
Pregnancy Toxemia in Ewes: A Review of Molecular Metabolic Mechanisms and Management Strategies



Abstract
:1. Introduction
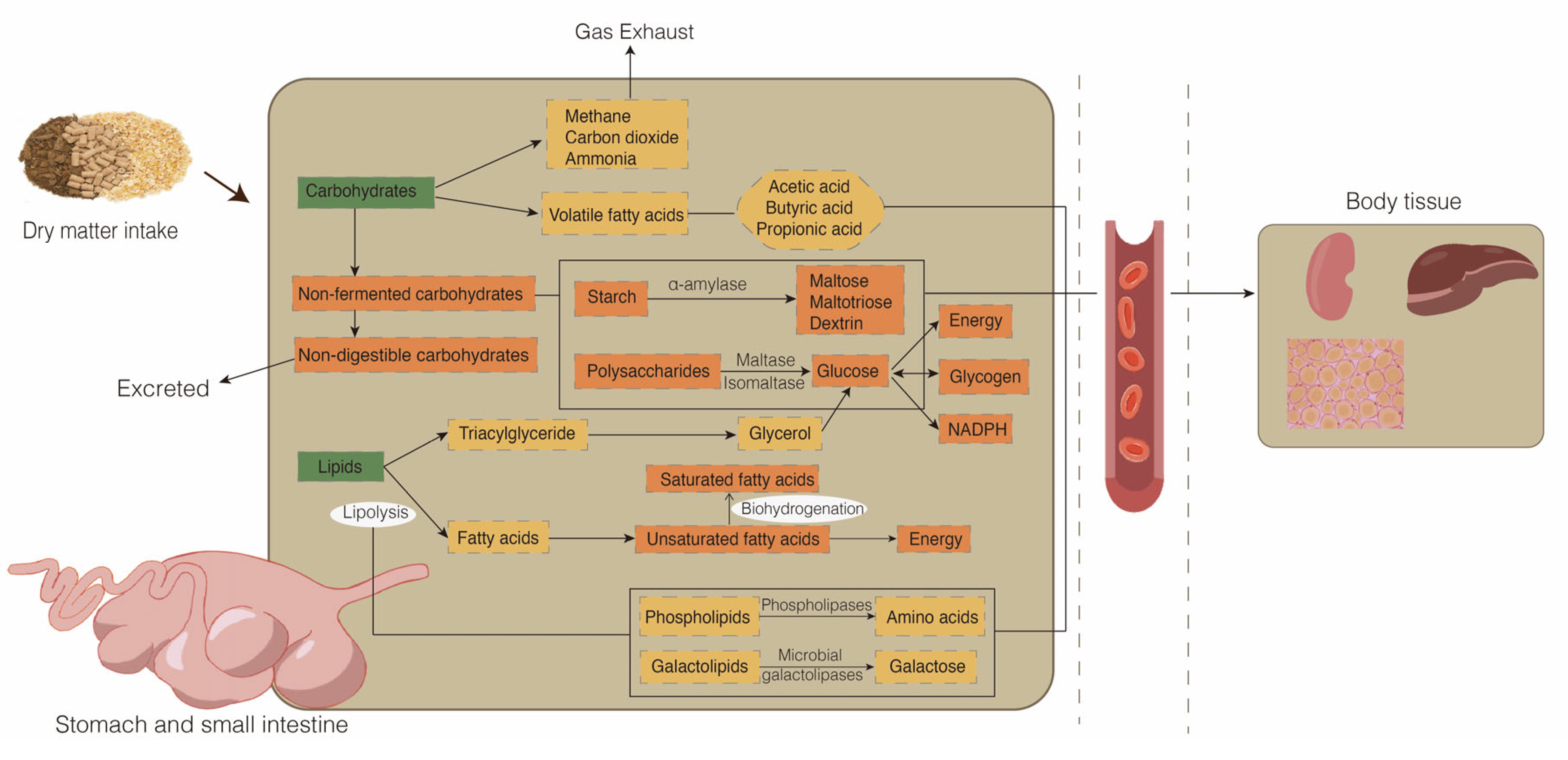
2. Glycolipid Metabolism in Ruminants
2.1. Glucose Metabolism in Ruminants
2.2. Lipid Metabolism in Ruminants
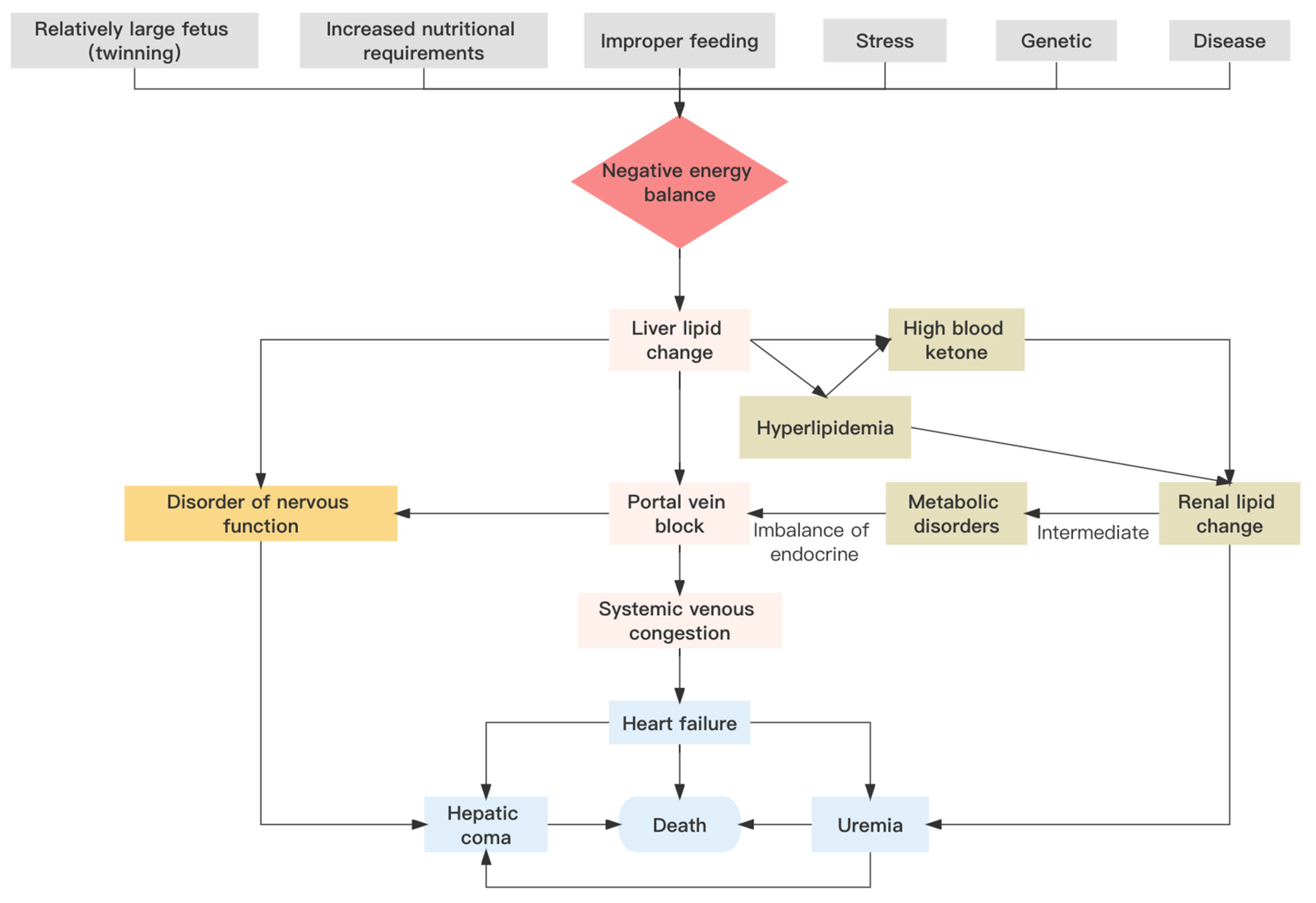
3. Causes of Pregnancy Toxemia
3.1. Nutrition and Digestion
3.2. Genetics
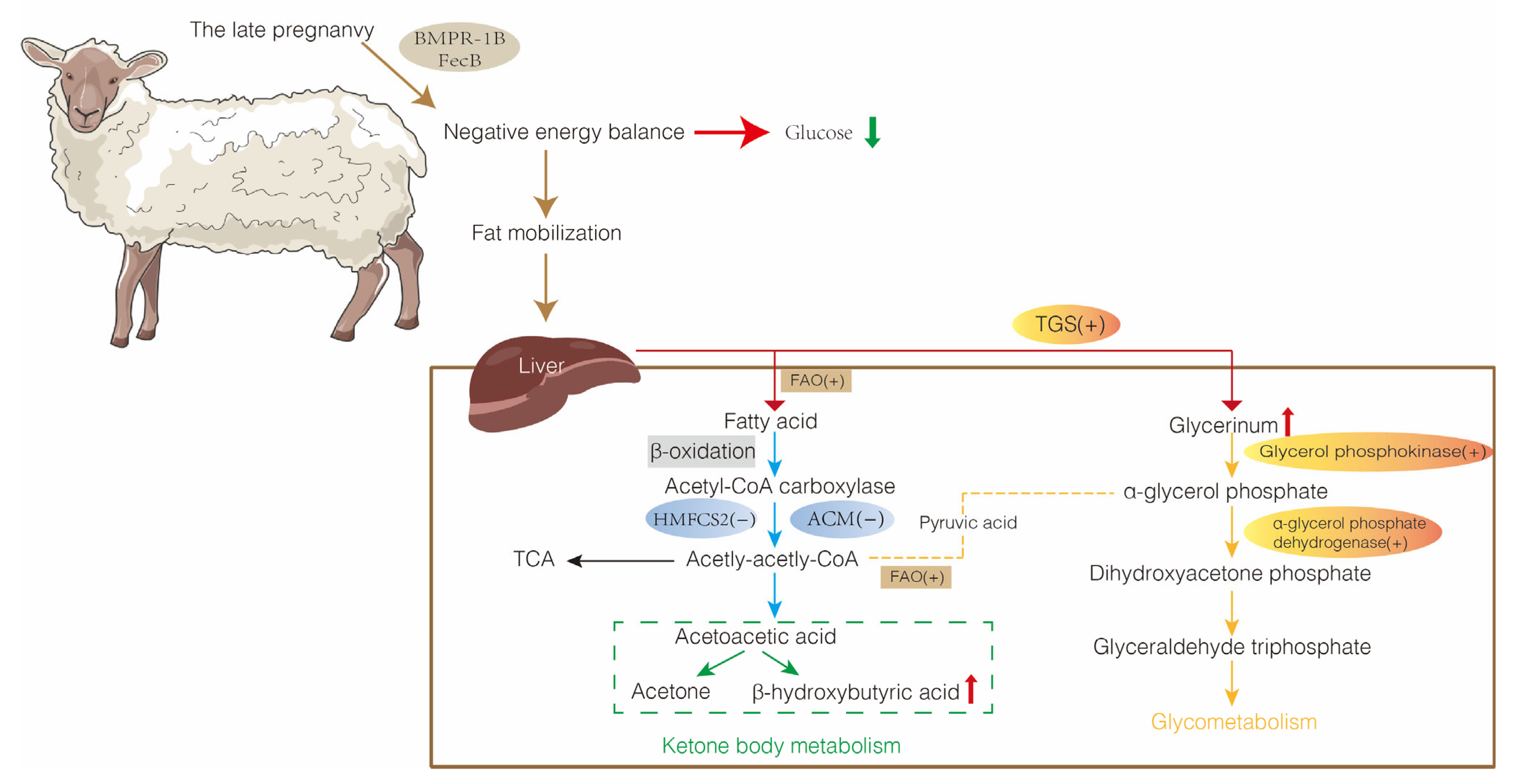
4. Lipid Metabolism-Related Molecular Mechanisms in Pregnancy Toxemia
4.1. Fatty Acid Oxidation-Related Genes
4.2. Acetyl Coenzyme Metabolism-Related Genes
4.3. Triglyceride Synthesis and Related Genes
5. Management Strategies for Pregnancy Toxemia
5.1. Diagnosis of Pregnancy Toxemia
5.1.1. Histological Changes of Liver
5.1.2. Biochemical Parameters in the Blood
5.2. Treatment of Pregnancy Toxemia
5.3. Prevention of Pregnancy Toxemia
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rook, J.S. Pregnancy Toxemia of Ewes, Does, and Beef Cows. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 293–317. [Google Scholar] [CrossRef] [PubMed]
- Kemp, M.W.; Musk, G.C.; Saito, M. Animal Models for the Study of Infection-Associated Preterm Birth. In Animal Models for the Study of Human Disease; Conn, P.M., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 863–888. [Google Scholar]
- Xue, Y.; Guo, C.; Hu, F.; Zhu, W.; Mao, S. Maternal Undernutrition Induces Fetal Hepatic Lipid Metabolism Disorder and Affects the Development of Fetal Liver in a Sheep Model. FASEB J. 2019, 33, 9990–10004. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Hu, F.; Guo, C.; Mei, S.; Xie, F.; Zeng, H.; Mao, S. Undernutrition Shifted Colonic Fermentation and Digest-Associated Bacterial Communities in Pregnant Ewes. Appl. Microbiol. Biotechnol. 2020, 104, 5973–5984. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Xue, Y.; Guo, C.; Liu, J.; Mao, S. The Response of Ruminal Fermentation, Epithelium-Associated Microbiota, and Epithelial Barrier Function to Severe Feed Restriction in Pregnant Ewes. J. Anim. Sci. 2018, 96, 4293–4305. [Google Scholar] [CrossRef] [PubMed]
- Bulgin, M. Diseases of the Periparturient Ewe. In Current Therapy in Large Animal Theriogenology, 2nd ed.; Youngquist, R.S., Threlfall, W.R., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2007; pp. 695–700. [Google Scholar]
- Edmondson, M.A.; Pugh, D.G. Pregnancy Toxemia in Sheep and Goats. In Food Animal Practice, 5th ed.; Anderson, D.E., Rings, D.M., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2009; pp. 144–145. [Google Scholar]
- Schlumbohm, C.; Harmeyer, J. Twin-Pregnancy Increases Susceptibility of Ewes to Hypoglycaemic Stress and Pregnancy Toxaemia. Res. Vet. Sci. 2008, 84, 286–299. [Google Scholar] [CrossRef]
- Duehlmeier, R.; Fluegge, I.; Schwert, B.; Ganter, M. Post-Glucose Load Changes of Plasma Key Metabolite and Insulin Concentrations during Pregnancy and Lactation in Ewes with Different Susceptibility to Pregnancy Toxaemia. J. Anim. Physiol. Anim. Nutr. 2013, 97, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Kempf, K.; Röhling, M.; Lenzen-Schulte, M.; Schloot, N.C.; Martin, S. Ketone Bodies: From Enemy to Friend and Guardian Angel. BMC Med. 2021, 19, 313. [Google Scholar] [CrossRef]
- Lorenz, M.D.; Coates, J.R.; Kent, M. Systemic or Multifocal Signs. In Handbook of Veterinary Neurology, 5th ed.; Lorenz, M.D., Coates, J.R., Kent, M., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2011; pp. 432–487. [Google Scholar]
- Vasava, P.R.; Jani, R.G.; Goswami, H.V.; Rathwa, S.D.; Tandel, F.B. Studies on Clinical Signs and Biochemical Alteration in Pregnancy Toxemic Goats. Vet. World 2016, 9, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Owens, F.N.; Zinn, R.A.; Kim, Y.K. Limits to Starch Digestion in the Ruminant Small Intestine. J. Anim. Sci 1986, 63, 1634–1648. [Google Scholar] [CrossRef]
- Huntington, G.B. Starch Utilization by Ruminants: From Basics to the Bunk. J. Anim. Sci. 1997, 75, 852–867. [Google Scholar] [CrossRef]
- Harmon, D.L.; Yamka, R.M.; Elam, N.A. Factors Affecting Intestinal Starch Digestion in Ruminants: A Review. Can. J. Anim. Sci. 2004, 84, 309–318. [Google Scholar] [CrossRef]
- Nichols, B.L.; Quezada-Calvillo, R.; Robayo-Torres, C.C.; Ao, Z.; Hamaker, B.R.; Butte, N.F.; Marini, J.; Jahoor, F.; Sterchi, E.E. Mucosal Maltase-Glucoamylase Plays a Crucial Role in Starch Digestion and Prandial Glucose Homeostasis of Mice. J. Nutr. 2009, 139, 684–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urrutia, O.; Mendizabal, J.A.; Alfonso, L.; Soret, B.; Insausti, K.; Arana, A. Adipose Tissue Modification through Feeding Strategies and Their Implication on Adipogenesis and Adipose Tissue Metabolism in Ruminants. Int. J. Mol. Sci. 2020, 21, 3183. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, T.I.R.C.; Chen, Y.; Furusho-Garcia, I.F.; Perez, J.R.O.; Hopkins, D.L. Manipulation of Omega-3 PUFAs in Lamb: Phenotypic and Genotypic Views. Compr. Rev. Food Sci. Food Saf. 2015, 14, 189–204. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, Rumen Biohydrogenation and Nutritional Quality of Cow and Goat Milk Fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Hocquette, J.F.; Bauchart, D. Intestinal Absorption, Blood Transport and Hepatic and Muscle Metabolism of Fatty Acids in Preruminant and Ruminant Animals. Reprod. Nutr. Dev. 1999, 39, 27–48. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.S.; Pascoal, R.A.; Stilwell, G.T. Glycaemia as a Sign of the Viability of the Foetuses in the Last Days of Gestation in Dairy Goats with Pregnancy Toxaemia. Ir. Vet. J. 2012, 65, 1. [Google Scholar] [CrossRef] [Green Version]
- Duehlmeier, R.; Noldt, S.; Ganter, M. Pancreatic Insulin Release and Peripheral Insulin Sensitivity in German Black Headed Mutton and Finish Landrace Ewes: Evaluation of the Role of Insulin Resistance in the Susceptibility to Ovine Pregnancy Toxemia. Domest. Anim. Endocrinol. 2013, 44, 213–221. [Google Scholar] [CrossRef]
- Kalyesubula, M.; Rosov, A.; Alon, T.; Moallem, U.; Dvir, H. Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial. Animals 2019, 9, 731. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Guo, C.; Hu, F.; Liu, J.; Mao, S. Hepatic Metabolic Profile Reveals the Adaptive Mechanisms of Ewes to Severe Undernutrition during Late Gestation. Metabolites 2018, 8, 85. [Google Scholar] [CrossRef]
- Sargison, N.D.; Scott, P.R.; Penny, C.D.; Pirie, R.S.; Kelly, J.M. Plasma Enzymes and Metabolites as Potential Prognostic Indices of Ovine Pregnancy Toxaemia--a Preliminary Study. Br. Vet. J. 1994, 150, 271–277. [Google Scholar] [CrossRef]
- Van Saun, R.J. Pregnancy Toxemia in a Flock of Sheep. J. Am. Vet. Med. Assoc. 2000, 217, 1536–1539. [Google Scholar] [CrossRef]
- Al-Qudah, K.M. Oxidant and Antioxidant Profile of Hyperketonemic Ewes Affected by Pregnancy Toxemia. Vet. Clin. Pathol. 2011, 40, 60–65. [Google Scholar] [CrossRef]
- Moallem, U.; Rozov, A.; Gootwine, E.; Honig, H. Plasma Concentrations of Key Metabolites and Insulin in Late-Pregnant Ewes Carrying 1 to 5 Fetuses. J. Anim. Sci. 2012, 90, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Han, S.B. Correlation of HMGCS2 and DAGLA Genes with Toxemia of Pregnancy in Sheep. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2017. [Google Scholar]
- Barakat, I.A.; Salem, L.M.; Daoud, N.M.; Khalil, W.K.; Mahrous, K.F. Genetic Polymorphism of Candidate Genes for Fecundity Traits in Egyptian Sheep Breeds. Biomed. Res. 2017, 28, 851–857. [Google Scholar]
- El-Seedy, A.S.; Hashem, N.M.; El-Azrak, K.M.; Nour El-Din, A.; Ramadan, T.A.; Taha, T.A.; Salem, M.H. Genetic Screening of FecB, FecXG and FecXI Mutations and Their Linkage with Litter Size in Barki and Rahmani Sheep Breeds. Reprod. Domest. Anim. 2017, 52, 1133–1137. [Google Scholar] [CrossRef]
- Mohammad, M.K.; Seyed, H.H.; Nosratollah, H. Genetic Polymorphism BMP15 and GDF9 Genes in Sangsari Sheep of Iran. Int. J. Genet. Mol. Biol. 2011, 3, 31–34. [Google Scholar]
- Adkinson, A.Y.; Adkinson, R.W. The FecB (Booroola) Gene and Implications for the Turkish Sheep Industry. Turk. J. Vet. Anim. Sci. 2013, 37, 621–624. [Google Scholar] [CrossRef]
- Lan, Z.J.; Gu, P.; Xu, X.; Jackson, K.J.; DeMayo, F.J.; O’Malley, B.W.; Cooney, A.J. GCNF-Dependent Repression of BMP-15 and GDF-9 Mediates Gamete Regulation of Female Fertility. EMBO J. 2003, 22, 4070–4081. [Google Scholar] [CrossRef] [Green Version]
- Regan, S.L.P.; Knight, P.G.; Yovich, J.L.; Leung, Y.; Arfuso, F.; Dharmarajan, A. Involvement of Bone Morphogenetic Proteins (BMP) in the Regulation of Ovarian Function. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: New York, NY, USA, 2018; Volume 107, pp. 227–261. [Google Scholar]
- Zheng, J.; Wang, Z.; Yang, H.; Yao, X.; Yang, P.; Ren, C.; Wang, F.; Zhang, Y. Pituitary Transcriptomic Study Reveals the Differential Regulation of LncRNAs and MRNAs Related to Prolificacy in Different FecB Genotyping Sheep. Genes 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, S.; Li, F.; Pan, X.; Li, C.; Zhang, X.; Ma, Y.; La, Y.; Xi, R.; Li, T. Polymorphisms of the Ovine BMPR-IB, BMP-15 and FSHR and Their Associations with Litter Size in Two Chinese Indigenous Sheep Breeds. Int. J. Mol. Sci. 2015, 16, 11385–11397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNatty, K.P.; Heath, D.A.; Hudson, N.L.; Lun, S.; Juengel, J.L.; Moore, L.G. Gonadotrophin-Responsiveness of Granulosa Cells from Bone Morphogenetic Protein 15 Heterozygous Mutant Sheep. Reproduction 2009, 138, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Guerra, A.; Wiltbank, M.C.; Battista, S.E.; Kirkpatrick, B.W.; Sartori, R. Mechanisms Regulating Follicle Selection in Ruminants: Lessons Learned from Multiple Ovulation Models. Anim. Reprod. 2018, 15, 660–679. [Google Scholar] [CrossRef]
- Strauss, J.F.; Williams, C.J. Ovarian Life Cycle. In Yen and Jaffe’s Reproductive Endocrinology, 8th ed.; Strauss, J.F., Barbieri, R.L., Eds.; Elsevier: Philadelphia, PA, USA, 2019; pp. 167–205. [Google Scholar]
- Xue, Y.; Guo, C.; Hu, F.; Zhu, W.; Mao, S. PPARA/RXRA Signalling Regulates the Fate of Hepatic Non-Esterified Fatty Acids in a Sheep Model of Maternal Undernutrition. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158548. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, Y.; Zhang, Y.; Long, M.; Guo, Y.; Wang, Z.; Li, X.; Zhang, C.; Li, X.; He, J.; et al. Effect of Non-Esterified Fatty Acids on Fatty Acid Metabolism-Related Genes in Calf Hepatocytes Cultured in Vitro. Cell. Physiol. Biochem. 2013, 32, 1509–1516. [Google Scholar] [CrossRef]
- Young, P.A.; Senkal, C.E.; Suchanek, A.L.; Grevengoed, T.J.; Lin, D.D.; Zhao, L.; Crunk, A.E.; Klett, E.L.; Füllekrug, J.; Obeid, L.M.; et al. Long-Chain Acyl-CoA Synthetase 1 Interacts with Key Proteins That Activate and Direct Fatty Acids into Niche Hepatic Pathways. J. Biol. Chem. 2018, 293, 16724–16740. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Liu, G.; Du, X.; Shi, Z.; Jin, M.; Sha, X.; Li, X.; Wang, Z.; Li, X. Expression Patterns of Hepatic Genes Involved in Lipid Metabolism in Cows with Subclinical or Clinical Ketosis. J. Dairy Sci. 2019, 102, 1725–1735. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Yang, X.F.; Liu, H.L.; Fu, N.; Ouyang, Y.; Qing, K. Long-Chain Acyl-CoA Synthetase in Fatty Acid Metabolism Involved in Liver and Other Diseases: An Update. World J. Gastroenterol. 2015, 21, 3492–3498. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of Metabolism and as Therapeutic Targets in Cardiovascular Disease. Part I: PPAR-α. Future Cardiol. 2017, 13, 259–278. [Google Scholar] [CrossRef]
- Tahri-Joutey, M.; Andreoletti, P.; Surapureddi, S.; Nasser, B.; Cherkaoui-Malki, M.; Latruffe, N. Mechanisms Mediating the Regulation of Peroxisomal Fatty Acid Beta-Oxidation by PPARα. Int. J. Mol. Sci. 2021, 22, 8969. [Google Scholar] [CrossRef] [PubMed]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR Control of Metabolism and Cardiovascular Functions. Nat. Rev. Cardiol. 2021, 18, 809–823. [Google Scholar] [CrossRef]
- Li, T.; Li, X.; Meng, H.; Chen, L.; Meng, F. ACSL1 Affects Triglyceride Levels through the PPARγ Pathway. Int. J. Med. Sci. 2020, 17, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Liang, K.; Zhao, S.; Jia, W.; Liu, Y.; Wu, H.; Lv, J.; Cao, C.; Chen, T.; Zhuang, S.; et al. Chemoproteomics Reveals Baicalin Activates Hepatic CPT1 to Ameliorate Diet-Induced Obesity and Hepatic Steatosis. Proc. Natl. Acad. Sci. USA 2018, 115, E5896–E5905. [Google Scholar] [CrossRef] [Green Version]
- Huh, J.Y.; Reilly, S.M.; Abu-Odeh, M.; Murphy, A.N.; Mahata, S.K.; Zhang, J.; Cho, Y.; Seo, J.B.; Hung, C.-W.; Green, C.R.; et al. TANK-Binding Kinase 1 Regulates the Localization of Acyl-CoA Synthetase ACSL1 to Control Hepatic Fatty Acid Oxidation. Cell Metab. 2020, 32, 1012–1027. [Google Scholar] [CrossRef]
- Pietrocola, F.; Galluzzi, L.; Bravo-San Pedro, J.M.; Madeo, F.; Kroemer, G. Acetyl Coenzyme A: A Central Metabolite and Second Messenger. Cell Metab. 2015, 21, 805–821. [Google Scholar] [CrossRef] [Green Version]
- Puchalska, P.; Crawford, P.A. Multi-Dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Tozzi, R.; Risi, R.; Tuccinardi, D.; Mariani, S.; Basciani, S.; Spera, G.; Lubrano, C.; Gnessi, L. Beneficial Effects of the Ketogenic Diet on Nonalcoholic Fatty Liver Disease: A Comprehensive Review of the Literature. Obes. Rev. 2020, 21, e13024. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.J.; Peng, L.; Cline, G.W.; Petersen, K.F.; Shulman, G.I. A Non-Invasive Method to Assess Hepatic Acetyl-CoA in Vivo. Cell Metab. 2017, 25, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, P.C. Acetyl-CoA Metabolism and Histone Acetylation in the Regulation of Aging and Lifespan. Antioxidants 2021, 10, 572. [Google Scholar] [CrossRef]
- Pesántez-Pacheco, J.L.; Heras-Molina, A.; Torres-Rovira, L.; Sanz-Fernández, M.V.; García-Contreras, C.; Vázquez-Gómez, M.; Feyjoo, P.; Cáceres, E.; Frías-Mateo, M.; Hernández, F.; et al. Influence of Maternal Factors (Weight, Body Condition, Parity, and Pregnancy Rank) on Plasma Metabolites of Dairy Ewes and Their Lambs. Animals 2019, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Li, X.B.; Fu, S.X.; Wu, C.C.; Wang, X.X.; Yu, G.J.; Long, M.; Wang, Z.; Liu, G.W. Alterations of Fatty Acid β-Oxidation Capability in the Liver of Ketotic Cows. J. Dairy Sci. 2012, 95, 1759–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Li, X.; Li, D.; Li, Y.; Song, Y.; Deng, Q.; Wang, J.; Zhang, Y.; Ding, H.; Yin, L.; et al. β-Hydroxybutyrate Activates the NF-ΚB Signaling Pathway to Promote the Expression of pro-Inflammatory Factors in Calf Hepatocytes. Cell. Physiol. Biochem. 2014, 33, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-Induced Ketosis Alters Metabolic and Signaling Gene Networks in Liver of Periparturient Dairy Cows. Physiol. Genom. 2007, 32, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asif, S.; Kim, R.Y.; Fatica, T.; Sim, J.; Zhao, X.; Oh, Y.; Denoncourt, A.; Cheung, A.C.; Downey, M.; Mulvihill, E.E.; et al. Hmgcs2-Mediated Ketogenesis Modulates High-Fat Diet-Induced Hepatosteatosis. Mol. Metab. 2022, 61, 101494. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.T.; Li, C.; Weiss, H.L.; Zhou, Y.; Liu, C.; Wang, Q.; Evers, B.M. Regulation of Ketogenic Enzyme HMGCS2 by Wnt/β-Catenin/PPARγ Pathway in Intestinal Cells. Cells 2019, 8, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Zou, D.; Liu, R.; Shi, S.; Du, J.; Tian, M.; Wang, X.; Hou, M.; Duan, Z.; Ma, Y. BHBA Regulates the Expressions of Lipid Synthesis and Oxidation Genes in Sheep Hepatocytes through the AMPK Pathway. Res. Vet. Sci. 2021, 140, 153–163. [Google Scholar] [CrossRef]
- Liu, S.; Jing, F.; Yu, C.; Gao, L.; Qin, Y.; Zhao, J. AICAR-Induced Activation of AMPK Inhibits TSH/SREBP-2/HMGCR Pathway in Liver. PLoS ONE 2015, 10, e0124951. [Google Scholar] [CrossRef]
- Huang, C.; Gao, X.; Shi, Y.; Guo, L.; Zhou, C.; Li, N.; Chen, W.; Yang, F.; Li, G.; Zhuang, Y.; et al. Inhibition of Hepatic AMPK Pathway Contributes to Free Fatty Acids-Induced Fatty Liver Disease in Laying Hen. Metabolites 2022, 12, 825. [Google Scholar] [CrossRef]
- Arfianti, A.; Pok, S.; Barn, V.; Haigh, W.G.; Yeh, M.M.; Ioannou, G.N.; Teoh, N.C.-H.; Farrell, G.C. Exercise Retards Hepatocarcinogenesis in Obese Mice Independently of Weight Control. J. Hepatol. 2020, 73, 140–148. [Google Scholar] [CrossRef]
- Xue, Y.F.; Guo, C.Z.; Hu, F.; Sun, D.M.; Liu, J.H.; Mao, S.Y. Molecular Mechanisms of Lipid Metabolism Disorder in Livers of Ewes with Pregnancy Toxemia. Animal 2019, 13, 992–999. [Google Scholar] [CrossRef]
- Coleman, R.A.; Lewin, T.M.; Muoio, D.M. Physiological and Nutritional Regulation of Enzymes of Triacylglycerol Synthesis. Annu. Rev. Nutr. 2000, 20, 77–103. [Google Scholar] [CrossRef]
- Neschen, S.; Morino, K.; Hammond, L.E.; Zhang, D.; Liu, Z.X.; Romanelli, A.J.; Cline, G.W.; Pongratz, R.L.; Zhang, X.-M.; Choi, C.S.; et al. Prevention of Hepatic Steatosis and Hepatic Insulin Resistance in Mitochondrial Acyl-CoA:Glycerol-Sn-3-Phosphate Acyltransferase 1 Knockout Mice. Cell Metab. 2005, 2, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [PubMed]
- Bhatt-Wessel, B.; Jordan, T.W.; Miller, J.H.; Peng, L. Role of DGAT Enzymes in Triacylglycerol Metabolism. Arch. Biochem. Biophys. 2018, 655, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Cases, S.; Jensen, D.R.; Chen, H.C.; Sande, E.; Tow, B.; Sanan, D.A.; Raber, J.; Eckel, R.H.; Farese, R.V. Obesity Resistance and Multiple Mechanisms of Triglyceride Synthesis in Mice Lacking Dgat. Nat. Genet. 2000, 25, 87–90. [Google Scholar] [CrossRef]
- Stone, S.J.; Myers, H.M.; Watkins, S.M.; Brown, B.E.; Feingold, K.R.; Elias, P.M.; Farese, R.V. Lipopenia and Skin Barrier Abnormalities in DGAT2-Deficient Mice. J. Biol. Chem. 2004, 279, 11767–11776. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Choi, S.H.; Li, Q.; Wang, Y.; Sun, B.; Tang, L.; Wang, E.Z.; Hua, H.; Li, X.Z. Overexpression of DGAT2 Stimulates Lipid Droplet Formation and Triacylglycerol Accumulation in Bovine Satellite Cells. Animals 2022, 12, 1847. [Google Scholar] [CrossRef]
- Basu, D.; Goldberg, I.J. Regulation of Lipoprotein Lipase-Mediated Lipolysis of Triglycerides. Curr. Opin. Lipidol. 2020, 31, 154–160. [Google Scholar] [CrossRef]
- Kristensen, K.K.; Leth-Espensen, K.Z.; Mertens, H.D.T.; Birrane, G.; Meiyappan, M.; Olivecrona, G.; Jørgensen, T.J.D.; Young, S.G.; Ploug, M. Unfolding of Monomeric Lipoprotein Lipase by ANGPTL4: Insight into the Regulation of Plasma Triglyceride Metabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 4337–4346. [Google Scholar] [CrossRef]
- Wu, S.A.; Kersten, S.; Qi, L. Lipoprotein Lipase and Its Regulators: An Unfolding Story. Trends Endocrinol. Metab. 2021, 32, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Role and Mechanism of the Action of Angiopoietin-like Protein ANGPTL4 in Plasma Lipid Metabolism. J. Lipid Res. 2021, 62, 100150. [Google Scholar] [CrossRef]
- Sylvers-Davie, K.L.; Davies, B.S.J. Regulation of Lipoprotein Metabolism by ANGPTL3, ANGPTL4, and ANGPTL8. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E493–E508. [Google Scholar] [CrossRef] [PubMed]
- Mandard, S.; Zandbergen, F.; van Straten, E.; Wahli, W.; Kuipers, F.; Müller, M.; Kersten, S. The Fasting-Induced Adipose Factor/Angiopoietin-like Protein 4 Is Physically Associated with Lipoproteins and Governs Plasma Lipid Levels and Adiposity. J. Biol. Chem. 2006, 281, 934–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitler, K.M.; Shetty, S.K.; Cushing, E.M.; Sylvers-Davie, K.L.; Davies, B.S.J. Regulation of Plasma Triglyceride Partitioning by Adipose-Derived ANGPTL4 in Mice. Sci. Rep. 2021, 11, 7873. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, K. An Updated ANGPTL3-4-8 Model as a Mechanism of Triglyceride Partitioning between Fat and Oxidative Tissues. Prog. Lipid Res. 2022, 85, 101140. [Google Scholar] [CrossRef]
- Haller, J.F.; Mintah, I.J.; Shihanian, L.M.; Stevis, P.; Buckler, D.; Alexa-Braun, C.A.; Kleiner, S.; Banfi, S.; Cohen, J.C.; Hobbs, H.H.; et al. ANGPTL8 Requires ANGPTL3 to Inhibit Lipoprotein Lipase and Plasma Triglyceride Clearance. J. Lipid Res. 2017, 58, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Adam, R.C.; Mintah, I.J.; Alexa-Braun, C.A.; Shihanian, L.M.; Lee, J.S.; Banerjee, P.; Hamon, S.C.; Kim, H.I.; Cohen, J.C.; Hobbs, H.H.; et al. Angiopoietin-like Protein 3 Governs LDL-Cholesterol Levels through Endothelial Lipase-Dependent VLDL Clearance. J. Lipid Res. 2020, 61, 1271–1286. [Google Scholar] [CrossRef]
- Wen, Y.; Chen, Y.Q.; Konrad, R.J. The Regulation of Triacylglycerol Metabolism and Lipoprotein Lipase Activity. Adv. Biol. 2022, 6, e2200093. [Google Scholar] [CrossRef]
- Su, X.; Peng, H.; Chen, X.; Wu, X.; Wang, B. Hyperlipidemia and Hypothyroidism. Clin. Chim. Acta 2022, 527, 61–70. [Google Scholar] [CrossRef]
- Bai, Y.; Lu, D.; Qu, D.; Li, Y.; Zhao, N.; Cui, G.; Li, X.; Sun, X.; Liu, Y.; Wei, M.; et al. The Role of ANGPTL Gene Family Members in Hepatocellular Carcinoma. Dis. Markers 2022, 2022, 1844352. [Google Scholar] [CrossRef]
- Yang, J.; Song, Q.-Y.; Niu, S.-X.; Chen, H.-J.; Petersen, R.B.; Zhang, Y.; Huang, K. Emerging Roles of Angiopoietin-like Proteins in Inflammation: Mechanisms and Potential as Pharmacological Targets. J. Cell. Physiol. 2022, 237, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Souto, R.J.C.; Afonso, J.A.B.; Mendonça, C.L.; Dantas, A.F.M.; Cajueiro, J.F.P.; Gonçalves, D.N.A.; Olivera, E.F.; Soares, P.C. Biochemical, Endocrine, and Histopathological Profile of Liver and Kidneys of Sheep with Pregnancy Toxemia. Pesq. Vet. Bras. 2019, 39, 780–788. [Google Scholar] [CrossRef]
- Wang, M.; Li, E.; Wang, G. Histopathological Analysis of Liver During Pregnancy Toxemia in Small-tailed Han Sheep. Agri. Sci. Technol. 2014, 15, 470–473. [Google Scholar]
- Lima, M.S.; Cota, J.B.; Vaz, Y.M.; Ajuda, I.G.; Pascoal, R.A.; Carolino, N.; Hjerpe, C.A. Glucose Intolerance in Dairy Goats with Pregnancy Toxemia: Lack of Correlation between Blood PH and Beta Hydroxybutyric Acid Values. Can. Vet. J. 2016, 57, 635–640. [Google Scholar]
- Iqbal, R.; Beigh, S.A.; Mir, A.Q.; Shaheen, M.; Hussain, S.A.; Nisar, M.; Dar, A.A. Evaluation of Metabolic and Oxidative Profile in Ovine Pregnancy Toxemia and to Determine Their Association with Diagnosis and Prognosis of Disease. Trop. Anim. Health Prod. 2022, 54, 338. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.S.; Silveira, J.M.; Carolino, N.; Lamas, L.P.; Pascoal, R.A.; Hjerpe, C.A. Usefulness of Clinical Observations and Blood Chemistry Values for Predicting Clinical Outcomes in Dairy Goats with Pregnancy Toxaemia. Ir. Vet. J. 2016, 69, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, D.E.; Bedford, S.; Darby, S.; Palmisano, M.; MacKay, R.J.; Renaud, D.L. Acid-base Disorders in Sick Goats and Their Association with Mortality: A Simplified Strong Ion Difference Approach. J. Vet. Intern. Med. 2020, 34, 2776–2786. [Google Scholar] [CrossRef] [PubMed]
- Pregnancy Toxemia in Sheep and Goats—Metabolic Disorders. Available online: https://www.merckvetmanual.com/metabolic-disorders/hepatic-lipidosis/pregnancy-toxemia-in-sheep-and-goats (accessed on 9 January 2023).
- Brozos, C.; Mavrogianni, V.S.; Fthenakis, G.C. Treatment and Control of Peri-Parturient Metabolic Diseases: Pregnancy Toxemia, Hypocalcemia, Hypomagnesemia. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 105–113. [Google Scholar] [CrossRef]
- Gomez, D.E.; Kuthiala, S.; Liu, H.L.; Durosier, D.L.; Cao, M.; Burns, P.; Desrochers, A.; Fecteau, G.; Frasch, M.G. Effect of Maternal Ketoacidosis on the Ovine Fetus. Can. Vet. J. 2015, 56, 863–866. [Google Scholar] [PubMed]
- Andrade, I.M.; Simões, P.B.A.; Lamas, L.P.; Carolino, N.; Lima, M.S. Blood Lactate, PH, Base Excess and PCO2 as Prognostic Indicators in Caesarean-Born Kids from Goats with Pregnancy Toxaemia. Ir. Vet. J. 2019, 72, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayan, M.; El-abedeen, A.E.Z.; Ellah, M.F.A. Some Metabolic Parameters during Transition Period in Dairy Cows with and without Retained Fetal Membranes. J. Adv. Vet. Res. 2019, 9, 45–48. [Google Scholar]
- Constable, P.D.; Hinchcliff, K.W.; Done, S.H.; Grünberg, W.; Radostits, O.M. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 11th ed.; Elsevier: St. Louis, MO, USA, 2017. [Google Scholar]
- Malkan, U.Y.; Gunes, G.; Corakci, A. Rational Diagnoses of Diabetes: The Comparison of 1,5-Anhydroglucitol with Other Glycemic Markers. Springerplus 2015, 4, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doré, V.; Dubuc, J.; Bélanger, A.M.; Buczinski, S. Definition of Prepartum Hyperketonemia in Dairy Goats. J. Dairy Sci. 2015, 98, 4535–4543. [Google Scholar] [CrossRef]
- Marteniuk, J.V.; Herdt, T.H. Pregnancy Toxemia and Ketosis of Ewes and Does. Vet. Clin. N. Am. Food Anim. Pract. 1988, 4, 307–315. [Google Scholar] [CrossRef]
- Bani Ismail, Z.A.; Al-Majali, A.M.; Amireh, F.; Al-Rawashdeh, O.F. Metabolic Profiles in Goat Does in Late Pregnancy with and without Subclinical Pregnancy Toxemia. Vet. Clin. Pathol. 2008, 37, 434–437. [Google Scholar] [CrossRef]
- Lynch, G.P.; Jackson, C., Jr. A Method for Assessing the Nutritional Status of Gestating Ewes. Can. J. Anim. Sci. 1983, 63, 603–611. [Google Scholar] [CrossRef]
- Cal-Pereyra, L.; González-Montaña, J.R.; Benech, A.; Acosta-Dibarrat, J.; Martín, M.; Perini, S.; Abreu, M.; Da Silva, S.; Rodríguez, P. Evaluation of Three Therapeutic Alternatives for the Early Treatment of Ovine Pregnancy Toxaemia. Ir. Vet. J. 2015, 68, 25. [Google Scholar] [CrossRef] [Green Version]
- Oetzel, G.R. Monitoring and Testing Dairy Herds for Metabolic Disease. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 651–674. [Google Scholar] [CrossRef]
- Panousis, N.; Brozos, C.; Karagiannis, I.; Giadinis, N.D.; Lafi, S.; Kritsepi-Konstantinou, M. Evaluation of Precision Xceed® Meter for On-Site Monitoring of Blood β-Hydroxybutyric Acid and Glucose Concentrations in Dairy Sheep. Res. Vet. Sci. 2012, 93, 435–439. [Google Scholar] [CrossRef]
- Araújo, C.A.S.C.; Minervino, A.H.H.; Sousa, R.S.; Oliveira, F.L.C.; Rodrigues, F.A.M.L.; Mori, C.S.; Ortolani, E.L. Validation of a Handheld β-Hydroxybutyrate Acid Meter to Identify Hyperketonaemia in Ewes. PeerJ 2020, 8, e8933. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.K.; Gately, R.E.; Kellogg, T.D.; Zinn, S.A.; Govoni, K.E.; Reed, S.A. Evaluation of the Nova Vet Meter for Sheep-Side Monitoring of β-Hydroxybutyric Acid (BHBA) and Description of Ewe BHBA during Late Gestation in Three Flocks from the Northeastern U.S. Res. Vet. Sci. 2018, 118, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.M.; Mendonça, C.L.; Assis, R.N.; Filho, E.F.O.; Soares, G.S.L.; Souto, R.J.C.; Soares, P.C.; Afonso, J.A.B. Changes in Cardiac Biomarkers in Goats Naturally Affected by Pregnancy Toxemia. Res. Vet. Sci. 2020, 130, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Alon, T.; Rosov, A.; Lifshitz, L.; Dvir, H.; Gootwine, E.; Moallem, U. The Distinctive Short-Term Response of Late-Pregnant Prolific Ewes to Propylene Glycol or Glycerol Drenching. J. Dairy Sci. 2020, 103, 10245–10257. [Google Scholar] [CrossRef]
- Da Silva, S.; Cal-Pereyra, L.G.; Benech, A.; Acosta-Dibarrat, J.; Martin, M.J.; Abreu, M.C.; Perini, S.; González-Montaña, J.R. Evaluation of a Fibrate, Specific Stimulant of PPARα, as a Therapeutic Alternative to the Treatment of Clinical Ovine Pregnancy Toxaemia. J. Vet. Pharmacol. Ther. 2016, 39, 497–503. [Google Scholar] [CrossRef]
- Martín-Alonso, M.J.; Escalera-Valente, F.; Cal-Pereyra, L.G.; Benech, A.; Alonso, M.P.; González-Montaña, J.R. Energetic Metabolism in Fasting Sheep: Regularization of Metabolic Profile by Treatment with Oral Glucose, with Prior Handling of Gastric Groove. Rev. Bras. Zootec. 2019, 48, e20180290. [Google Scholar] [CrossRef]
- Zamir, S.; Rozov, A.; Gootwine, E. Treatment of Pregnancy Toxaemia in Sheep with Flunixin Meglumine. Vet. Rec. 2009, 165, 265–266. [Google Scholar] [CrossRef]
- Kang, D.Y.; Sp, N.; Do Park, K.; Lee, H.K.; Song, K.D.; Yang, Y.M. Silibinin Inhibits in Vitro Ketosis by Regulating HMGCS2 and NF-KB: Elucidation of Signaling Molecule Relationship under Ketotic Conditions. Vitr. Cell Dev. Biol. Anim. 2019, 55, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Liu, C.L.; Chiu, W.C.; Twu, Y.C.; Liao, Y.J. HMGCS2 Mediates Ketone Production and Regulates the Proliferation and Metastasis of Hepatocellular Carcinoma. Cancers 2019, 11, 1876. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.Y.; SP, N.; Darvin, P.; Joung, Y.H.; Byun, H.J.; Do, C.H.; Park, K.D.; Park, M.N.; Cho, K.H.; Yang, Y.M. Momilactone B Inhibits Ketosis In Vitro by Regulating the ANGPTL3-LPL Pathway and Inhibiting HMGCS2. Anim. Biotechnol. 2017, 28, 189–197. [Google Scholar] [CrossRef]
- Hunt, E.R. Treatment of Pregnancy Toxaemia in Ewes by Induction of Parturition. Aust. Vet. J. 1976, 52, 338–339. [Google Scholar] [CrossRef]
- Lima, M.; Pascoal, R.A.; Stilwell, G.; Hjerpe, C.A. Clinical Findings, Blood Chemistry Values, and Epidemiologic Data from Dairy Goats with Pregnancy Toxemia. Bov. Pract. 2012, 46, 102–110. [Google Scholar]
- Menzies, P.I. Lambing Management and Neonatal Care. In Current Therapy in Large Animal Theriogenology, 2nd ed.; Youngquist, R.S., Threlfall, W.R., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2007; pp. 680–695. [Google Scholar]
- Harrison, F.A. Dexamethasone-Induced Parturition in Sheep. Br. Vet. J. 1982, 138, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sir, C.; Bartlewski, P.M. Analyses of Parental and Seasonal Influences on the Synchrony of Dexamethasone-Induced Lambing and Lamb Characteristics. Livest. Sci. 2010, 131, 119–124. [Google Scholar] [CrossRef]
- Özalp, G.R.; Yavuz, A.; Seker, I.; Udum-Küçükşen, D.; Rişvanlı, A.; Korlu, Y. Evaluation of an Alternative Treatment Protocol by Aglepristone to Induce Parturition in Ewes with an Experimental Model of Early Pregnancy Toxemia. Theriogenology 2018, 116, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Özalp, R.G.; Yavuz, A.; Orman, A.; Seker, İ.; Udum Küçükşen, D.; Rişvanlı, A.; Demiral, Ö.O.; Wehrend, A. Parturition Induction in Ewes by a Progesterone Receptor Blocker, Aglepristone, and Subsequent Neonatal Survival: Preliminary Results. Theriogenology 2017, 87, 141–147. [Google Scholar] [CrossRef]
- Fthenakis, G.C.; Arsenos, G.; Brozos, C.; Fragkou, I.A.; Giadinis, N.D.; Giannenas, I.; Mavrogianni, V.S.; Papadopoulos, E.; Valasi, I. Health Management of Ewes during Pregnancy. Anim. Reprod. Sci. 2012, 130, 198–212. [Google Scholar] [CrossRef]
- Sejian, V.; Silpa, M.V.; Nair, M.R.; Devaraj, C.; Krishnan, G.; Bagath, M.; Chauhan, S.S.; Suganthi, R.U.; Fonseca, V.F.C.; König, S.; et al. Heat Stress and Goat Welfare: Adaptation and Production Considerations. Animals 2021, 11, 1021. [Google Scholar] [CrossRef]
- Kasimanickam, R.K. Subclinical Pregnancy Toxemia-Induced Gene Expression Changes in Ovine Placenta and Uterus. Front. Vet. Sci. 2016, 3, 69. [Google Scholar] [CrossRef] [Green Version]
- Qing, G.L.T. Causes, Differential Diagnosis and Prevention of Pregnancy Toxaemia in Sheep. China Anim. Health 2021, 23, 40. [Google Scholar]
- Lacetera, N.; Bernabucci, U.; Ronchi, B.; Nardone, A. Effects of Subclinical Pregnancy Toxemia on Immune Responses in Sheep. Am. J. Vet. Res. 2001, 62, 1020–1024. [Google Scholar] [CrossRef]
- Silva, S.R.; Sacarrão-Birrento, L.; Almeida, M.; Ribeiro, D.M.; Guedes, C.; Montaña, J.R.G.; Pereira, A.F.; Zaralis, K.; Geraldo, A.; Tzamaloukas, O.; et al. Extensive Sheep and Goat Production: The Role of Novel Technologies towards Sustainability and Animal Welfare. Animals 2022, 12, 885. [Google Scholar] [CrossRef] [PubMed]
- Fazio, E.; Bionda, A.; Chiofalo, V.; Crepaldi, P.; Lopreiato, V.; Medica, P.; Liotta, L. Adaptive Responses of Thyroid Hormones, Insulin, and Glucose during Pregnancy and Lactation in Dairy Cows. Animals 2022, 12, 1395. [Google Scholar] [CrossRef] [PubMed]
- Polizel, D.M.; Susin, I.; Gentil, R.S.; Ferreira, E.M.; de Souza, R.A.; Freire, A.P.A.; Pires, A.V.; Ferraz, M.V.C.; Rodrigues, P.H.M.; Eastridge, M.L. Crude Glycerin Decreases Nonesterified Fatty Acid Concentration in Ewes during Late Gestation and Early Lactation. J. Anim. Sci. 2017, 95, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Barimanloo, A.M.; Chalmeh, A.; Pourjafar, M.; Mirzaei, A. Effects of Intravenous Butaphosphan and Cyanocobalamin to Late Pregnant Ewes on the Metabolic Indices around Parturition and Weight Gain of Their Lambs after Birth. Vet. Med. Sci. 2021, 8, 781–791. [Google Scholar] [CrossRef]
- Jia, X.J.; Tang, J.H.; Zhang, K.; Tu, Y.L.; Gao, L.P.; Bai, Y.F. Progress in Nutritional Regulation of Pregnancy Toxemia in Perinatal Ewes. Feed Res. 2020, 43, 121–124. [Google Scholar]
- Araújo, C.A.S.C.; Sousa, R.S.; Monteiro, B.M.; Oliveira, F.L.C.; Minervino, A.H.H.; Rodrigues, F.A.M.L.; Vale, R.G.; Mori, C.S.; Ortolani, E.L. Potential Prophylactic Effect of Recombinant Bovine Somatotropin (RbST) in Sheep with Experimentally Induced Hyperketonemia. Res. Vet. Sci. 2018, 119, 215–220. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Description | Designation | Location |
|---|---|---|
| Fatty acid oxidation | Long-chain acyl-CoA synthase 1 (ASCL1) | Chromosome 3 (172306676..172309463) |
| Long-chain acyl-CoA synthase 2 (ASCL2) | Chromosome 21 (45685760..45687628) | |
| Long-chain acyl-CoA dehydrogenase (ACADL) | Chromosome 2 (211860043..211904014, complement) | |
| Carnitine palmitoyltransferase 1B (CPT1B) | Chromosome 3 (225815614..225823875, complement) | |
| Carnitine palmitoyl transferase 1A (CPT1A) | Chromosome 21 (42553789..42613314, complement) | |
| Hydroxyacyl-CoA dehydrogenase (HADH) | Chromosome 6 (17565219..17608867, complement) | |
| Enolyl-CoA hydratase (ECH1) | Chromosome 14 (47913667..47922968, complement) | |
| Acetyl-CoA acyltransferase 1 (ACAA1) | Chromosome 23 (49746617..49779436, complement) | |
| Acetyl-CoA acyltransferase 2 (ACAA2) | Chromosome 19 (11608660..11633147, complement) | |
| Acetyl coenzyme metabolism | 3-Hydroxy-3-methylglutaryl-CoA (HMG-CoA) | |
| Hydroxy methylglutaryl-CoA synthase 1 (HMGCS1) | Chromosome 16 (31543986..31566698) | |
| Hydroxy methylglutaryl-CoA synthase 2 (HMGCS2) | Chromosome 1 (97112935..97137264, complement) | |
| Hydroxy methylglutaryl-CoA lyase (HMGCL) | Chromosome 2 (242831761..242851036) | |
| Hydroxy methylglutaryl-CoA reductases (HMGCR) | Chromosome 7 (6749627..6771079) | |
| Triglyceride synthesis | Glycerol kinase (GK) | Chromosome X (29849127..29924685) |
| Glycerol-3-phosphate dehydrogenase 1 like (GPD1L) | Chromosome 19 (6617541..6675362) | |
| Glycerol-3-phosphate acyltransferase (GPA) | Chromosome 22 (32219904..32288784, complement) | |
| Diacylglycerol acyltransferase 1 (DGAT1) | Chromosome 9 (13560420..13569123) | |
| Diacylglycerol acyltransferase 2 (DGAT2) | Chromosome 15 (53943498..53976554) |
| Parameters | Pregnancy Toxemia | Reference Range | References |
|---|---|---|---|
| PH | ↓ | 7.32–7.5 | [92] |
| HCO3− (mmol/L) | ↓ | 20–29 | [92] |
| BE (mmol/L) | ↓ | −5–+4 | [92] |
| pCO2 (mmHg) | ↓ | 38–45 | [92] |
| Glucose (mg/dL) | ↓ | 50–80 | [93] |
| NEFA (mmol/L) | ↑ | <0.400 | [93] |
| BHBA (mmol/L) | ↑ | <0.600 | [94] |
| Fructosamine (mmol/L) | ↓ | 1.25–1.36 | [93] |
| Albumin (g/dL) | ↓ | 2.4–3.0 | [93] |
| Cholesterol (mg/dL) | ↑ | 52–76 | [93] |
| Triglyceride (mg/dL) | ↑ | 12.22 | [93] |
| AST (U/L) | ↑ | 60–280 | [93] |
| GGT (U/L) | ↑ | 20–52 | [93] |
| BUN (mmol/L) | ↓ | 8–20 | [93] |
| LDH (U/L) | ↑ | 238–440 | [93] |
| CK (U/L) | ↑ | 8.1–12.9 | [93] |
| Potassium (mEq/L) | ↓ | 4.7–7.1 | [93] |
| Calcium (mEq/L) | ↓ | 11.5–12.8 | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Liu, N.; Wang, Y.; Ding, K.; Huang, S.; Zhang, C. Pregnancy Toxemia in Ewes: A Review of Molecular Metabolic Mechanisms and Management Strategies. Metabolites 2023, 13, 149. https://doi.org/10.3390/metabo13020149
Ji X, Liu N, Wang Y, Ding K, Huang S, Zhang C. Pregnancy Toxemia in Ewes: A Review of Molecular Metabolic Mechanisms and Management Strategies. Metabolites. 2023; 13(2):149. https://doi.org/10.3390/metabo13020149
Chicago/Turabian StyleJi, Xiaoyu, Ning Liu, Yuqin Wang, Ke Ding, Shucheng Huang, and Cai Zhang. 2023. "Pregnancy Toxemia in Ewes: A Review of Molecular Metabolic Mechanisms and Management Strategies" Metabolites 13, no. 2: 149. https://doi.org/10.3390/metabo13020149
APA StyleJi, X., Liu, N., Wang, Y., Ding, K., Huang, S., & Zhang, C. (2023). Pregnancy Toxemia in Ewes: A Review of Molecular Metabolic Mechanisms and Management Strategies. Metabolites, 13(2), 149. https://doi.org/10.3390/metabo13020149