

A Metabolomic Analysis to Assess the Responses of the Male Gonads of Mytilus galloprovincialis after Heavy Metal Exposure

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Exposure of Mussels
2.3. Processing and Sampling of Spermatozoa
2.3.1. Metabolite Sample Processing: Extraction, Purification, and Derivatization
2.3.2. Gas Chromatography–Mass Spectrometry (GC-MS) Metabolomic Profiling
2.3.3. Metabolite Identification
2.4. Statistical Analysis
2.4.1. Profiling Animal Characteristics
2.4.2. In-Depth Metabolomic Data Examination
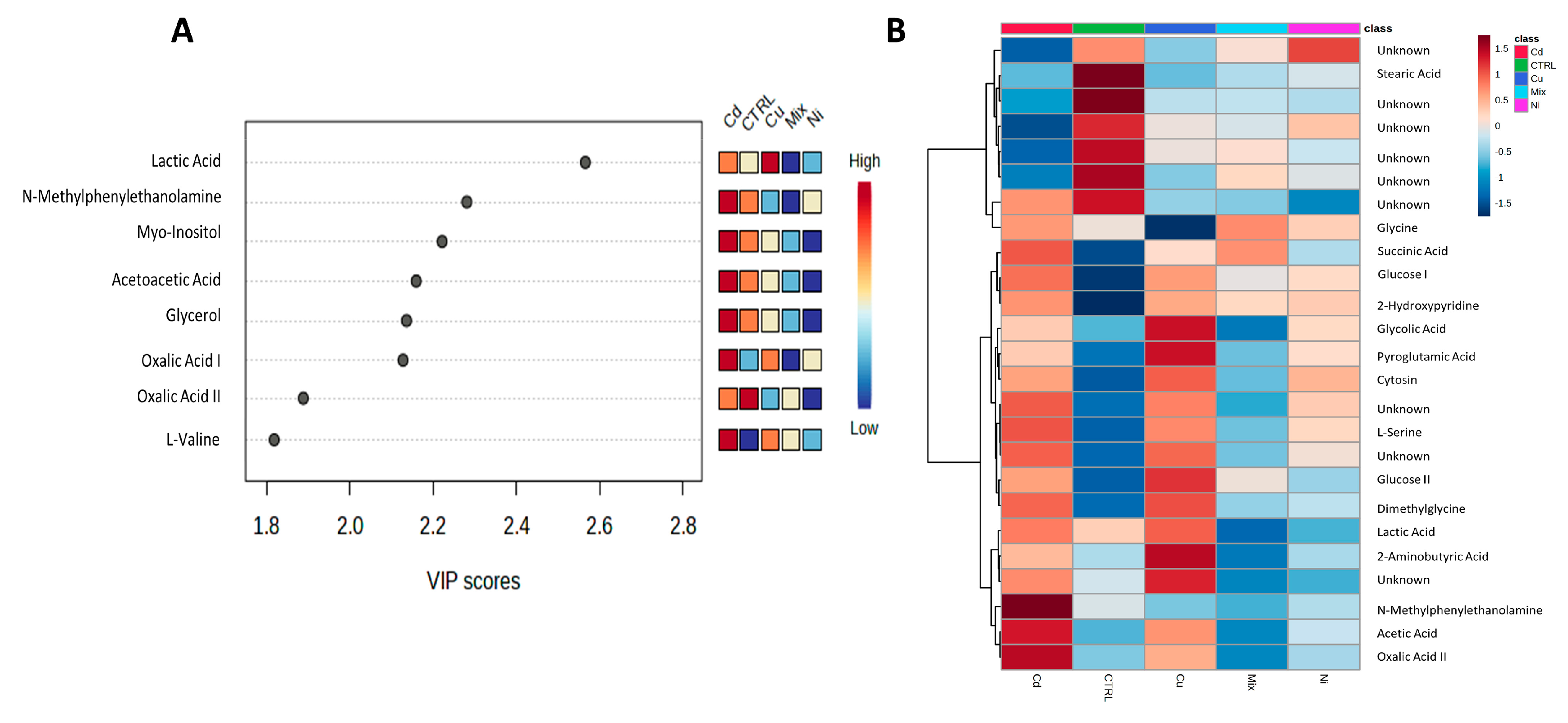
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Xu, H.; Qiu, S.; Wang, X. Cell Metabolomics. OMICS 2013, 17, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-Q.; Chen, H.; Chen, L.; Tang, D.-D.; Miao, H.; Zhao, Y.-Y. Metabolomic Application in Toxicity Evaluation and Toxicological Biomarker Identification of Natural Product. Chem. Biol. Interact. 2016, 252, 114–130. [Google Scholar] [CrossRef]
- Ali, M.M.; Hossain, D.; Al-Imran, A.; Khan, M.S.; Begum, M.; Osman, M.H. Environmental Pollution with Heavy Metals: A Public Health Concern. In Heavy Metals—Their Environmental Impacts and Mitigation; IntechOpen: London, UK, 2021; ISBN 978-1-83968-122-6. [Google Scholar]
- Felici, A.; Bilandžić, N.; Magi, G.E.; Iaffaldano, N.; Fiordelmondo, E.; Doti, G.; Roncarati, A. Evaluation of Long Sea Snail Hinia Reticulata (Gastropod) from the Middle Adriatic Sea as a Possible Alternative for Human Consumption. Foods 2020, 9, 905. [Google Scholar] [CrossRef] [PubMed]
- Perić, L.; Nerlović, V.; Žurga, P.; Žilić, L.; Ramšak, A. Variations of Biomarkers Response in Mussels Mytilus Galloprovincialis to Low, Moderate and High Concentrations of Organic Chemicals and Metals. Chemosphere 2017, 174, 554–562. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus Galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef] [PubMed]
- De Luca-Abbott, S.B.; Richardson, B.J.; McClellan, K.E.; Zheng, G.J.; Martin, M.; Lam, P.K.S. Field Validation of Antioxidant Enzyme Biomarkers in Mussels (Perna viridis) and Clams (Ruditapes philippinarum) Transplanted in Hong Kong Coastal Waters. Mar. Pollut. Bull. 2005, 51, 694–707. [Google Scholar] [CrossRef]
- Gorinstein, S.; Jung, S.-T.; Moncheva, S.; Arancibia-Avila, P.; Park, Y.-S.; Kang, S.-G.; Goshev, I.; Trakhtenberg, S.; Namiesnik, J. Partial Characterization of Proteins from Mussel Mytilus Galloprovincialis as a Biomarker of Contamination. Arch. Environ. Contam. Toxicol. 2005, 49, 504–510. [Google Scholar] [CrossRef]
- Santovito, G.; Piccinni, E.; Cassini, A.; Irato, P.; Albergoni, V. Antioxidant Responses of the Mediterranean Mussel, Mytilus Galloprovincialis, to Environmental Variability of Dissolved Oxygen. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 321–329. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, S.P.; Parakh, S.K.; Tong, Y.W. Health Hazards of Hexavalent Chromium (Cr (VI)) and Its Microbial Reduction. Bioengineered 2022, 13, 4923–4938. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, S.P.; Iqbal, H.M.N.; Tong, Y.W. Omics Approaches in Bioremediation of Environmental Contaminants: An Integrated Approach for Environmental Safety and Sustainability. Environ. Res. 2022, 211, 113102. [Google Scholar] [CrossRef]
- Lin, C.Y.; Viant, M.R.; Tjeerdema, R.S. Metabolomics: Methodologies and Applications in the Environmental Sciences. J. Pestic. Sci. 2006, 31, 245–251. [Google Scholar] [CrossRef]
- Booth, S.C.; Workentine, M.L.; Weljie, A.M.; Turner, R.J. Metabolomics and Its Application to Studying Metal Toxicity. Metallomics 2011, 3, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.; Yeung, W.H.; Craft, J.; Brown, M.; Kennedy, J.; Bignell, J.; Stentiford, G.D.; Viant, M.R. Comparison of Histological, Genetic, Metabolomics, and Lipid-Based Methods for Sex Determination in Marine Mussels. Anal. Biochem. 2007, 369, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Izral, N.M.; Brua, R.B.; Culp, J.M.; Yates, A.G. Crayfish Tissue Metabolomes Effectively Distinguish Impacts of Wastewater and Agriculture in Aquatic Ecosystems. Sci. Total Environ. 2021, 760, 143322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wang, Q.; Li, F.; Cong, M.; Li, L.; Zhao, J.; Liu, D.; et al. Benzo(a)Pyrene-Induced Metabolic Responses in Manila Clam Ruditapes Philippinarum by Proton Nuclear Magnetic Resonance ((1)H NMR) Based Metabolomics. Environ. Toxicol. Pharmacol. 2011, 32, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wu, H.; Li, L.; Zhao, J.; Feng, J.; Yu, J. Metabolic Responses in Gills of Manila Clam Ruditapes Philippinarum Exposed to Copper Using NMR-Based Metabolomics. Mar. Environ. Res. 2011, 72, 33–39. [Google Scholar] [CrossRef]
- Wu, H.; Wang, W.-X. NMR-Based Metabolomic Studies on the Toxicological Effects of Cadmium and Copper on Green Mussels Perna viridis. Aquat. Toxicol. 2010, 100, 339–345. [Google Scholar] [CrossRef]
- Lettieri, G.; Marinaro, C.; Notariale, R.; Perrone, P.; Lombardi, M.; Trotta, A.; Troisi, J.; Piscopo, M. Impact of Heavy Metal Exposure on Mytilus Galloprovincialis Spermatozoa: A Metabolomic Investigation. Metabolites 2023, 13, 943. [Google Scholar] [CrossRef]
- Carbone, G.; Lettieri, G.; Marinaro, C.; Costabile, M.; Notariale, R.; Bianchi, A.R.; De Maio, A.; Piscopo, M. A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus Galloprovincialis: An In Vitro Study. Biomolecules 2023, 13, 520. [Google Scholar] [CrossRef]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus Galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Cavallo, P.; Richards, S.; Symes, S.; Colucci, A.; Sarno, L.; Landolfi, A.; Scala, G.; Adair, D.; Ciccone, C.; et al. Noninvasive Screening for Congenital Heart Defects Using a Serum Metabolomics Approach. Prenat. Diagn. 2021, 41, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Li, J. The Exploration of the Approach to Data Preparation for Chinese Text Analysis Based on R Language. Open Access Libr. J. 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Wehrens, R.; Bloemberg, T.G.; Eilers, P.H.C. Fast Parametric Time Warping of Peak Lists. Bioinformatics 2015, 31, 3063–3065. [Google Scholar] [CrossRef] [PubMed]
- Sysi-Aho, M.; Katajamaa, M.; Yetukuri, L.; Oresic, M. Normalization Method for Metabolomics Data Using Optimal Selection of Multiple Internal Standards. BMC Bioinform. 2007, 8, 93. [Google Scholar] [CrossRef]
- Yi, X.; Xu, Y.; Hu, Q.; Krishnamoorthy, S.; Li, W.; Tang, Z. ASN-SMOTE: A Synthetic Minority Oversampling Method with Adaptive Qualified Synthesizer Selection. Complex. Intell. Syst. 2022, 8, 2247–2272. [Google Scholar] [CrossRef]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; van Ommen, B.; Smilde, A.K. Large-Scale Human Metabolomics Studies: A Strategy for Data (Pre-) Processing and Validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef]
- Gautam, P.K.; Gautam, R.; Banerjee, S.; Chattopadhyaya, M.; Pandey, J. Heavy Metals in the Environment: Fate, Transport, Toxicity and Remediation Technologies. In Heavy Metals: Sources, Toxicity and Remediation Techniques; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2016; pp. 101–130. ISBN 978-1-63484-740-7. [Google Scholar]
- Mousavi, A.; Pourakbar, L.; Siavash Moghaddam, S. Effects of Malic Acid and EDTA on Oxidative Stress and Antioxidant Enzymes of Okra (Abelmoschus esculentus L.) Exposed to Cadmium Stress. Ecotoxicol. Environ. Saf. 2022, 248, 114320. [Google Scholar] [CrossRef]
- Mol, S.; Alakavuk, D.U. Heavy Metals in Mussels (Mytilus galloprovincialis) from Marmara Sea, Turkey. Biol. Trace Elem. Res. 2011, 141, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Li, X.; Petriello, M.C.; Wang, C.; Morris, A.J.; Hennig, B. Application of Metabolomics to Characterize Environmental Pollutant Toxicity and Disease Risks. Rev. Environ. Health 2019, 34, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Boeck, G.D.; Smet, H.D.; Blust, R. The Effect of Sublethal Levels of Copper on Oxygen Consumption and Ammonia Excretion in the Common Carp, Cyprinus Carpio. Aquat. Toxicol. 1995, 32, 127. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.; Li, D.; Xiao, T.; Li, J. Exposure of Silver Carp (Hypophthalmichthys molitrix) to Environmentally Relevant Levels of Cadmium: Hematology, Muscle Physiology, and Implications for Stock Enhancement in the Xiangjiang River (Hunan, China). Sci. China Life Sci. 2013, 56, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Menezes, E.B.; Velho, A.L.C.; Santos, F.; Dinh, T.; Kaya, A.; Topper, E.; Moura, A.A.; Memili, E. Uncovering Sperm Metabolome to Discover Biomarkers for Bull Fertility. BMC Genom. 2019, 20, 714. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.G.; Martins, A.D.; Jarak, I.; Barros, A.; Silva, J.; Sousa, M.; Oliveira, P.F. Testicular Lactate Content Is Compromised in Men with Klinefelter Syndrome. Mol. Reprod. Dev. 2016, 83, 208–216. [Google Scholar] [CrossRef]
- Courtens, J.L.; Plöen, L. Improvement of Spermatogenesis in Adult Cryptorchid Rat Testis by Intratesticular Infusion of Lactate. Biol. Reprod. 1999, 61, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Erkkilä, K.; Aito, H.; Aalto, K.; Pentikäinen, V.; Dunkel, L. Lactate Inhibits Germ Cell Apoptosis in the Human Testis. Mol. Human. Reprod. 2002, 8, 109–117. [Google Scholar] [CrossRef]
- Moorthikumar, K.; Krishnamoorthi, A.; Muthulingam, M. Effect of Nickel Chloride on Lactate Dehydrogenase Activity in Selected Tissues of Fish Labeo Rohita (Hamilton). Eco. Env. Cons. 2020, 26, 643–647. [Google Scholar]
- Vazquez-Levin, M.H.; Verón, G.L. Myo-Inositol in Health and Disease: Its Impact on Semen Parameters and Male Fertility. Andrology 2020, 8, 277–298. [Google Scholar] [CrossRef]
- Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Vitale, S.G.; Laganà, A.S.; Cimino, L.; Calogero, A.E. Myo-Inositol as a Male Fertility Molecule: Speed Them Up! Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 30–35. [Google Scholar] [PubMed]
- De Luca, M.N.; Colone, M.; Gambioli, R.; Stringaro, A.; Unfer, V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants 2021, 10, 1283. [Google Scholar] [CrossRef] [PubMed]
- Benvenga, S.; Micali, A.; Pallio, G.; Vita, R.; Malta, C.; Puzzolo, D.; Irrera, N.; Squadrito, F.; Altavilla, D.; Minutoli, L. Effects of Myo-Inositol Alone and in Combination with Seleno-Lmethionine on Cadmium-Induced Testicular Damage in Mice. Curr. Mol. Pharmacol. 2019, 12, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Fokina, N.N.; Ruokolainen, T.R.; Nemova, N.N.; Bakhmet, I.N. Changes of Blue Mussels Mytilus Edulis L. Lipid Composition under Cadmium and Copper Toxic Effect. Biol. Trace Elem. Res. 2013, 154, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Takahashi, T.; Iguchi, N.; Kitamura, K.; Miyagawa, Y.; Tsujimura, A.; Matsumiya, K.; Okuyama, A.; Nishimune, Y. Ketone Bodies Could Support the Motility but Not the Acrosome Reaction of Mouse Sperm. Int. J. Androl. 2004, 27, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.C.; Hatwalne, V.G. Effect of Acetoacetic Acid on Reduced Glutathione Content of Blood in Rabbits. Nature 1950, 166, 692–693. [Google Scholar] [CrossRef] [PubMed]
- Marinaro, C.; Lettieri, G.; Chianese, T.; Bianchi, A.R.; Zarrelli, A.; Palatucci, D.; Scudiero, R.; Rosati, L.; De Maio, A.; Piscopo, M. Exploring the Molecular and Toxicological Mechanism Associated with Interactions between Heavy Metals and the Reproductive System of Mytilus galloprovincialis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 275, 109778. [Google Scholar] [CrossRef]
- Crisóstomo, L.; Alves, M.G.; Calamita, G.; Sousa, M.; Oliveira, P.F. Glycerol and Testicular Activity: The Good, the Bad and the Ugly. Mol. Hum. Reprod. 2017, 23, 725–737. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, S.; Wang, J.; Liu, Y.; Deng, Y. Roles of High Osmolarity Glycerol and Cell Wall Integrity Pathways in Cadmium Toxicity in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 6169. [Google Scholar] [CrossRef]
- Kayashima, T.; Katayama, T. Oxalic Acid Is Available as a Natural Antioxidant in Some Systems. Biochim. Biophys. Acta 2002, 1573, 1–3. [Google Scholar] [CrossRef]
- Eiss, M.; Schieferstein, G.; Wahl, R. Oxalate in Human Seminal Plasma: Possible Significance and Problems of Oxalate Determination. Fertil. Steril. 2000, 73, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Ugur, M.R. Sperm Functional Genome and Epigenome Regulating Bull Fertility and Sperm Freezability. Ph.D. Thesis, Mississippi State University, Starkville, MS, USA, 2021. [Google Scholar]
- Rashidi, A.; Alizadeh, A.; Sharifi, S.D.; Najafi, A. The Histopathological Changes of Liver and Testis of Japanese Quail Chicks Fed Different Levels of Dietary L-Valine. Vet. Anim. Sci. 2023, 21, 100302. [Google Scholar] [CrossRef] [PubMed]
- Zamir-Nasta, T.; Pazhouhi, M.; Ghanbari, A.; Abdolmaleki, A.; Jalili, C. Expression of Cyclin D1, P21, and Estrogen Receptor Alpha in Aflatoxin G1-Induced Disturbance in Testicular Tissue of Albino Mice. Res. Pharm. Sci. 2021, 16, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, M.; Zhong, Z.; Chen, H.; Wang, M.; Lian, C.; Wang, H.; Zhang, H.; Cao, L.; Li, C. Toxicological Effects of Cadmium on Deep-Sea Mussel Gigantidas Platifrons Revealed by a Combined Proteomic and Metabolomic Approach. Front. Mar. Sci. 2023, 10, 1087411. [Google Scholar] [CrossRef]
- Ji, C.; Wei, L.; Zhao, J.; Wu, H. Metabolomic Analysis Revealed That Female Mussel Mytilus Galloprovincialis Was Sensitive to Bisphenol A Exposures. Environ. Toxicol. Pharmacol. 2014, 37, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus Galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Ricciardiello, M.; Palumbo, G.; Troisi, J. Selectivity of Metal Bioaccumulation and Its Relationship with Glutathione S-Transferase Levels in Gonadal and Gill Tissues of Mytilus Galloprovincialis Exposed to Ni (II), Cu (II) and Cd (II). Rend. Fis. Acc. Lincei 2016, 27, 737–748. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lettieri, G.; Marinaro, C.; Brogna, C.; Montano, L.; Lombardi, M.; Trotta, A.; Troisi, J.; Piscopo, M. A Metabolomic Analysis to Assess the Responses of the Male Gonads of Mytilus galloprovincialis after Heavy Metal Exposure. Metabolites 2023, 13, 1168. https://doi.org/10.3390/metabo13121168
Lettieri G, Marinaro C, Brogna C, Montano L, Lombardi M, Trotta A, Troisi J, Piscopo M. A Metabolomic Analysis to Assess the Responses of the Male Gonads of Mytilus galloprovincialis after Heavy Metal Exposure. Metabolites. 2023; 13(12):1168. https://doi.org/10.3390/metabo13121168
Chicago/Turabian StyleLettieri, Gennaro, Carmela Marinaro, Carlo Brogna, Luigi Montano, Martina Lombardi, Alessio Trotta, Jacopo Troisi, and Marina Piscopo. 2023. "A Metabolomic Analysis to Assess the Responses of the Male Gonads of Mytilus galloprovincialis after Heavy Metal Exposure" Metabolites 13, no. 12: 1168. https://doi.org/10.3390/metabo13121168
APA StyleLettieri, G., Marinaro, C., Brogna, C., Montano, L., Lombardi, M., Trotta, A., Troisi, J., & Piscopo, M. (2023). A Metabolomic Analysis to Assess the Responses of the Male Gonads of Mytilus galloprovincialis after Heavy Metal Exposure. Metabolites, 13(12), 1168. https://doi.org/10.3390/metabo13121168