fatty acid analogue. Binding to fatty acid-binding proteins, intracellular localization, and metabolism. Mol. Cell. Biochem. 2007, 299, 67–73. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phantom | μa 1 (cm−1) | μs′ 2 (cm−1) | [Hb] (μM) | [Bodipy] (nM) | [TMRE] (nM) |
|---|---|---|---|---|---|
| 1 | 0.14 | 16.72 | 15.09 | 1000 | 15 |
| 2 | 0.23 | 13.93 | 25.15 | 833.33 | 12.5 |
| 3 | 0.29 | 11.94 | 32.33 | 714.29 | 10.71 |
| 4 | 0.34 | 10.45 | 37.72 | 625 | 9.38 |
| 5 | 0.38 | 9.29 | 41.91 | 555.56 | 8.33 |
| 6 | 0.41 | 8.36 | 45.26 | 500 | 7.5 |
| 7 | 0.44 | 7.60 | 48.01 | 454.55 | 6.82 |
| 8 | 0.46 | 6.97 | 50.29 | 416.67 | 6.25 |
| 9 | 0.48 | 6.43 | 52.23 | 384.62 | 5.77 |
| 10 | 0.49 | 5.97 | 53.89 | 357.14 | 5.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deutsch, R.J.; D’Agostino, V.W.; Sunassee, E.D.; Kwan, M.; Madonna, M.C.; Palmer, G.; Crouch, B.T.; Ramanujam, N. A Spectroscopic Technique to Simultaneously Characterize Fatty Acid Uptake, Mitochondrial Activity, Vascularity, and Oxygen Saturation for Longitudinal Studies In Vivo. Metabolites 2022, 12, 369. https://doi.org/10.3390/metabo12050369
Deutsch RJ, D’Agostino VW, Sunassee ED, Kwan M, Madonna MC, Palmer G, Crouch BT, Ramanujam N. A Spectroscopic Technique to Simultaneously Characterize Fatty Acid Uptake, Mitochondrial Activity, Vascularity, and Oxygen Saturation for Longitudinal Studies In Vivo. Metabolites. 2022; 12(5):369. https://doi.org/10.3390/metabo12050369
Chicago/Turabian StyleDeutsch, Riley J., Victoria W. D’Agostino, Enakshi D. Sunassee, Michelle Kwan, Megan C. Madonna, Gregory Palmer, Brian T. Crouch, and Nimmi Ramanujam. 2022. "A Spectroscopic Technique to Simultaneously Characterize Fatty Acid Uptake, Mitochondrial Activity, Vascularity, and Oxygen Saturation for Longitudinal Studies In Vivo" Metabolites 12, no. 5: 369. https://doi.org/10.3390/metabo12050369
APA StyleDeutsch, R. J., D’Agostino, V. W., Sunassee, E. D., Kwan, M., Madonna, M. C., Palmer, G., Crouch, B. T., & Ramanujam, N. (2022). A Spectroscopic Technique to Simultaneously Characterize Fatty Acid Uptake, Mitochondrial Activity, Vascularity, and Oxygen Saturation for Longitudinal Studies In Vivo. Metabolites, 12(5), 369. https://doi.org/10.3390/metabo12050369