

Quercetin, a Plant Flavonol Attenuates Diabetic Complications, Renal Tissue Damage, Renal Oxidative Stress and Inflammation in Streptozotocin-Induced Diabetic Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal Ethics
2.3. Animals and Treatment
2.4. Animal Groups and Treatment Plan
| Group Name | Short Name | Treatment Plan |
| Normal control | C | Rats with free access to rat pellets and orally given saline as a placebo |
| Negative control | NC | STZ-induced diabetic rats at 55 mg/kg b.w. [23] and orally given saline. |
| Positive control | PC | STZ-induced diabetic rats and oral gavage treatment with glibenclamide (5 mg/kg b.w.) [25] as a standard drug. |
| Quercetin Treatment | QT | STZ-induced diabetic rats and oral gavage treatment with quercetin (50 mg/kg b.w.) |
2.5. Measurement of Body Weight
2.6. Fasting Blood Glucose and Insulin Level Measurement
2.7. Measurement of Lipid Profile
2.8. Oral Glucose Tolerance Test (OGTT)
2.9. Measurement of Kidney Function Parameters
2.10. Measurement of TNF-α, IL-6 and IL-1β Pro-Inflammatory Parameters
2.11. Measurement of Lipid Peroxidation
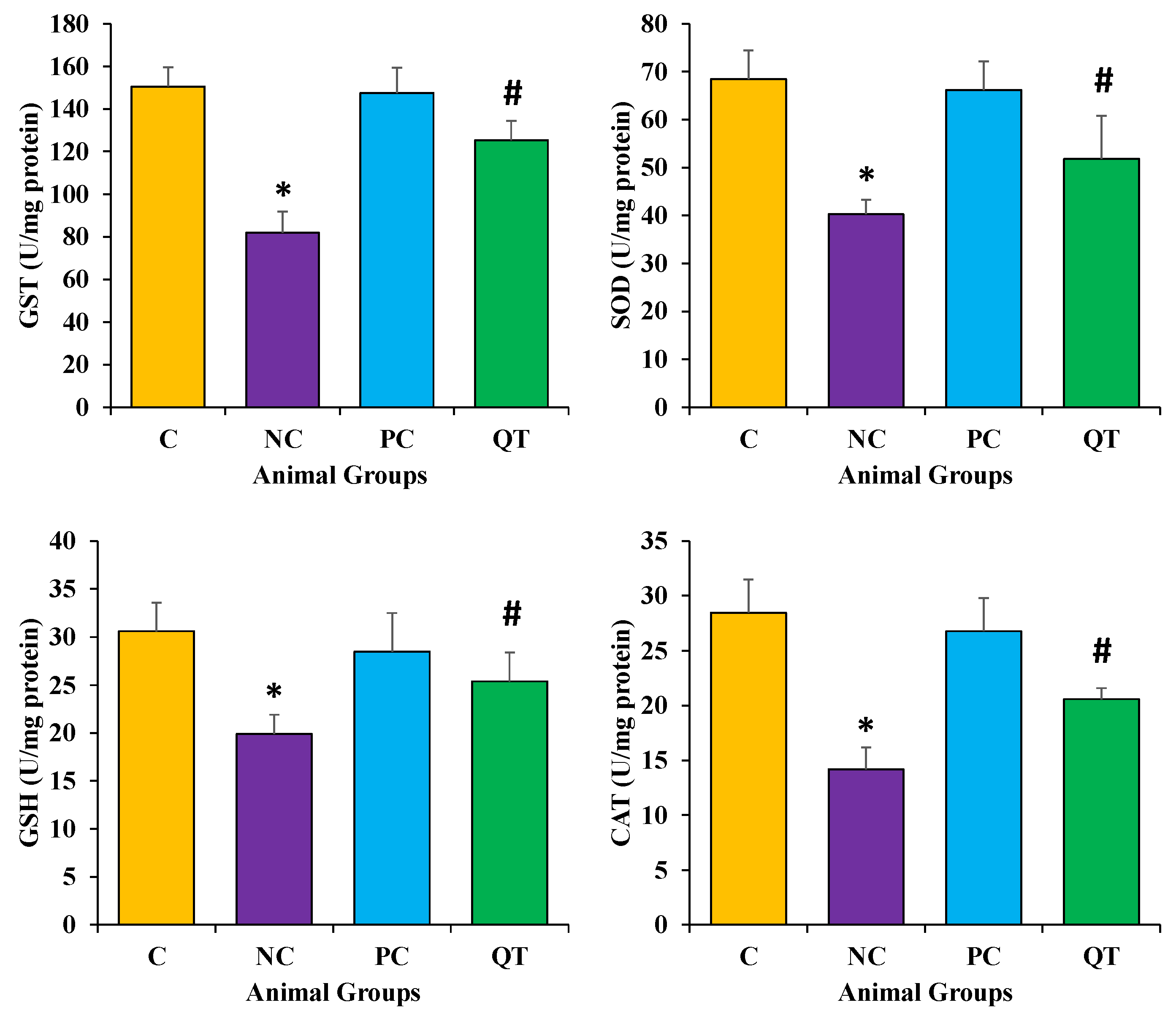
2.12. Determination of SOD, GST, CAT and GSH Levels
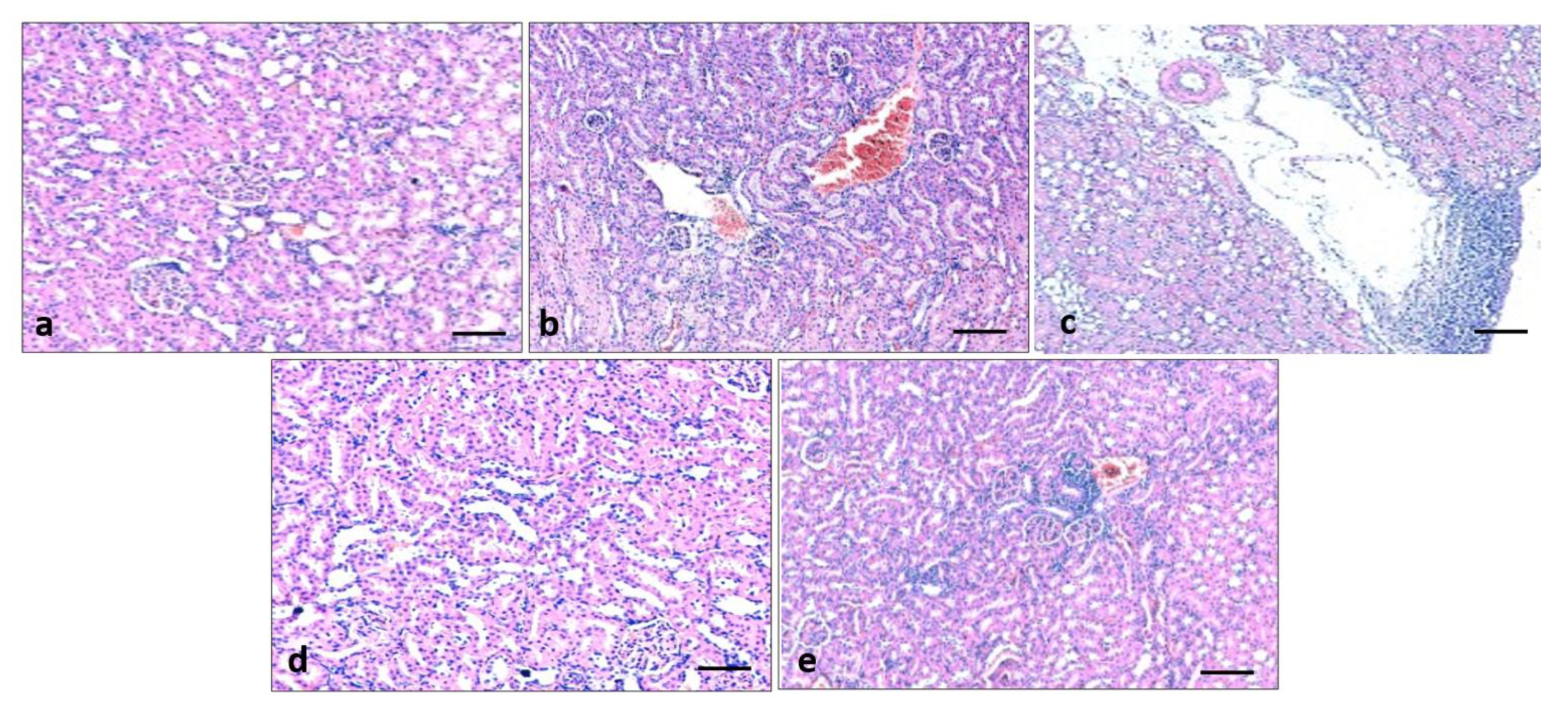
2.13. Histopathological Examination
2.14. Masson’s Trichrome Staining
2.15. Picro Sirius Red Staining
2.16. Expressional Evaluation of COX-2 Protein through Immunohistochemical Staining
2.17. Statistical Analysis
3. Results
3.1. Quercetin Effects on Body Weight
3.2. Effect of Quercetin on Glucose and Insulin Levels
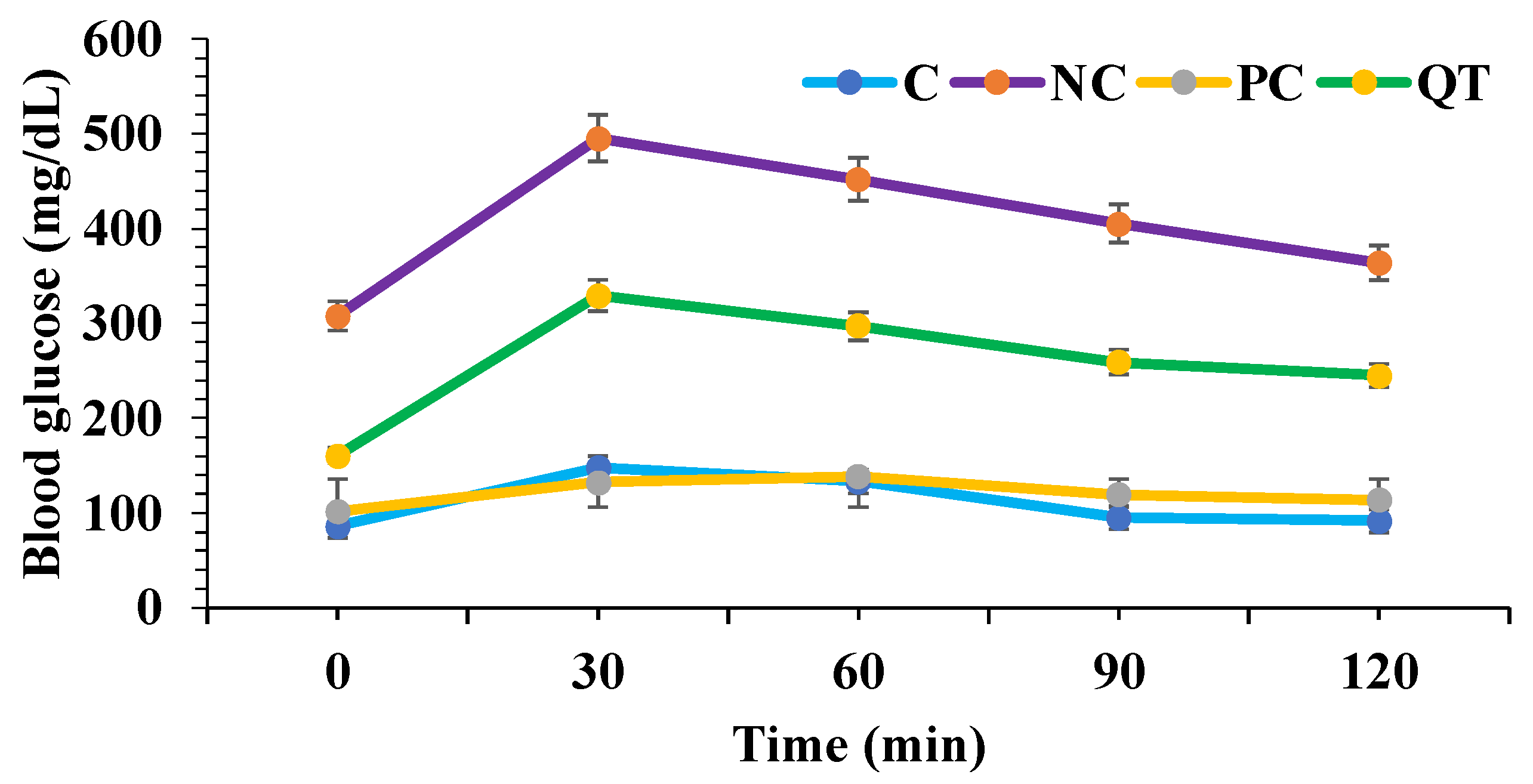
3.3. Effect of Quercetin on Oral Glucose Tolerance Tests
3.4. Effect of Quercetin on Lipid Profile
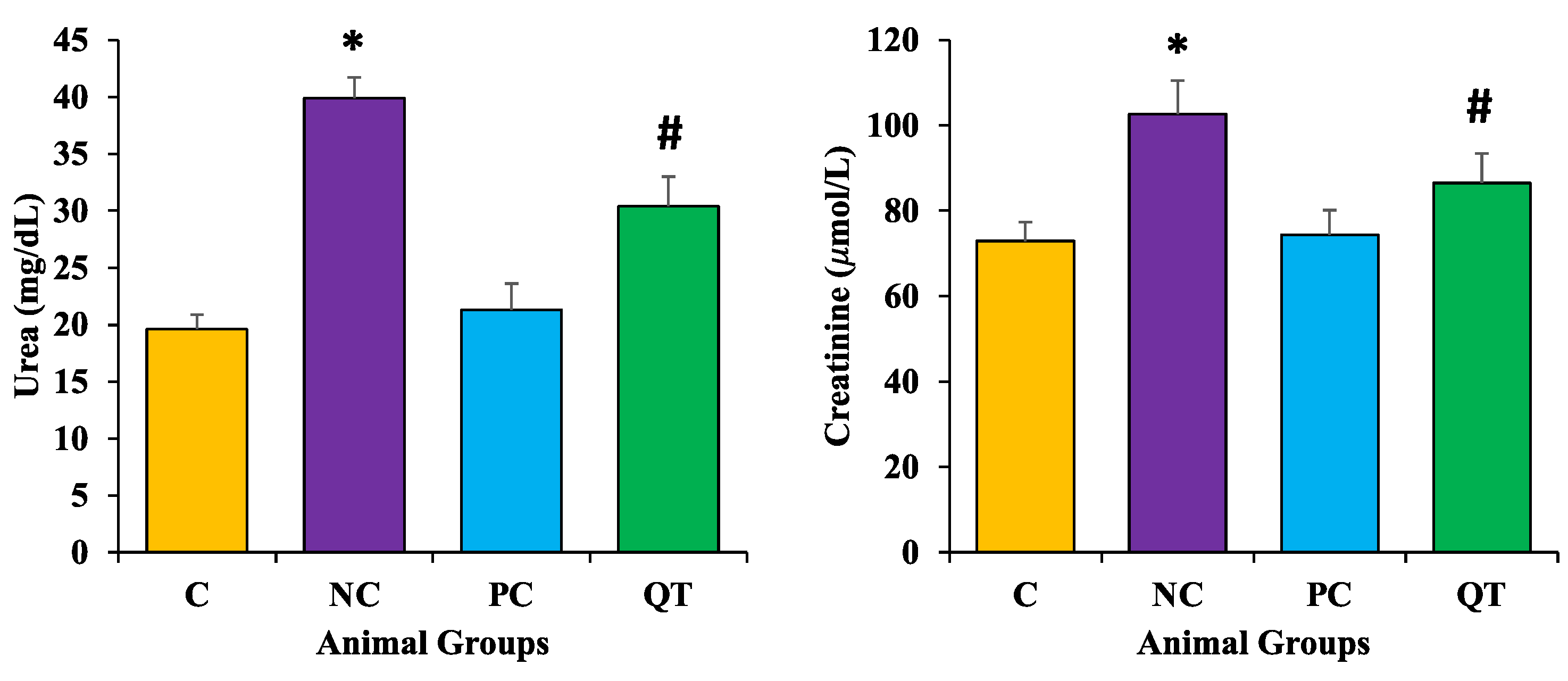
3.5. Effect of Quercetin on Kidney Function Profile
3.6. Effect of Quercetin on Oxidative Stress
3.7. Effect of Quercetin Extract on Lipid Peroxidation
3.8. Effect of Quercetin on Inflammatory Markers Level
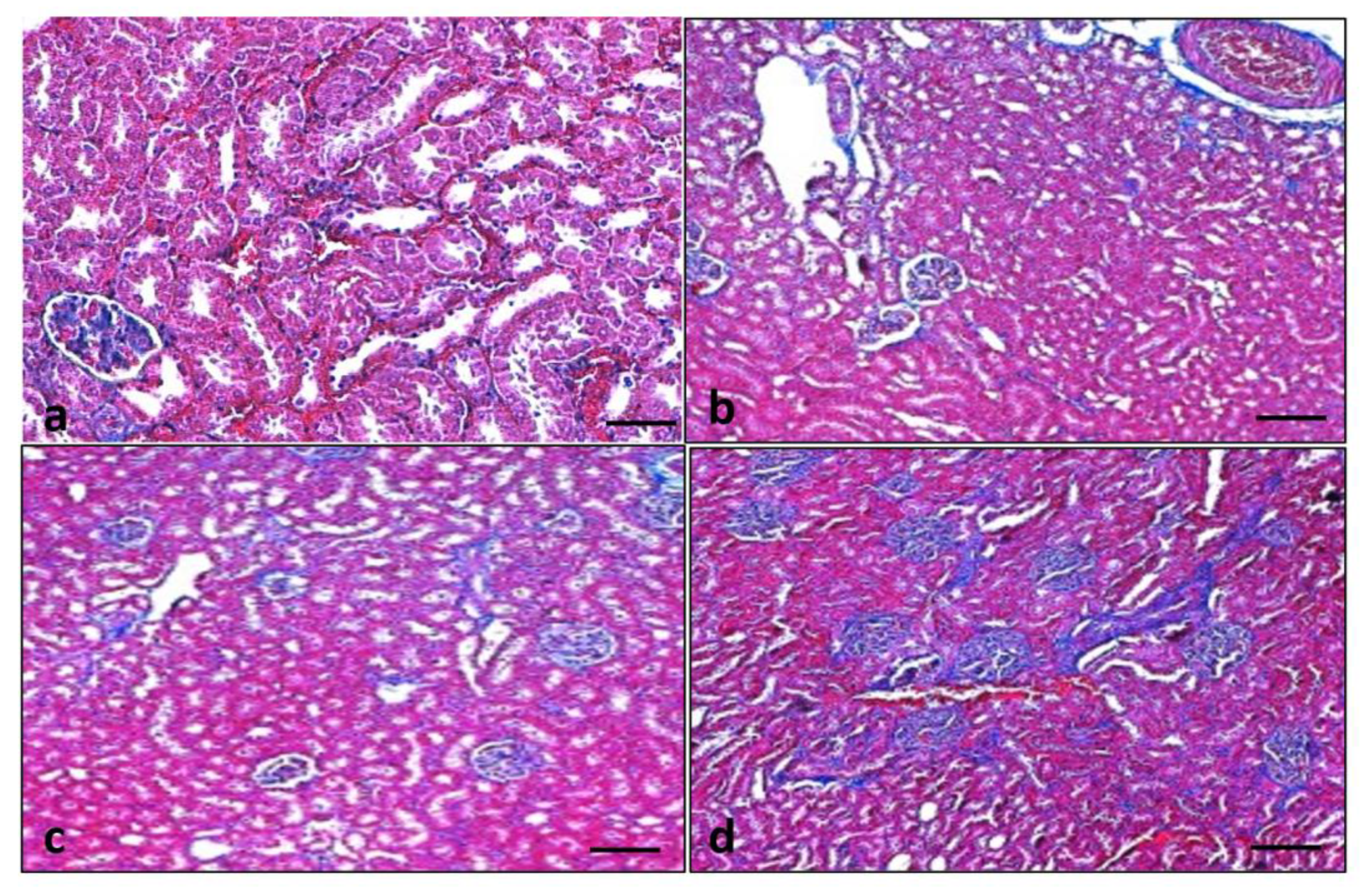
3.9. Effect of Quercetin on Kidney Histology
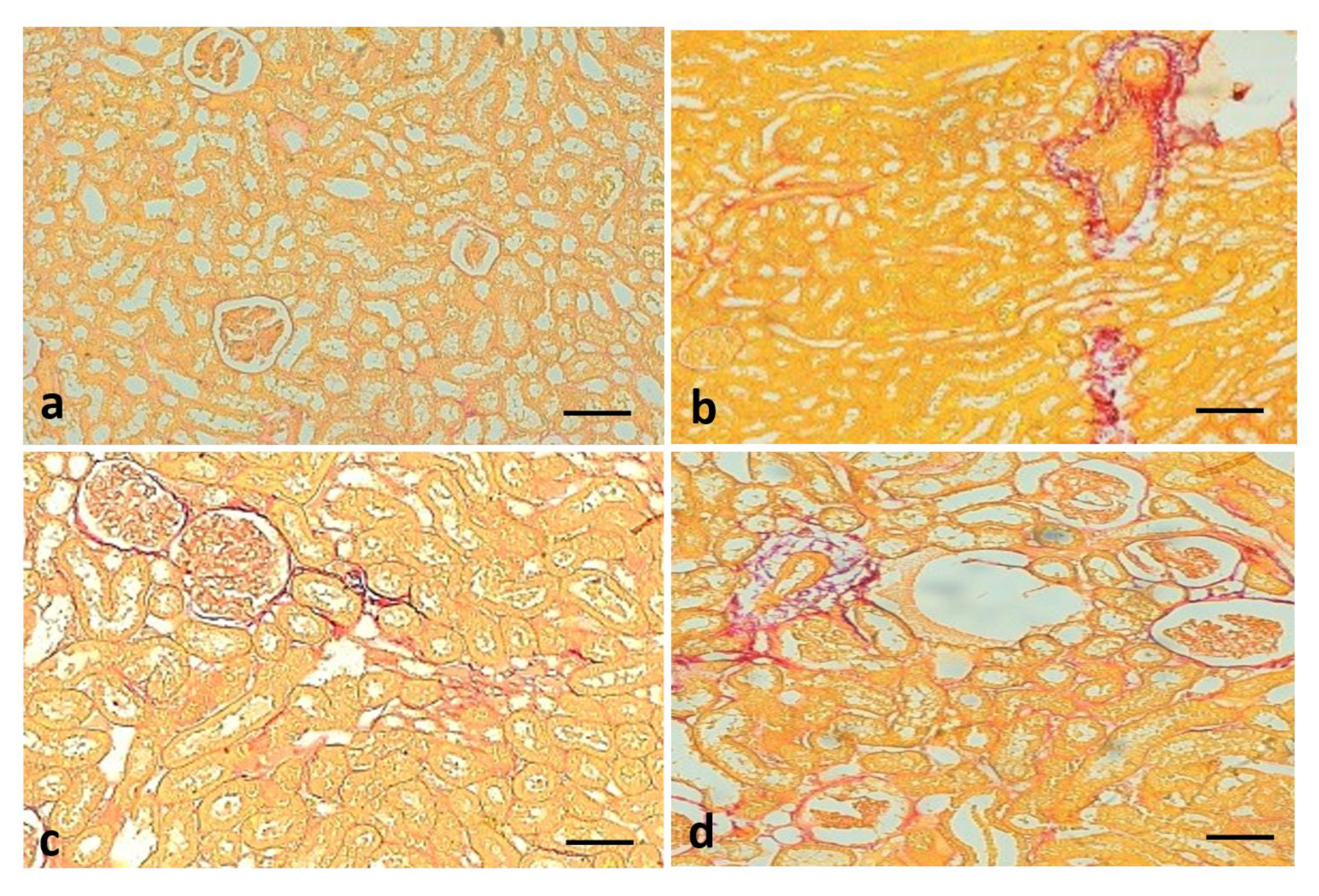
3.10. Effect of Quercetin on Renal Fibrosis
3.11. Effect of Quercetin on Renal Fibrosis of STZ-Induced Diabetic Rats
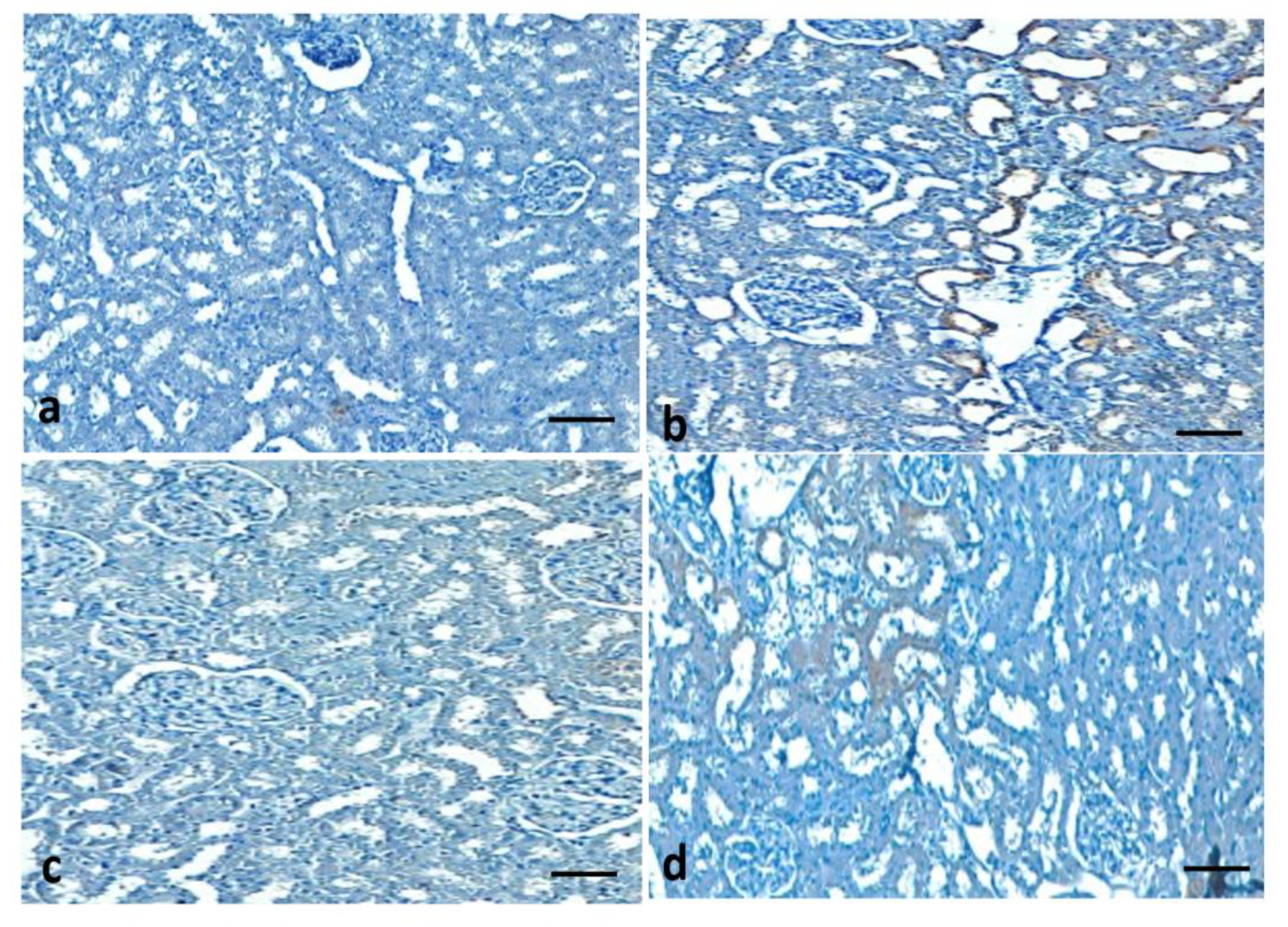
3.12. Effect of Quercetin on COX-2 Protein Expression of STZ-Induced Diabetic Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017. [Google Scholar]
- Nowotny, K.; Jung, T.; Hohn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [Green Version]
- Abdali, D.; Samson, S.E.; Grover, A.K. How effective are antioxidant supplements in obesity and diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Gurzov, E.N.; Tran, M.; Fernandez-Rojo, M.A.; Merry, T.L.; Zhang, X.; Xu, Y.; Fukushima, A.; Waters, M.J.; Watt, M.J.; Andrikopoulos, S.; et al. Hepatic oxidative stress promotes insulin-STAT-5 signaling and obesity by inactivating protein tyrosine phosphatase N2. Cell Metab. 2014, 20, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, W.; Chen, H.; Liao, N.; Wang, Z.; Zhang, X.; Hai, C. High selenium impairs hepatic insulin sensitivity through opposite regulation of ROS. Toxicol. Lett. 2014, 224, 16–23. [Google Scholar] [CrossRef]
- Taskinen, M.R. Diabetic dyslipidemia. Atheroscler. Suppl. 2002, 3, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, F.; Gharib, H.; Golbert, A.; Griz, L.; Faria, M. An overview on management of diabetic dyslipidemia. J. Diabetes Endocrinol. 2014, 4, 27–36. [Google Scholar]
- Wu, L.; Parhofer, K.G. Diabetic dyslipidemia. Metab. Clin. Exp. 2014, 63, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Krentz, A.J. Comparative safety of newer oral antidiabetic drugs. Expert Opin. Drug Saf. 2006, 5, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Rafaeloff, R.; Pittenger, G.L.; Barlow, S.W.; Qin, X.F.; Yan, B.; Rosenberg, L.; Duguid, W.P.; Vinik, A.I. Cloning and sequencing of the pancreatic islet neogenesis associated protein (INGAP) gene and its expression in islet neogenesis in hamsters. J. Clin. Investig. 1997, 99, 2100–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Zhang, H.; Ye, J. Traditional Chinese medicine in treatment of metabolic syndrome. Endocr. Metab. Immune Disord. Drug Targets 2008, 8, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Dhasarathan, P.; Theriappan, P. Evaluation of anti-diabetic activity of Strychonous potatorum in alloxan induced diabetic rats. J. Med. Sci. 2011, 2, 670–674. [Google Scholar]
- Ramesh, B.; Pugalendi, K.V. Anti-hyperglycemic effect of Umbelliferone in Streptozotocin diabetic rats. J. Med. Plants. 2006, 9, 562–566. [Google Scholar]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [PubMed]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.M.; Murphy, A.; Carmichael, M.D.; Davis, B. Quercetin increases brain and muscle mitochondrial biogenesis and exercise tolerance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1071–R1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi-Boroujeni, A.; Mahmoudian-Sani, M.-R. Anti-inflammatory potential of Quercetin in COVID-19 treatment. J. Inflamm. 2021, 18, 3. [Google Scholar]
- Alzohairy, M.A.; Khan, A.A.; Ansari, M.A.; Babiker, A.Y.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effect of Quercetin, a Flavonol against Benzo(a)pyrene-Induced Lung Injury via Inflammation, Oxidative Stress, Angiogenesis and Cyclooxygenase-2 Signalling Molecule. Appl. Sci. 2021, 11, 8675. [Google Scholar] [CrossRef]
- Chen, B.H.; Park, J.H.; Ahn, J.H.; Cho, J.H.; Kim, I.H.; Lee, J.C.; Won, M.H.; Lee, C.H.; Hwang, I.K.; Kim, J.D.; et al. Pretreated quercetin protects gerbil hippocampal CA1 pyramidal neurons from transient cerebral ischemic injury by increasing the expression of antioxidant enzymes. Neural Regen Res. 2017, 12, 220–227. [Google Scholar]
- Braun, K.F.; Ehnert, S.; Freude, T.; Egaña, J.T.; Schenck, T.L.; Buchholz, A.; Schmitt, A.; Siebenlist, S.; Schyschka, L.; Neumaier, M.; et al. Quercetin protects primary human osteoblasts exposed to cigarette smoke through activation of the antioxidative enzymes HO-1 and SOD-1. Sci. World J. 2011, 11, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Izak-Shirian, F.; Najafi-Asl, M.; Azami, B.; Heidarian, E.; Najafi, M.; Khaledi, M.; Nouri, A. Quercetin exerts an ameliorative effect in the rat model of diclofenac-induced renal injury through mitigation of inflammatory response and modulation of oxidative stress. Eur. J. Inflamm. 2022, 20, 1721727X221086530. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential therapeutic targets of quercetin, a plant flavonol, and its role in the therapy of various types of cancer through the modulation of various cell signaling pathways. Molecules 2021, 26, 1315. [Google Scholar]
- Strugała, P.; Dzydzan, O.; Brodyak, I.; Kucharska, A.Z.; Kuropka, P.; Liuta, M.; Kaleta-Kuratewicz, K.; Przewodowska, A.; Michałowska, D.; Gabrielska, J.; et al. Antidiabetic and Antioxidative Potential of the Blue Congo Variety of Purple Potato Extract in Streptozotocin-Induced Diabetic Rats. Molecules 2019, 24, 3126. [Google Scholar] [CrossRef] [Green Version]
- Ola, M.S.; Ahmed, M.M.; Shams, S.; Al-Rejaie, S.S. Neuroprotective effects of quercetin in diabetic rat retina. Saudi J. Biol. Sci. 2017, 24, 1186–1194. [Google Scholar] [CrossRef]
- Nazir, N.; Zahoor, M.; Nisar, M.; Khan, I.; Ullah, R.; Alotaibi, A. Antioxidants Isolated from Elaeagnus umbellata (Thunb.) Protect against Bacterial Infections and Diabetes in Streptozotocin-Induced Diabetic Rat Model. Molecules 2021, 26, 4464. [Google Scholar] [CrossRef] [PubMed]
- Husain, N.E.; Babiker, A.Y.; Albutti, A.S.; Alsahli, M.A.; Aly, S.M.; Rahmani, A.H. Clinicopathological Significance of Vimentin and Cytokeratin Protein in the Genesis of Squamous Cell Carcinoma of Cervix. Obstet. Gynecol. Int. 2016, 2016, 8790120. [Google Scholar]
- Babiker, A.Y.; Almatroudi, A.; Allemailem, K.S.; Husain, N.E.O.; Alsammani, M.A.; Alsahli, M.A.; Rahmani, A.H. Clinicopathologic Aspects of Squamous Cell Carcinoma of the Uterine Cervix: Role of PTEN, BCL2 and P53. Applied Sciences 2018, 8, 2124. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Sultana, S. Abrogation of potassium bromate-induced renal oxidative stress and subsequent cell proliferation response by soy isoflavones in Wistar rats. Toxicology 2004, 201, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Bigagli, E.; Lodovici, M. Circulating oxidative stress biomarkers in clinical studies on type 2 diabetes and its complications. Oxid. Med. Cell. Longev. 2019, 2019, 5953685. [Google Scholar] [PubMed] [Green Version]
- Rahmani, A.H.; Aldebasi, Y.H.; Srikar, S.; Khan, A.A.; Aly, S.M. Aloe vera: Potential candidate in health management via modulation of biological activities. Pharmacogn. Rev. 2015, 9, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmani, A.H. Cassia fistula Linn: Potential candidate in the health management. Pharmacogn. Res. 2015, 7, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, A.H.; Aly, S.M.; Ali, H.; Babiker, A.Y.; Srikar, S.; Khan, A.A. Therapeutic effects of date fruits (Phoenix dactylifera) in the prevention of diseases via modulation of anti-inflammatory, anti-oxidant and anti-tumour activity. Int. J. Clin. Exp. Med. 2014, 7, 483–491. [Google Scholar]
- Almatroodi, S.A.; Almatroudi, A.; Alsahli, M.A.; Rahmani, A.H. Grapes and their Bioactive Compounds: Role in Health Management Through Modulating Various Biological Phcogj.com Activities. Pharmacogn. J. 2020, 12, 1455–1462. [Google Scholar] [CrossRef]
- Priscilla, D.H.; Jayakumar, M.; Thirumurgan, K. Flavanone naringenin: An effective antihyperglycemic and antihyperlipidemic nutraceutical agent on high fat diet fed streptozotocin induced type 2 diabetic rats. J. Func. Foods 2015, 14, 363–373. [Google Scholar]
- Bhutada, P.; Mundhada, Y.; Bansod, K.; Bhutada, C.; Tawari, S.; Dixit, P.; Mundhada, D. Ameliorative effect of quercetin on memory dysfunction in streptozotocin-induced diabetic rats. Neurobiol. Learn Mem. 2010, 94, 293–302. [Google Scholar] [CrossRef]
- Gomes, I.B.S.; Porto, M.L.; Santos, M.C.L.F.S.; Campagnaro, B.P.; Gava, A.L.; Meyrelles, S.S.; Pereira, T.M.C.; Vasquez, E.C. The protective effects of oral low-dose quercetin on diabetic nephropathy in hypercholesterolemic mice. Front. Physiol. 2015, 6, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, I.; Porto, M.L.; Santos, M.C.L.; Campagnaro, B.P.; Pereira, T.; Meyrelles, S.S.; Vasquez, E.C. Renoprotective, anti-oxidative and anti-apoptotic effects of oral low-dose quercetin in the C57BL/6J model of diabetic nephropathy. Lipids Health Dis. 2014, 13, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozcelik, D.; Tuncdemir, M.; Ozturk, M.; Uzun, H. Evaluation of trace elements and oxidative stress levels in the liver and kidney of streptozotocin-induced experimental diabetic rat model. Gen. Physiol. Biophys. 2011, 30, 356–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.K.; Kang, H.S. Anti-Diabetic Effect of Cotreatment with Quercetin and Resveratrol in Streptozotocin-Induced Diabetic Rats. Biomol. Ther. 2018, 26, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvind, K.; Pradeep, R.; Deepa, R.; Mohan, V. Diabetes and coronary artery diseases. Indian J. Med. Res. 2002, 116, 163–176. [Google Scholar]
- Palumbo, P.J. Metformin: Effect on cardiovascular risk factor in patients with non-insulin dependent diabetes mellitus. J. Diabetes Its Complicat. 1998, 12, 110–119. [Google Scholar] [CrossRef]
- Goldberg, R.B. Lipid disorders in diabetes. Diabetes Care 1981, 4, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, P.; Vijayakumar, S.; Kothandaraman, S.; Palani, M. Anti-diabetic activity of quercetin extracted from Phyllanthus emblica L. fruit: In silico and in vivo approaches. J. Pharm. Anal. 2018, 8, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Guo, Y.; Sun, L.; Zhuang, Y. Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice. Nutrients 2017, 9, 801. [Google Scholar] [CrossRef] [Green Version]
- Brown, W.V. Microvascular complications of diabetes mellitus: Renal protection accompanies cardiovascular protection. Am. J. Cardiol. 2008, 102, 10L–13L. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Gajski, G.; Garaj-Vrhovac, V.; Dikić, D.; Prskalo, Z.Š.; Sirovina, D. DNA-protective effects of quercetin or naringenin in alloxan-induced diabetic mice. Eur. J. Pharmacol. 2011, 656, 110–118. [Google Scholar] [CrossRef]
- Wang, C.; Pan, Y.; Zhang, Q.Y.; Wang, F.M.; Kong, L.D. Quercetin and allopurinol ameliorate kidney injury in STZ-treated rats with regulation of renal NLRP3 inflammasome activation and lipid accumulation. PLoS ONE 2012, 7, e38285. [Google Scholar] [CrossRef] [Green Version]
- Elbe, H.; Vardi, N.; Esrefoglu, M.; Ates, B.; Yologlu, S.; Taskapan, C. Amelioration of streptozotocin-induced diabetic nephropathy by melatonin, quercetin, and resveratrol in rats. Hum. Exp. Toxicol. 2014; epub ahead of priint. [Google Scholar]
- Chen, P.; Chen, J.; Zheng, Q.; Chen, W.; Wang, Y.; Xu, X. Pioglitazone, extract of compound Danshen dripping pill, and quercetin ameliorate diabetic nephropathy in diabetic rats. J. Endocrinol. Invest. 2013, 36, 422–427. [Google Scholar]
- Lai, P.B.; Zhang, L.; Yang, L.Y. Quercetin ameliorates diabetic nephropathy by reducing the expressions of transforming growth factor-β1 and connective tissue growth factor in streptozotocin-induced diabetic rats. Ren. Fail. 2012, 34, 83–87. [Google Scholar] [CrossRef]
- Oh, P.S.; Lee, S.J.; Lim, K.T. Hypolipidemic and antioxydative effects of the plant glycoprotein (36 kDa) from Rhus verniciflua stokes fruit in triton Wr-1339 induced hyperlipidemic mice. Biosci. Biotechnol. Biochem. 2006, 70, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Anjaneyulu, M.; Chopra, K. Quercetin, an anti-oxidant bioflavonoid, attenuates diabetic nephropathy in rats. Clin. Exp. Pharmacol. Physiol. 2004, 31, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.L.; Lu, H.Q.; Li, W.N.; Huang, H.P.; Zhou, H.Y.; Leng, E.N.; Zhang, Y.Y. Protective effects of quercetin and crocin in the kidneys and liver of obese Sprague-Dawley rats with Type 2 diabetes: Effects of quercetin and crocin on T2DM rats. Hum. Exp. Toxicol. 2021, 40, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.D.; Wu, C.C.; Hung, C.N.; Wang, C.J.; Huang, H.P. Myrciaria cauliflora extract improves diabetic nephropathy via suppression of oxidative stress and inflammation in streptozotocin-nicotinamide mice. J. Food Drug Anal. 2016, 24, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roslan, J.; Giribabu, N.; Karim, K.; Salleh, N. Quercetin ameliorates oxidative stress, inflammation and apoptosis in the heart of streptozotocin-nicotinamide-induced adult male diabetic rats. Biomed. Pharmacother. 2017, 86, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.K.; Tesch, G.H. Inflammation in diabetic nephropathy. Mediat. Inflamm. 2012, 2012, 146154. [Google Scholar] [CrossRef]
- Wu, C.C.; Hung, C.N.; Shin, Y.C.; Wang, C.J.; Huang, H.P. Myrciaria cauliflora extracts attenuate diabetic nephropathy involving the Ras signaling pathway in streptozotocin/nicotinamide mice on a high fat diet. J. Food Drug analysis 2016, 24, 136–146. [Google Scholar] [CrossRef]












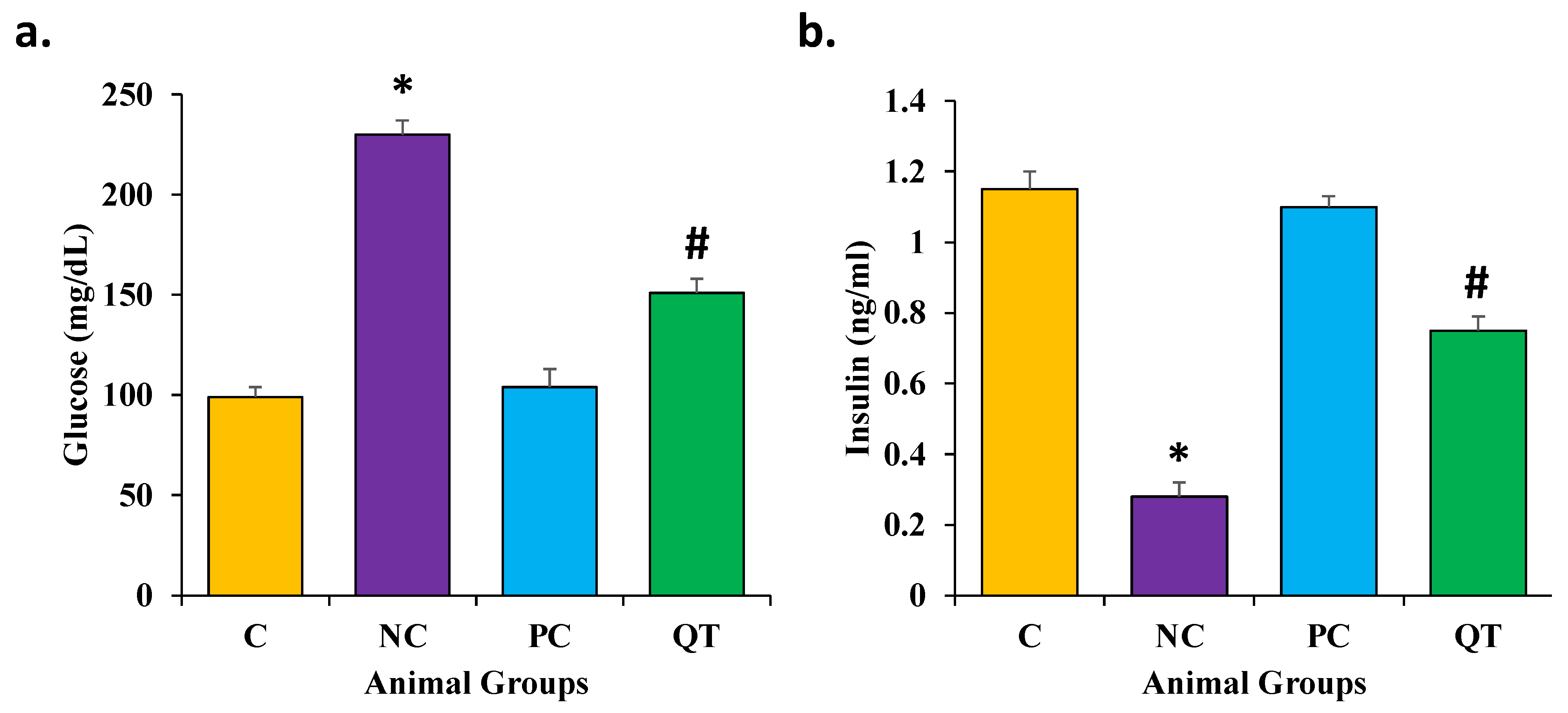{kind=link}
{kind=link}
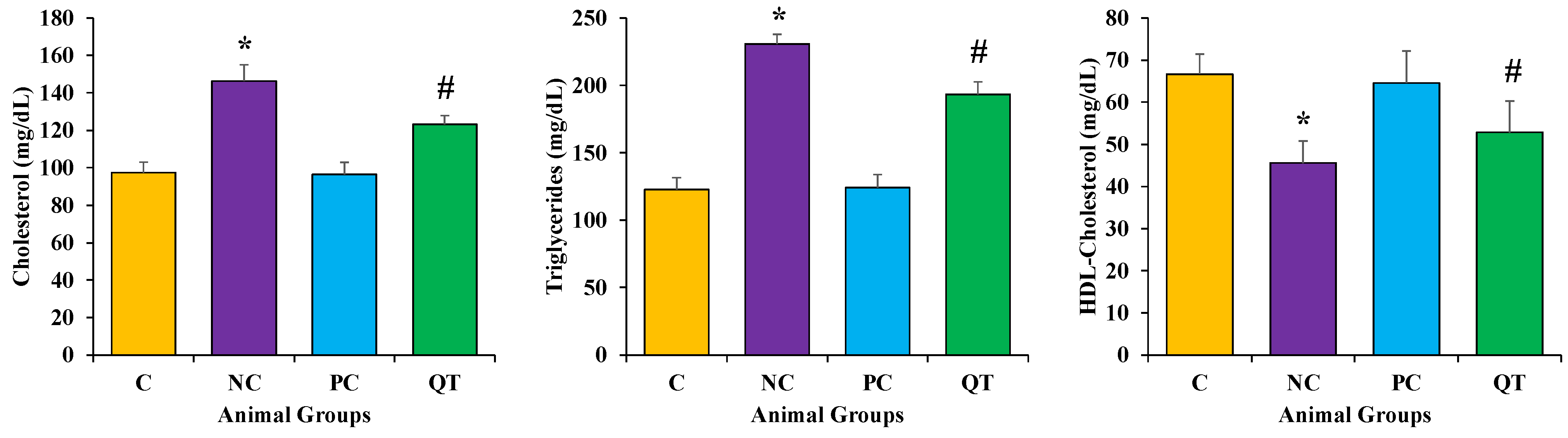{kind=link}
{kind=link}
{kind=link}
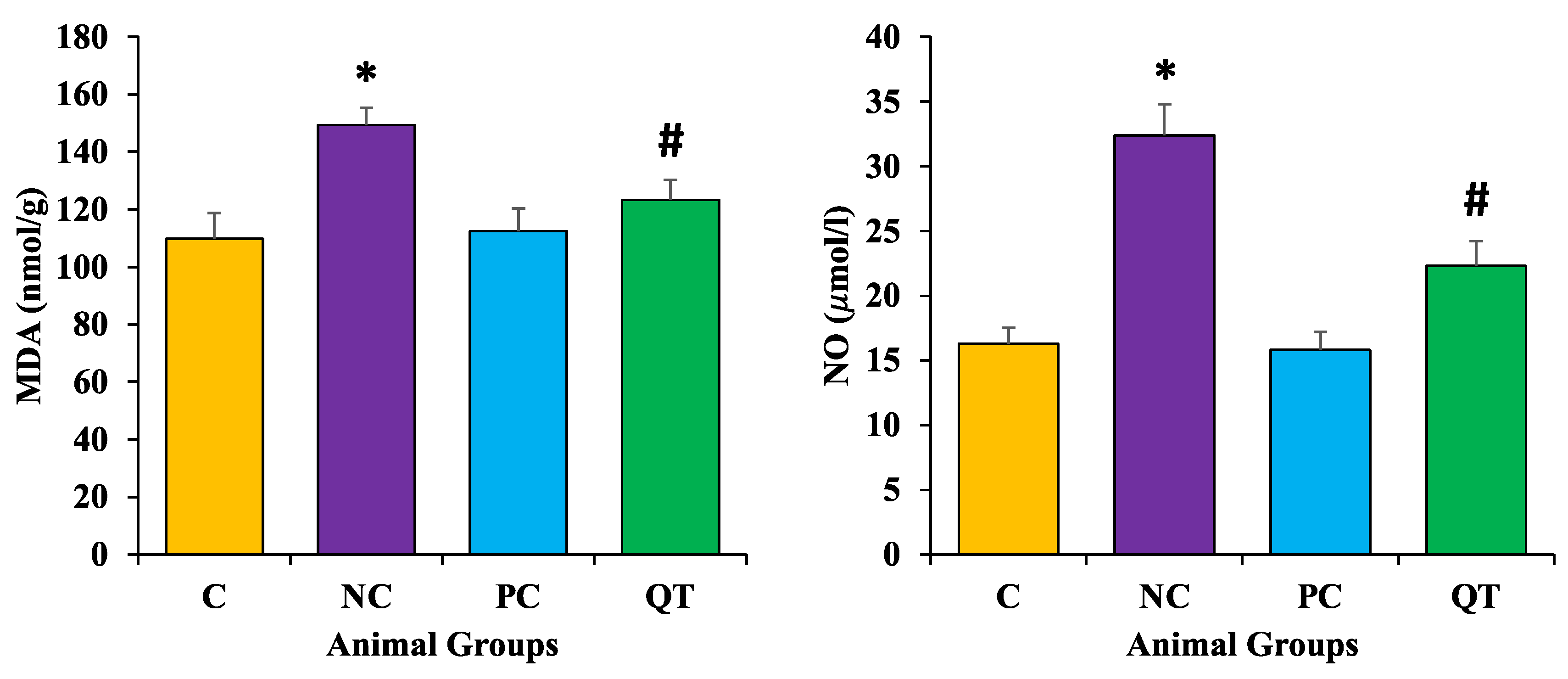{kind=link}
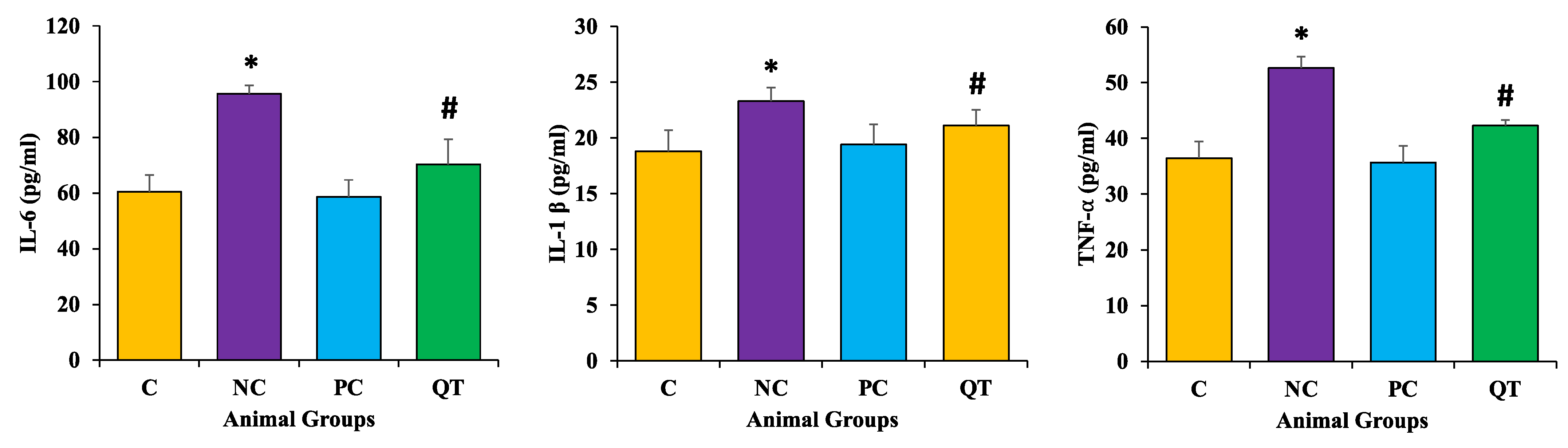{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Groups | Body Weight (0 Days) (g) | Body Weight (After 8 Weeks) (g) |
|---|---|---|
| Control (C) | 216 ± 12 | 295 ± 10 |
| Negative Control (NC) | 214 ± 13 | 198 ± 10 * |
| Positive Control (PC) | 213 ± 11 | 272 ± 12 |
| Quercetin Treatment (QT) | 219 ± 12 | 263 ± 14 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmani, A.H.; Alsahli, M.A.; Khan, A.A.; Almatroodi, S.A. Quercetin, a Plant Flavonol Attenuates Diabetic Complications, Renal Tissue Damage, Renal Oxidative Stress and Inflammation in Streptozotocin-Induced Diabetic Rats. Metabolites 2023, 13, 130. https://doi.org/10.3390/metabo13010130
Rahmani AH, Alsahli MA, Khan AA, Almatroodi SA. Quercetin, a Plant Flavonol Attenuates Diabetic Complications, Renal Tissue Damage, Renal Oxidative Stress and Inflammation in Streptozotocin-Induced Diabetic Rats. Metabolites. 2023; 13(1):130. https://doi.org/10.3390/metabo13010130
Chicago/Turabian StyleRahmani, Arshad Husain, Mohammed A. Alsahli, Amjad Ali Khan, and Saleh A. Almatroodi. 2023. "Quercetin, a Plant Flavonol Attenuates Diabetic Complications, Renal Tissue Damage, Renal Oxidative Stress and Inflammation in Streptozotocin-Induced Diabetic Rats" Metabolites 13, no. 1: 130. https://doi.org/10.3390/metabo13010130
APA StyleRahmani, A. H., Alsahli, M. A., Khan, A. A., & Almatroodi, S. A. (2023). Quercetin, a Plant Flavonol Attenuates Diabetic Complications, Renal Tissue Damage, Renal Oxidative Stress and Inflammation in Streptozotocin-Induced Diabetic Rats. Metabolites, 13(1), 130. https://doi.org/10.3390/metabo13010130