
Effects of Dietary L-TRP on Immunity, Antioxidant Capacity and Intestinal Microbiota of the Chinese Mitten Crab (Eriocheir Sinensis) in Pond Culture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Design and Animal Culturing
2.3. Sample Collection
2.4. Analysis of Immune and Antioxidant Capacity
2.5. Analysis of Intestinal Microbiota
2.5.1. Total DNA Extraction, 16S rRNA Amplification, Purification and Quantification
2.5.2. Illumina MiSeq Sequencing
2.5.3. Biodiversity Analysis
2.6. Statistical Analysis
3. Results
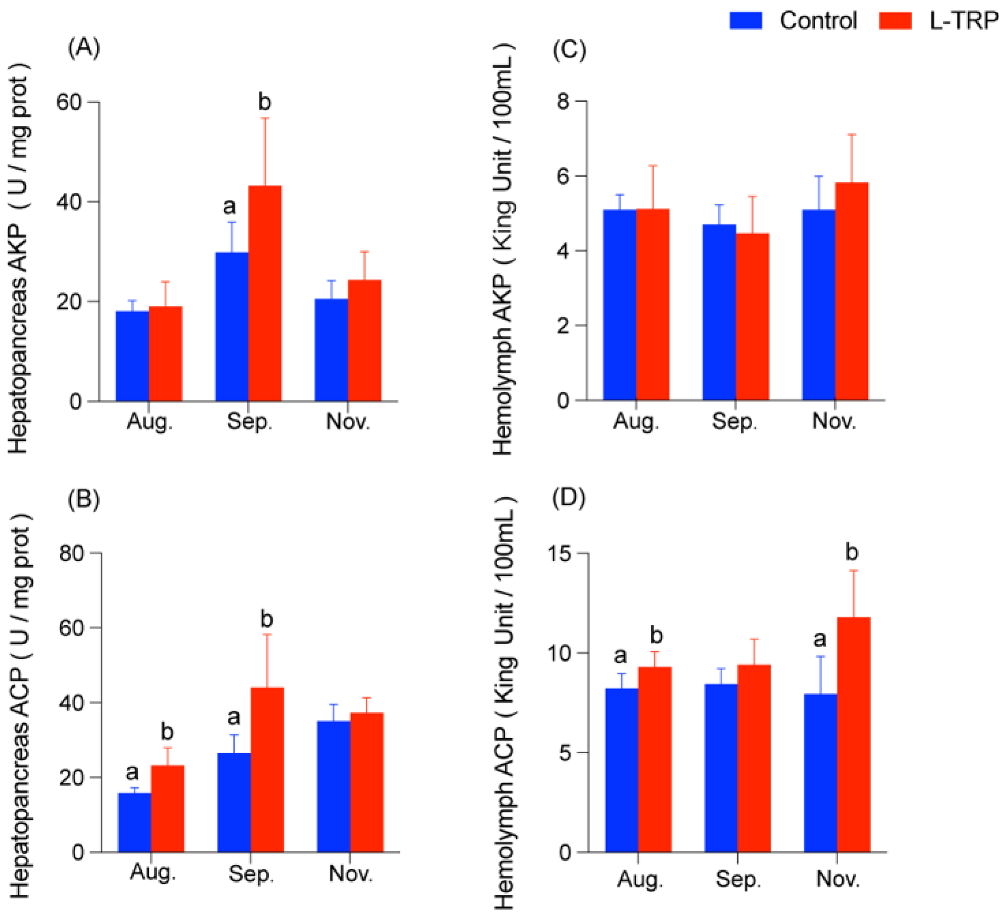
3.1. Immunity
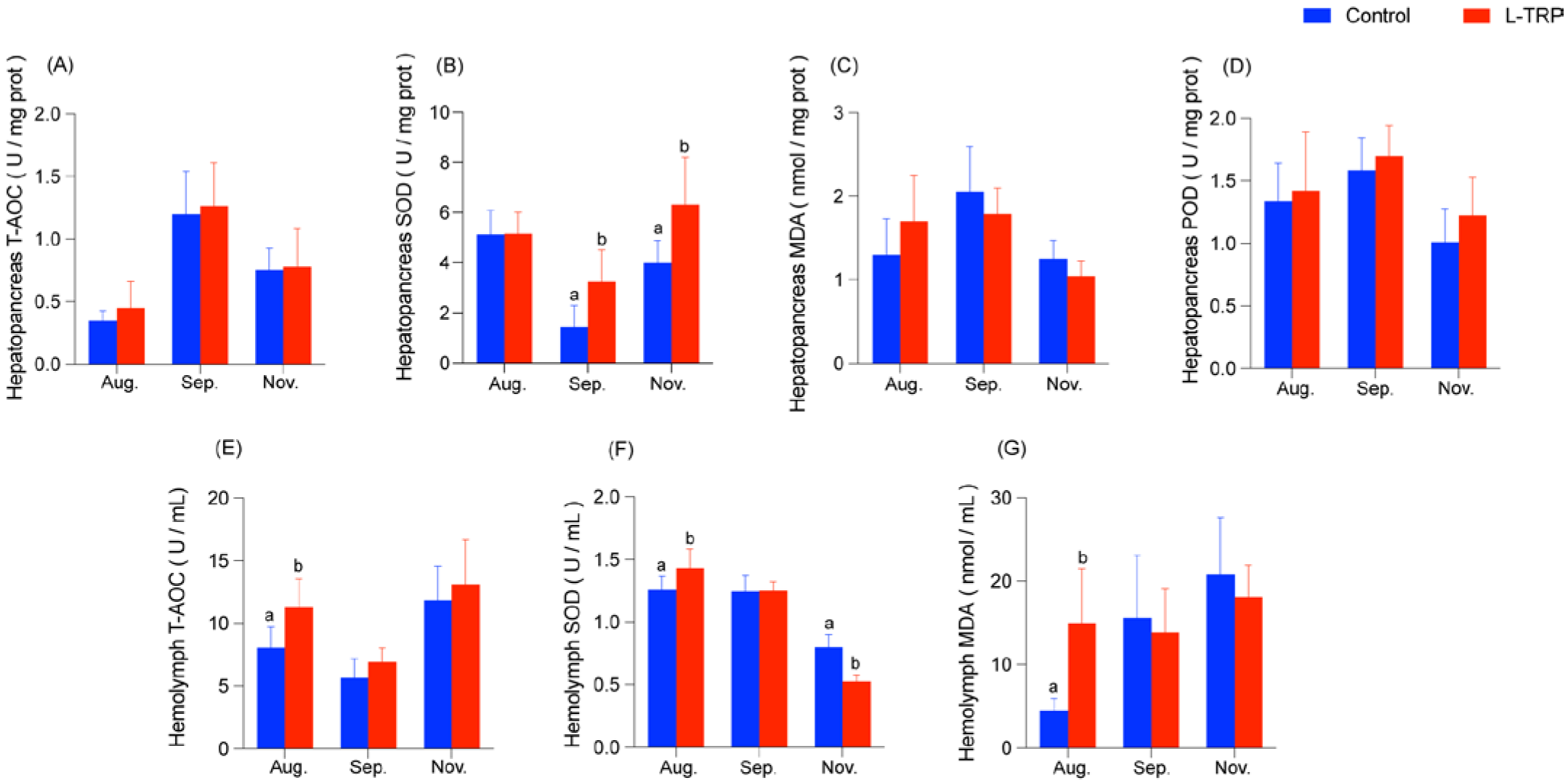
3.2. Antioxidant Capacity
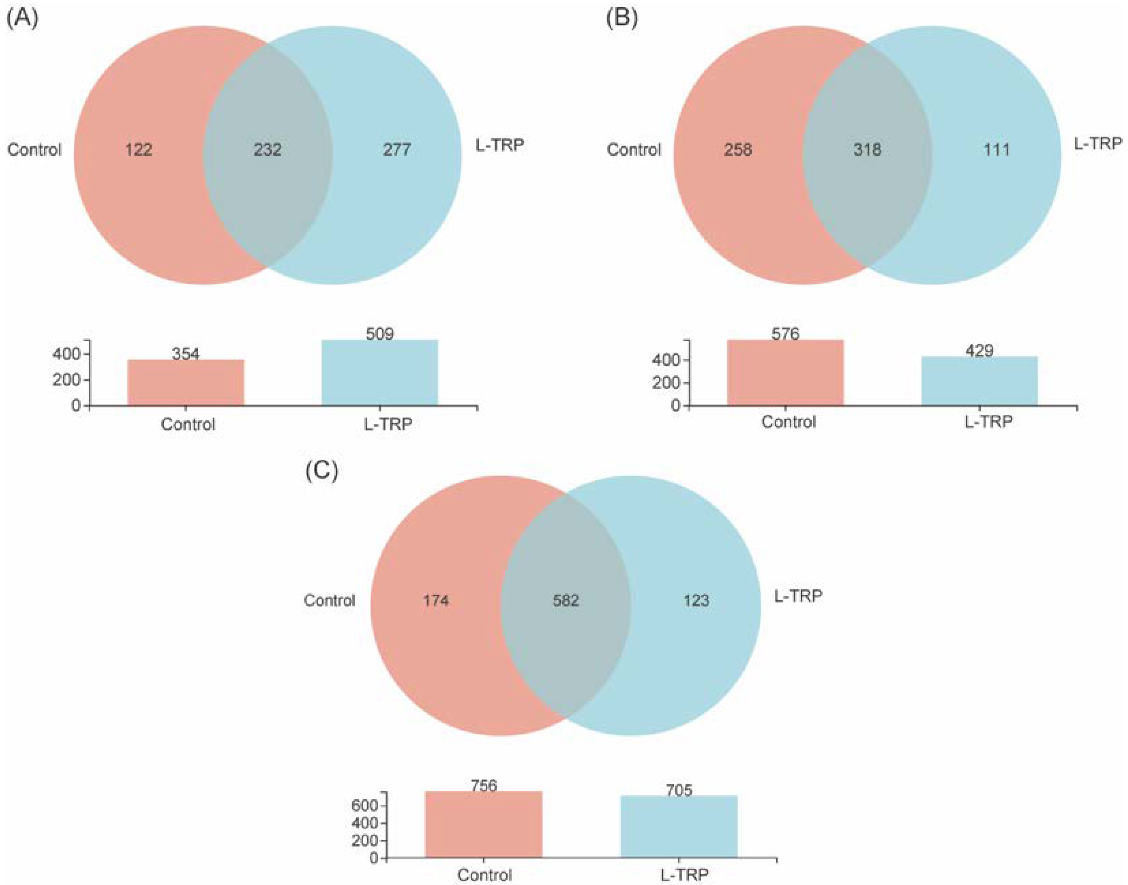
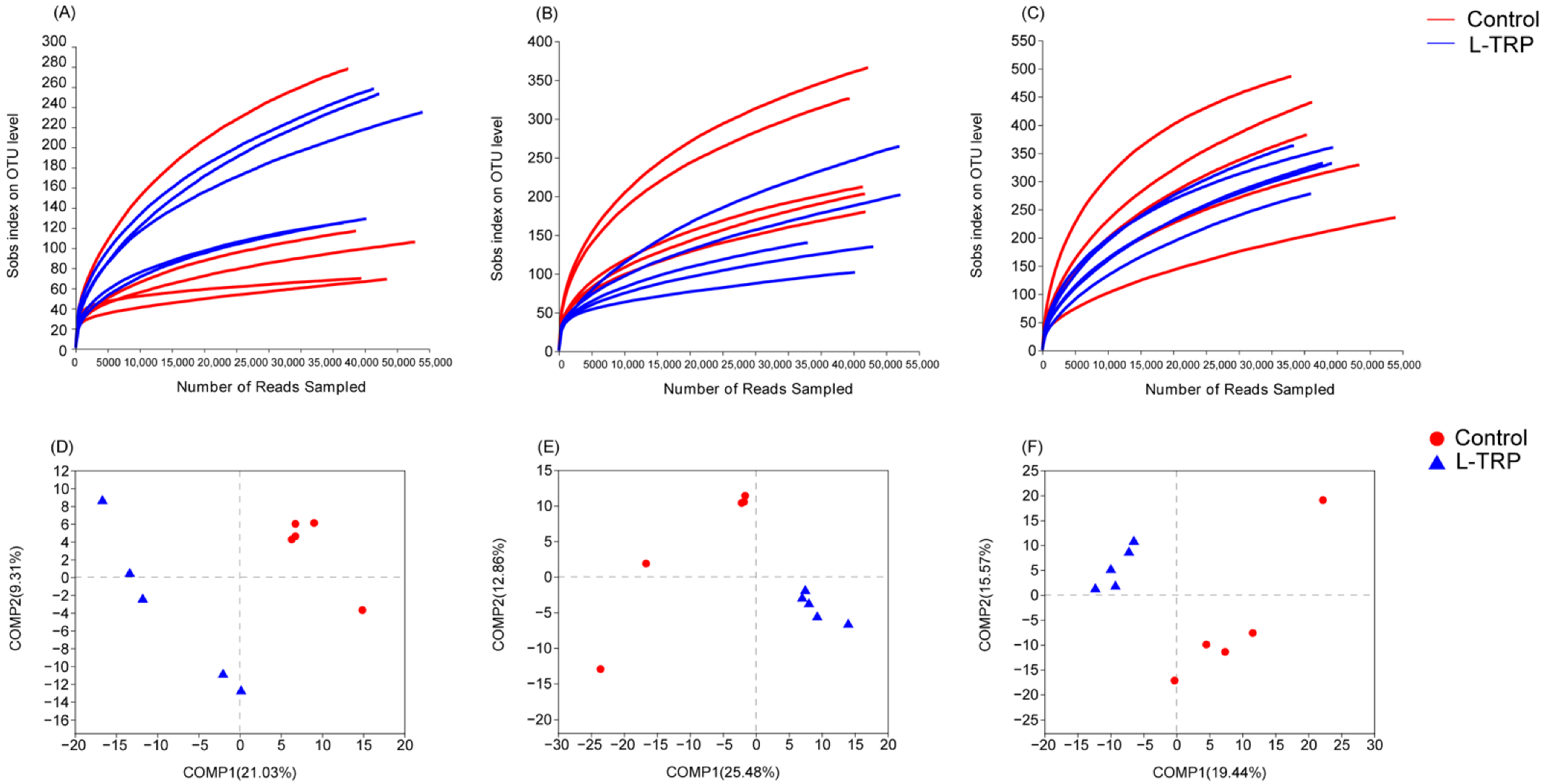
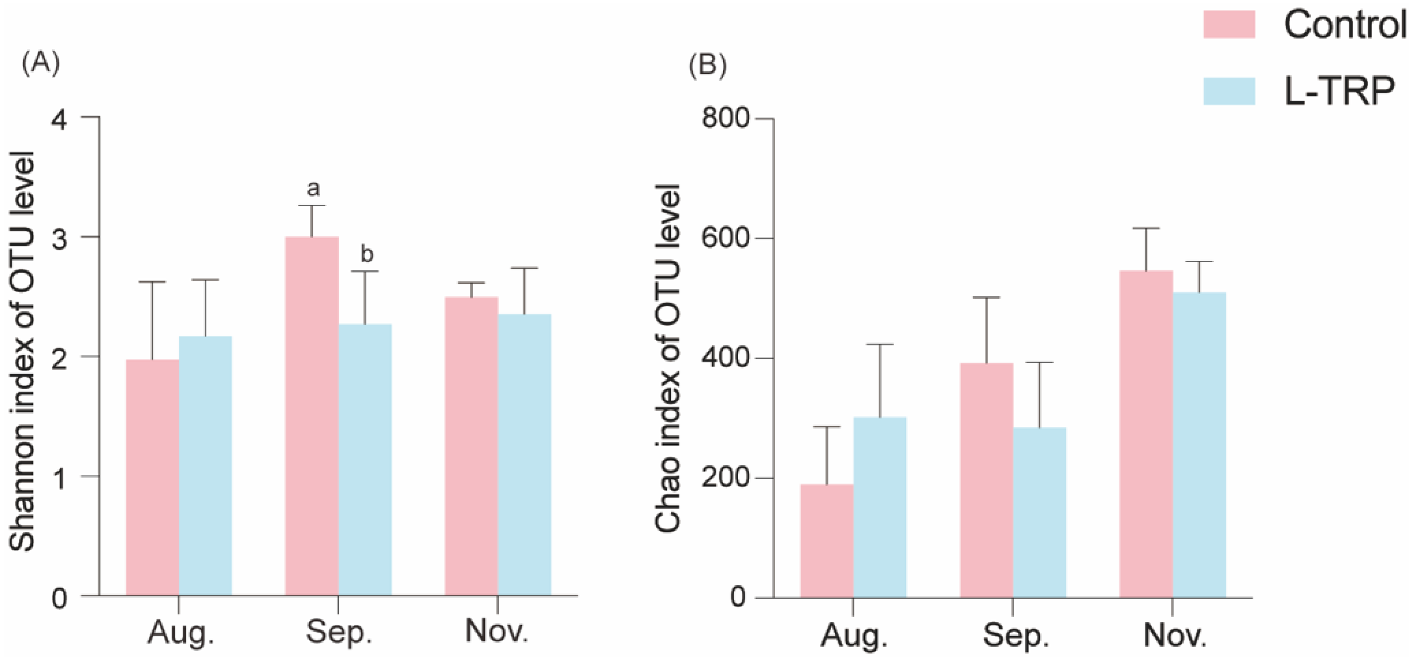
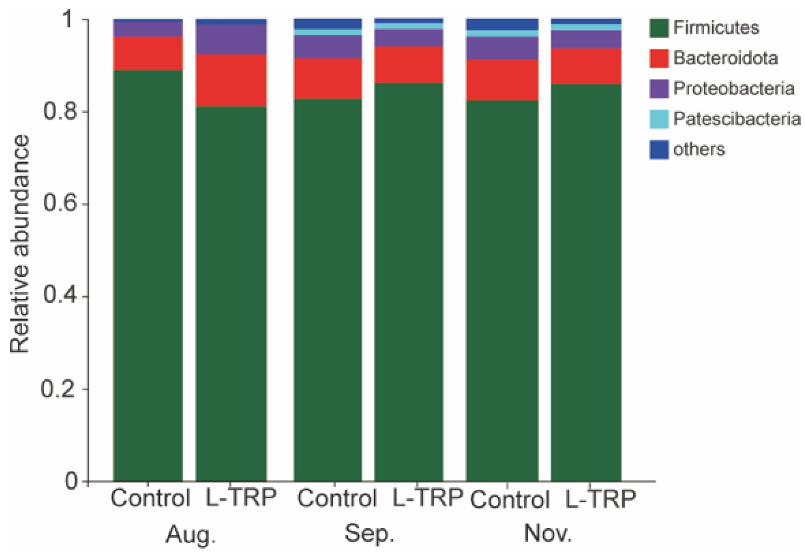
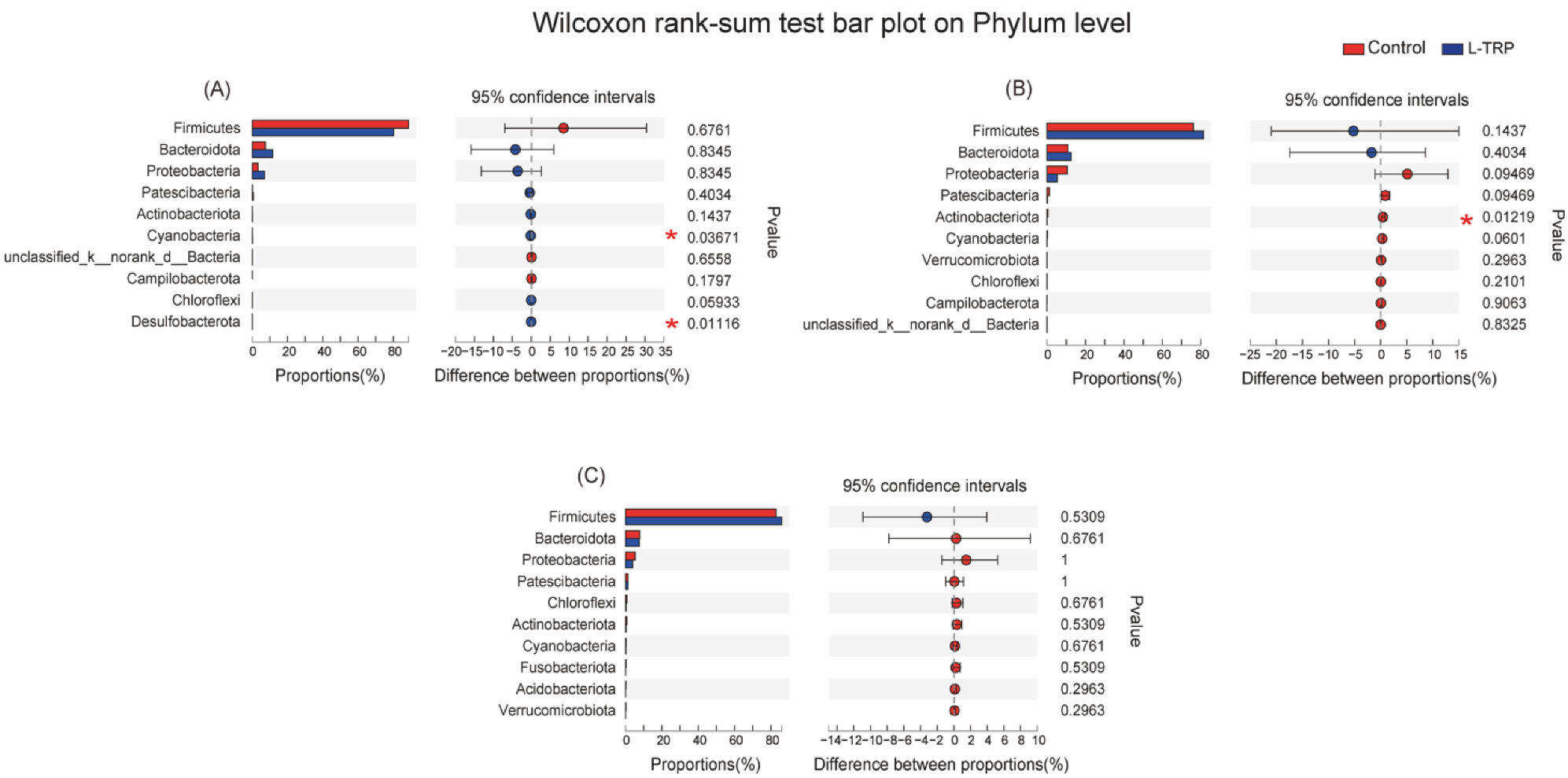
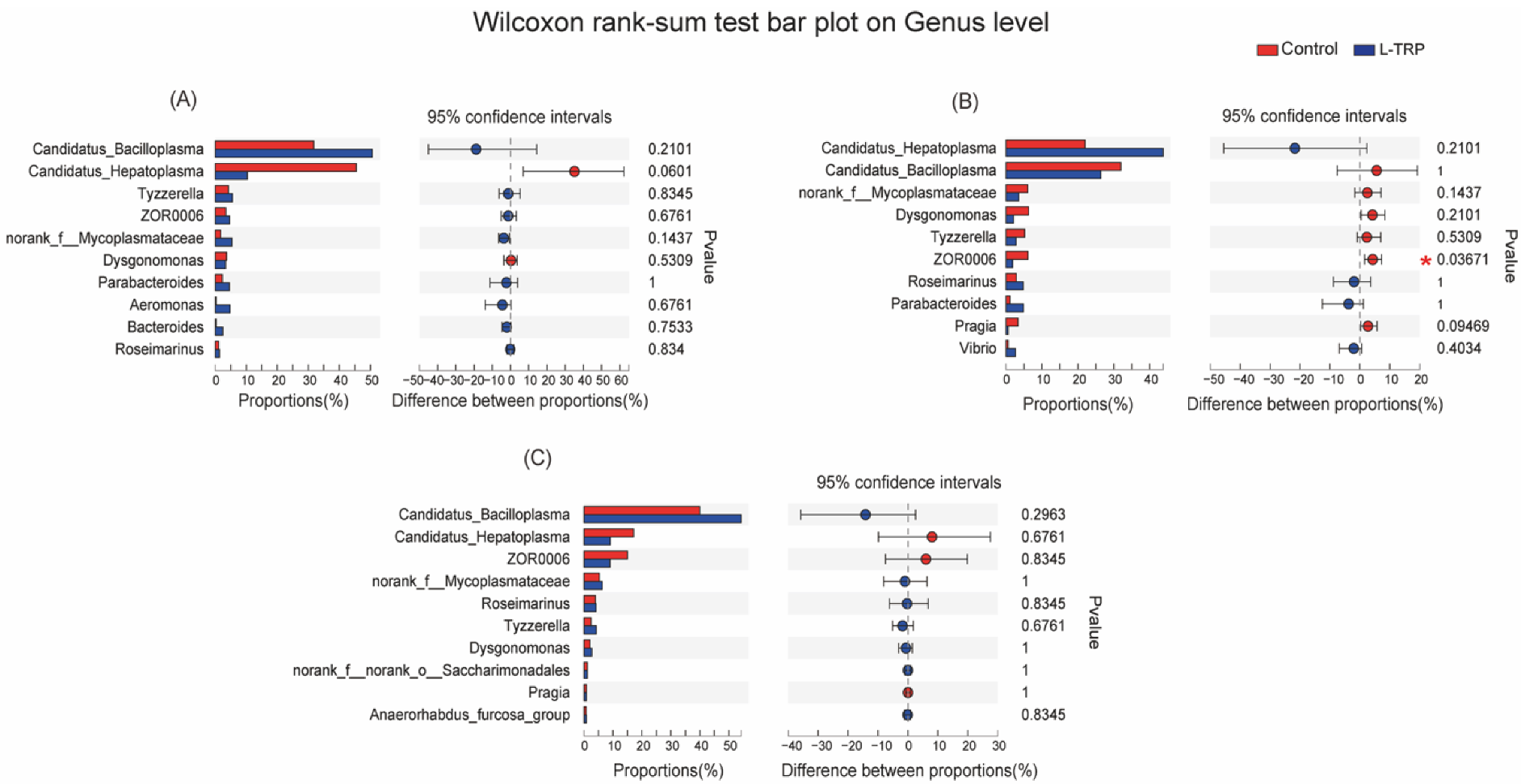
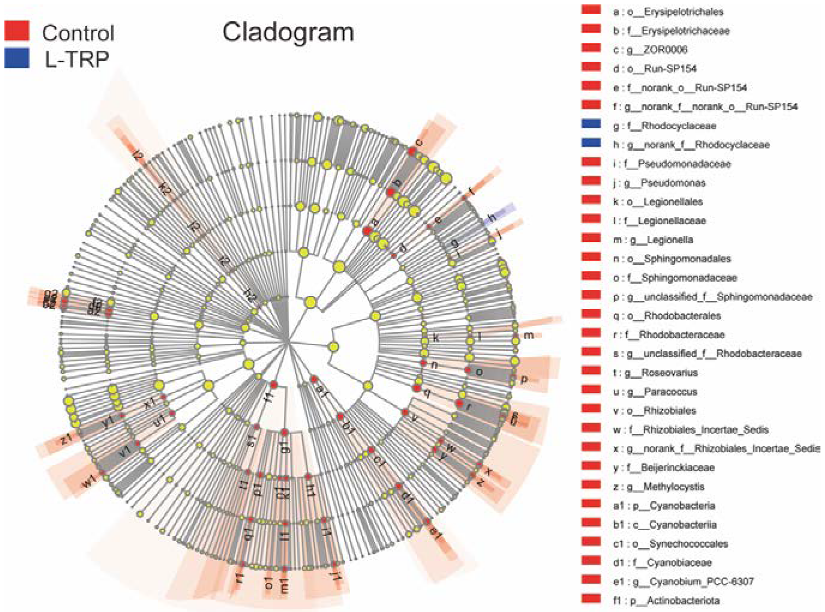
3.3. Intestinal Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qin, Z.D.; Babu, V.S.; Wan, Q.Y.; Muhammad, A.; Li, J.; Lan, J.F.; Lin, L. Antibacterial activity of hemocyanin from red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2018, 75, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Bacterial diseases of crabs: A review. J. Invertebrate Pathol. 2011, 106, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Shi, Z.; Zhang, J.; Bonami, J.R. Purification and characterization of a new reovirus from the Chinese mitten crab, Eriocheir sinensis. J. Fish Dis. 2004, 27, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, J. Ultrastructural study on a novel microsporidian, Endoreticulatus eriocheir sp. nov. (Microsporidia, Encephalitozoonidae), parasite of Chinese mitten crab, Eriocheir sinensis (Crustacea, Decapoda). J. Invertebr. Pathol. 2007, 94, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Dong, S.; Wang, F.; Tian, X.; Gao, Q. Effects of l-tryptophan on the growth, intestinal enzyme activities and non-specific immune response of sea cucumber (Apostichopus japonicus Selenka) exposed to crowding stress. Fish Shellfish Immunol. 2018, 75, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Pal, A.K.; Sahu, N.P.; Jha, A.K.; Kumar, N.; Christina, L.; Priya, P. Dietary L-Tryptophan potentiates non-specific immunity in Labeo rohita fingerlings reared under elevated temperature. J. Therm. Biol. 2018, 74, 55–62. [Google Scholar] [CrossRef]
- Huang, Z.S.; Aweya, J.J.; Zhu, C.H.; Tran, N.T.; Hong, Y.J.; Li, S.K.; Yao, D.F.; Zhang, Y.L. Modulation of Crustacean Innate Immune Response by Amino Acids and Their Metabolites: Inferences from Other Species. Front. Immunol. 2020, 11, 574721. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Domingues, A.; Fernandez-Boo, S.; Dias, J.; Conceio, L.; Costas, B. Dietary tryptophan deficiency and its supplementation compromises inflammatory mechanisms and disease resistance in a teleost fish. Sci. Rep. 2019, 9, 7689. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, R.; Machado, M.; Moura, J.; Martos-Sitcha, J.A.; Costas, B. Dietary tryptophan supplementation modulates imune status and disease resistance in senegalese sole, Solea senegalensis, reared at low or high density. In Proceedings of the International Meeting on Marine Research, Lausanne, Switzerland, 14 July 2016. [Google Scholar]
- Liu, F.; Qu, Y.-K.; Geng, C.; Wang, A.-M.; Zhang, J.-H.; Chen, K.-J.; Liu, B.; Tian, H.-Y.; Yang, W.-P.; Yu, Y.-B. Effects of hesperidin on the growth performance, antioxidant capacity, immune responses and disease resistance of red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 99, 154–166. [Google Scholar] [CrossRef]
- Sunish, K.S.; Biji, M.; Rosamma, P.; Sudheer, N.S.; Sreedharan, K.; Mohandas, A.; Singh, I.S.B. Marine actinomycetes Nocardiopsis alba MCCB 110 has immunomodulatory property in the tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2020, 102, 125–132. [Google Scholar] [CrossRef]
- Yang, X.; Xu, M.; Huang, G.; Zhang, C.; Pang, Y.; Cheng, Y. Effect of dietary L-tryptophan on the survival, immune response and gut microbiota of the Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2019, 84, 1007–1017. [Google Scholar] [CrossRef]
- Fanshuang, K.; Yanhong, Z.; Haojie, Y.; Xiaoyu, W.; Abouel Azm, F.R.; Junfa, Y.; Qingsong, T. Effect of dietary vitamin C on the growth performance, nonspecific immunity and antioxidant ability of red swamp crayfish (Procambarus clarkii). Aquaculture 2021, 541, 736785. [Google Scholar] [CrossRef]
- Frías-Espericueta, M.G.; Bautista-Covarrubias, J.C.; Osuna-Martínez, C.C.; Delgado-Alvarez, C.; Bojórquez, C.; Aguilar-Juárez, M.; Roos-Muñoz, S.; Osuna-López, I.; Páez-Osuna, F. Metals and oxidative stress in aquatic decapod crustaceans: A review with special reference to shrimp and crabs. Aquat. Toxicol. 2022, 242, 106024. [Google Scholar] [CrossRef]
- Hou, S.; Li, J.; Huang, J.; Cheng, Y. Effects of dietary phospholipid and cholesterol levels on antioxidant capacity, nonspecial immune response and intestinal microflora of juvenile female crayfish, Procambarus clarkii. Aquacult. Rep. 2022, 25, 101245. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Li, J.; Ji, F.; Si, Q.; Jiang, Q. The effect of a polystyrene nanoplastic on the intestinal microbes and oxidative stress defense of the freshwater crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Perez-Jimenez, A.; Costas, B.; Azeredo, R.; Gesto, M. Physiological roles of tryptophan in teleosts: Current knowledge and perspectives for future studies. Rev. Aquac. 2019, 11, 3–24. [Google Scholar] [CrossRef] [Green Version]
- Ji, K.; Liang, H.; Ren, M.; Ge, X.; Liu, B.; Xi, B.; Pan, L.; Yu, H. Effects of dietary tryptophan levels on antioxidant status and immunity for juvenile blunt snout bream (Megalobrama amblycephala) involved in Nrf2 and TOR signaling pathway. Fish Shellfish Immunol. 2019, 93, 474–483. [Google Scholar] [CrossRef]
- Morteza Hoseini, S.; Morteza, Y.; Taheri Mirghaed, A.; Ahamad Paray, B.; Hossein Hoseinifar, S.; Van Doan, H. Effects of rearing density and dietary tryptophan supplementation on intestinal immune and antioxidant responses in rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 528, 735537. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, X.-Y.; Xu, S.-X.; Xie, J.-Y.; Xiang, K.-W.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, J. Dietary tryptophan affects growth performance, digestive and absorptive enzyme activities, intestinal antioxidant capacity, and appetite and GH-IGF axis-related gene expression of hybrid catfish (Pelteobagrus vachelli female × Leiocassis longirostris male). Fish Physiol. Biochem. 2019, 45, 1627–1647. [Google Scholar] [CrossRef]
- Mandiki, S.N.M.; Redivo, B.; Baekelandt, S.; Douxfils, J.; Lund, I.; Höglund, E.; Kestemont, P. Long-term tryptophan supplementation decreased the welfare and innate immune status of pikeperch juveniles. Fish Shellfish Immunol. 2016, 53, 113–114. [Google Scholar] [CrossRef]
- Xiong, Y.; Li, Q.; Ding, Z.; Zheng, J.; Zhou, D.; Wei, S.; Han, X.; Cheng, X.; Li, X.; Xue, Y. Dietary α-lipoic acid requirement and its effects on antioxidant status, carbohydrate metabolism, and intestinal microflora in oriental river prawn Macrobrachium nipponense (De Haan). Aquaculture 2022, 547, 737531. [Google Scholar] [CrossRef]
- Gao, J.; Xu, K.; Liu, H.N.; Liu, G.; Bai, M.M.; Peng, C.; Li, T.J.; Yin, Y.L. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.W.; Liang, X.X.; Li, D.H.; Gao, H.; Wang, Y.D.; Li, W.T.; Xu, K.; Hu, F.Z. Effect of Dietary Tryptophan on Growth, Intestinal Microbiota, and Intestinal Gene Expression in an Improved Triploid Crucian Carp. Front. Nutr. 2021, 8, 322. [Google Scholar] [CrossRef] [PubMed]
- Laranja, J.L.Q.; Quinitio, E.T.; Catacutan, M.R.; Coloso, R.M. Effects of dietary l-tryptophan on the agonistic behavior, growth and survival of juvenile mud crab Scylla serrata. Aquaculture 2010, 310, 84–90. [Google Scholar] [CrossRef]
- Harlioglu, M.M.; Harlioglu, A.G.; Yonar, S.M.; Duran, T.C. Effects of dietary l-tryptophan on the agonistic behavior, growth, and survival of freshwater crayfish Astacus leptodactylus Eschscholtz. Aquacult. Int. 2014, 22, 733–748. [Google Scholar] [CrossRef]
- Qi, T.; Liu, J.; Zhao, P.; Ge, B.; Liu, Q.; Jiang, S.; Wang, Z.; Zhang, H.; Tang, B.; Ding, G.; et al. A novel modulation of physiological regulation in cultured Chinese mitten crab (Eriocheir japonica sinensis) in response to consistent salinity changes. Gene 2020, 756, 144914. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Jin, M.; Ding, L.; Lu, Y.; Ma, H.; Yuan, Y.; Zhou, Q. Dietary lipid levels could improve growth and intestinal microbiota of juvenile swimming crab, Portunus trituberculatus. Aquaculture 2018, 490, 208–216. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, J.; Li, J.; Zou, J.; Fan, L. The immune defense response of Pacific white shrimp (Litopenaeus vannamei) to temperature fluctuation. Fish Shellfish Immunol. 2020, 103, 103–110. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Song, X.; Pang, Y.; Song, Y.; Cheng, Y.; Yang, X. Dietary L-Tryptophan Modulates the Hematological Immune and Antibacterial Ability of the Chinese Mitten Crab, Eriocheir sinensis, Under Cheliped Autotomy Stress. Front. Immunol. 2018, 9, 2744. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Feng, L.; Jiang, W.; Liu, Y.; Jiang, J.; Li, S.; Tang, L.; Zhang, Y.; Kuang, S.; Zhou, X. Dietary tryptophan modulates intestinal immune response, barrier function, antioxidant status and gene expression of TOR and Nrf2 in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2014, 40, 275–287. [Google Scholar] [CrossRef]
- Terrón, M.P.; Delgado, J.; Paredes, S.D.; Barriga, C.; Reiter, R.J.; Rodríguez, A.B. Effect of melatonin and tryptophan on humoral immunity in young and old ringdoves (Streptopelia risoria). Exp. Gerontol. 2009, 44, 653–658. [Google Scholar] [CrossRef]
- She, Q.; Han, Z.; Liang, S.; Xu, W.; Li, X.; Zhao, Y.; Wei, H.; Dong, J.; Li, Y. Impacts of circadian rhythm and melatonin on the specific activities of immune and antioxidant enzymes of the Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2019, 89, 345–353. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, X.-Z.; Xu, M.-J.; Huang, G.-Y.; Zhang, Q.; Cheng, Y.-X.; He, L.; Ren, H.-Y. Melatonin Promotes Cheliped Regeneration, Digestive Enzyme Function, and Immunity Following Autotomy in the Chinese Mitten Crab, Eriocheir sinensis. Front. Physiol. 2018, 9, 269. [Google Scholar] [CrossRef]
- Yang, X.; Song, X.; Zhang, C.; Pang, Y.; Song, Y.; Cheng, Y.; Nie, L.; Zong, X. Effects of dietary melatonin on hematological immunity, antioxidant defense and antibacterial ability in the Chinese mitten crab, Eriocheir sinensis. Aquaculture 2020, 529, 735578. [Google Scholar] [CrossRef]
- Arrigoni, O.; De Tullio, M.C. Ascorbic acid: Much more than just an antioxidant. BBA-Gen. Subj. 2002, 1569, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Padh, H. Cellular functions of ascorbic acid. Int. J. Biochem. Cell Biol. 1990, 68, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Q.; Kosten, T.R.; Zhang, X.Y. Free radicals, antioxidant defense systems, and schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 46, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-L.; Zuo, D.; Wang, L.-M.; Sun, T.; Wang, Q.; Zhao, Y.-L. Effects of white spot syndrome virus infection on immuno-enzyme activities and ultrastructure in gills of Cherax quadricarinatus. Fish Shellfish Immunol. 2012, 32, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Sharf, Y.; Khan, M.A. Dietary tryptophan requirement of fingerling Channa punctatus (Bloch) based on growth, hematological parameters, intestinal enzymes, non-specific immune response, and antioxidant capacity. Aquaculture 2022, 562, 738745. [Google Scholar] [CrossRef]
- Costas, B.; Aragao, C.; Dias, J.; Afonso, A.; Conceicao, L.E.C. Interactive effects of a high-quality protein diet and high stocking density on the stress response and some innate immune parameters of Senegalese sole Solea senegalensis. Fish Physiol. Biochem. 2013, 39, 1141–1151. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species. J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Regenstein, J.M.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. The oxidative stress and antioxidant responses of Litopenaeus vannamei to low temperature and air exposure. Fish Shellfish Immunol. 2018, 72, 564–571. [Google Scholar] [CrossRef]
- González Esquivel, D.; Ramírez-Ortega, D.; Pineda, B.; Castro, N.; Ríos, C.; Pérez de la Cruz, V. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology 2017, 112, 331–345. [Google Scholar] [CrossRef]
- Réus, G.Z.; Becker, I.R.; Scaini, G.; Petronilho, F.; Oses, J.P.; Kaddurah-Daouk, R.; Ceretta, L.B.; Zugno, A.I.; Dal-Pizzol, F.; Quevedo, J.; et al. The inhibition of the kynurenine pathway prevents behavioral disturbances and oxidative stress in the brain of adult rats subjected to an animal model of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 55–63. [Google Scholar] [CrossRef]
- Requena, J.R.; Fu, M.X.; Ahmed, M.U.; Jenkins, A.J.; Lyons, T.J.; Thorpe, S.R. Lipoxidation products as biomarkers of oxidative damage to proteins during lipid peroxidation reactions. Nephrol. Dial. Transplant. 1996, 11, 48–53. [Google Scholar] [CrossRef]
- Lee, S.-L.; Wang, W.-W.; Fanburg, B.L. Superoxide as an Intermediate Signal for Serotonin-Induced Mitogenesis. (This study was supported by Research Grant HL 32723 from the Heart, Lung, and Blood Institute of the National Institutes of Health.). Free Radicals Biol. Med. 1998, 24, 855–858. [Google Scholar] [CrossRef]
- Mukhin, Y.V.; Garnovskaya, M.N.; Collinsworth, G.; Grewal, J.S.; Pendergrass, D.; Nagai, T.; Pinckney, S.; Greene, E.L.; Raymond, J.R. 5-Hydroxytryptamine1A receptor/Giβγ stimulates mitogen-activated protein kinase via NAD(P)H oxidase and reactive oxygen species upstream of Src in Chinese hamster ovary fibroblasts. Biochem. J. 2000, 347, 61–67. [Google Scholar] [CrossRef]
- Fang, X.-L.; Shu, G.; Yu, J.-J.; Wang, L.-N.; Yang, J.; Zeng, Q.-J.; Cheng, X.; Zhang, Z.-Q.; Wang, S.-B.; Gao, P.; et al. The Anorexigenic Effect of Serotonin Is Mediated by the Generation of NADPH Oxidase-Dependent ROS. PLoS ONE 2013, 8, e0053142. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.-D.; Wen, H.-L.; Liu, Y.; Jiang, J.; Wu, P.; Zhao, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Enhanced muscle nutrient content and flesh quality, resulting from tryptophan, is associated with anti-oxidative damage referred to the Nrf2 and TOR signalling factors in young grass carp (Ctenopharyngodon idella): Avoid tryptophan deficiency or excess. Food Chem. 2016, 199, 210–219. [Google Scholar] [CrossRef]
- Yao, K.; Fang, J.; Yin, Y.-L.; Feng, Z.-M.; Tang, Z.-R.; Wu, G. Tryptophan metabolism in animals: Important roles in nutrition and health. Front. Biosci. 2011, 3, 286–297. [Google Scholar]
- Sacristán, H.J.; Mufari, J.R.; Lorenzo, R.A.; Boy, C.C.; Lovrich, G.A. Ontogenetic changes in energetic reserves, digestive enzymes, amino acid and energy content of Lithodes santolla (Anomura: Lithodidae): Baseline for culture. PLoS ONE 2020, 15, e0232880. [Google Scholar] [CrossRef]
- Shan, H.; Wang, T.; Dong, Y.; Ma, S. Effects of dietary Ampithoe sp. supplementation on the growth, energy status, antioxidant capacity, and ammonia-N tolerance of the shrimp Litopenaeus vannamei: Continuous versus interval feeding. Aquaculture 2019, 509, 32–39. [Google Scholar] [CrossRef]
- Sun, F.; Xu, Z. Significant Differences in Intestinal Microbial Communities in Aquatic Animals from an Aquaculture Area. J. Mar. Sci. Eng. 2021, 9, 104. [Google Scholar] [CrossRef]
- Wu, J.; Tian, S.; Luo, K.; Zhang, Y.; Pan, H.; Zhang, W.; Mai, K. Dietary recombinant human lysozyme improves the growth, intestinal health, immunity and disease resistance of Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2022, 121, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Fiore, C.L.; Longnecker, K.; Kido Soule, M.C.; Kujawinski, E.B. Release of ecologically relevant metabolites by the cyanobacterium Synechococcus elongatus CCMP 1631. Environ. Microbiol. 2015, 17, 3949–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.; Huang, Z.; Liang, L.; Li, N.; Ali, Z.; Ding, L.; Hong, M.; Shi, H. Ammonia stress influences intestinal histomorphology, immune status and microbiota of Chinese striped-neck turtle (Mauremys sinensis). Ecotoxicol. Environ. Saf. 2021, 222, 112471. [Google Scholar] [CrossRef]
- Xu, C.; Zhong, X.-Q.; Li, X.-F.; Shi, H.-J.; Liu, W.-B. Regulation of growth, intestinal microflora composition and expression of immune-related genes by dietary supplementation of Streptococcus faecalis in blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2020, 105, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Gatesoupe, F.-J.; Li, T.; Wang, X.; Zhang, Q.; Feng, D.; Feng, Y.; Chen, H.; Li, A. Significant improvement of intestinal microbiota of gibel carp (Carassius auratus gibelio) after traditional Chinese medicine feeding. J. Appl. Microbiol. 2018, 124, 829–841. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, S.; Dong, X.; Chi, S.; Yang, Q.; Liu, H.; Tan, B.; Xie, S. Effects of fishmeal replacement by black soldier fly on growth performance, digestive enzyme activity, intestine morphology, intestinal flora and immune response of pearl gentian grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Fish Shellfish Immunol. 2022, 120, 497–506. [Google Scholar] [CrossRef]
- Nakazawa, H.; Enei, H.; Okumura, S.; Yamada, H. Synthesis of l-Tryptophan from Pyruvate, Ammonia and Indole. Agric. Biol. Chem. 1972, 36, 2523–2528. [Google Scholar] [CrossRef]
- Liang, H.; Dai, Z.; Liu, N.; Ji, Y.; Chen, J.; Zhang, Y.; Yang, Y.; Li, J.; Wu, Z.; Wu, G. Dietary L-Tryptophan Modulates the Structural and Functional Composition of the Intestinal Microbiome in Weaned Piglets. Front Microbiol. 2018, 9, 1736. [Google Scholar] [CrossRef]
- Qi, Q.; Li, J.; Yu, B.; Moon, J.-Y.; Chai, J.C.; Merino, J.; Hu, J.; Ruiz-Canela, M.; Rebholz, C.; Wang, Z.; et al. Host and gut microbial tryptophan metabolism and type 2 diabetes: An integrative analysis of host genetics, diet, gut microbiome and circulating metabolites in cohort studies. Gut 2022, 71, 1095–1105. [Google Scholar] [CrossRef]
- Tan, P.; Wu, X.; Zhu, W.L.; Lou, B.; Chen, R.Y.; Wang, L.G. Effect of tributyrin supplementation in high-soya bean meal diet on growth performance, body composition, intestine morphology and microbiota of juvenile yellow drum (Nibea albiflora). Aquacult. Res. 2020, 51, 2004–2019. [Google Scholar] [CrossRef]
- Oren, A. The Family Rhodocyclaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 975–998. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, M.; Pang, Y.; Niu, C.; Zhang, D.; Zhang, Y.; Liu, Z.; Song, Y.; Shi, A.; Chen, Q.; Zhang, J.; et al. Effects of Dietary L-TRP on Immunity, Antioxidant Capacity and Intestinal Microbiota of the Chinese Mitten Crab (Eriocheir Sinensis) in Pond Culture. Metabolites 2023, 13, 1. https://doi.org/10.3390/metabo13010001
Hou M, Pang Y, Niu C, Zhang D, Zhang Y, Liu Z, Song Y, Shi A, Chen Q, Zhang J, et al. Effects of Dietary L-TRP on Immunity, Antioxidant Capacity and Intestinal Microbiota of the Chinese Mitten Crab (Eriocheir Sinensis) in Pond Culture. Metabolites. 2023; 13(1):1. https://doi.org/10.3390/metabo13010001
Chicago/Turabian StyleHou, Mengna, Yangyang Pang, Chao Niu, Dongxin Zhang, Ying Zhang, Zhiqiang Liu, Yameng Song, Aoya Shi, Qing Chen, Junyan Zhang, and et al. 2023. "Effects of Dietary L-TRP on Immunity, Antioxidant Capacity and Intestinal Microbiota of the Chinese Mitten Crab (Eriocheir Sinensis) in Pond Culture" Metabolites 13, no. 1: 1. https://doi.org/10.3390/metabo13010001
APA StyleHou, M., Pang, Y., Niu, C., Zhang, D., Zhang, Y., Liu, Z., Song, Y., Shi, A., Chen, Q., Zhang, J., Cheng, Y., & Yang, X. (2023). Effects of Dietary L-TRP on Immunity, Antioxidant Capacity and Intestinal Microbiota of the Chinese Mitten Crab (Eriocheir Sinensis) in Pond Culture. Metabolites, 13(1), 1. https://doi.org/10.3390/metabo13010001