
A Four-Step Platform to Optimize Growth Conditions for High-Yield Production of Siderophores in Cyanobacteria

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Anabaena flos Aquae Genome Analyses
2.3. Bacterial Cultures
2.4. Cloning of Iuc-GFP Vector
2.5. Spectrofluorimetric Measurement of GFP Expression
2.6. LC-HRMS and Molecular Networking of the Extracts
3. Results
3.1. Identification of the Promoter Region of the Operon Iuc in Anabaena Flos Aquae
3.2. Construction of the Reporter Vector for Iuc Promoter Activity
3.3. Screening of the Environmental Conditions Promoting Iuc Promoter Activation
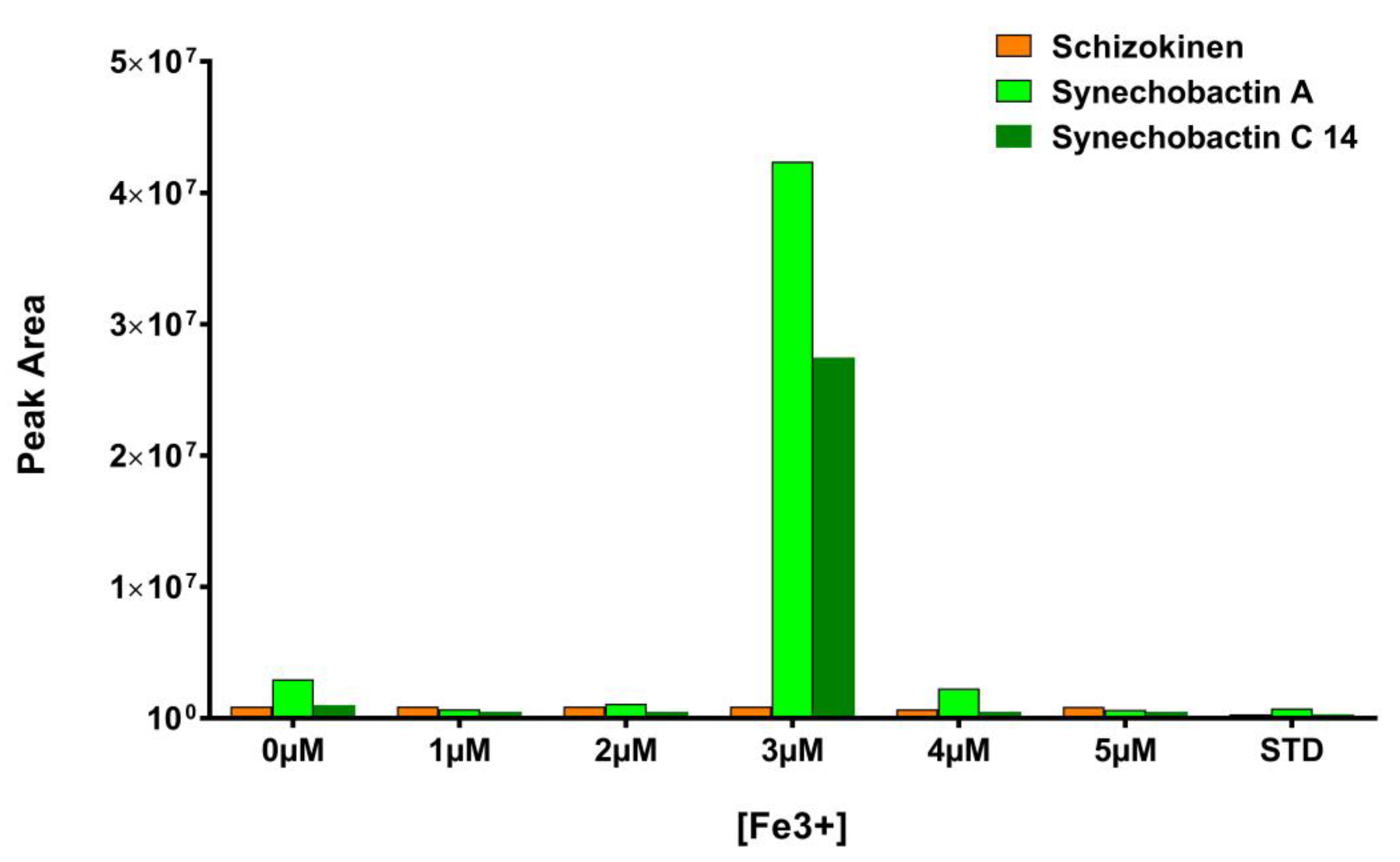
3.4. Optimization of Culture Conditions of Anabaena Flos-Aquae UTEX1444 for Siderophore Production
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [Green Version]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder Web-Services for Easy Identification of Acquired Antibiotic Resistance and E. Coli Virulence Genes in Bacteriophage and Prophage Nucleotide Sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.-B.; Lou, W.-J.; Ke, W.-T.; Song, W.-Y.; Price, N.M.; Qiu, B.-S. New Insights into Iron Acquisition by Cyanobacteria: An Essential Role for ExbB-ExbD Complex in Inorganic Iron Uptake. ISME J. 2015, 9, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.; Straus, N.A. Iron Deprivation in Cyanobacteria. J. Appl. Phycol. 1994, 6, 199–210. [Google Scholar] [CrossRef]
- Årstøl, E.; Hohmann-Marriott, M.F. Cyanobacterial Siderophores—Physiology, Structure, Biosynthesis, and Applications. Mar. Drugs 2019, 17, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Singh, P.; Srivastava, A. Synthesis, Nature and Utility of Universal Iron Chelator-Siderophore: A Review. Microbiol. Res. 2018, 212–213, 103–111. [Google Scholar] [CrossRef]
- Miethke, M.; Marahiel, M.A. Siderophore-Based Iron Acquisition and Pathogen Control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [Green Version]
- Leventhal, G.E.; Ackermann, M.; Schiessl, K.T. Why Microbes Secrete Molecules to Modify Their Environment: The Case of Iron-Chelating Siderophores. J. R. Soc. Interface 2019, 16, 20180674. [Google Scholar] [CrossRef]
- De Serrano, L.O. Biotechnology of Siderophores in High-Impact Scientific Fields. Biomol. Concepts 2017, 8, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Day, J.A.; Cohen, S.M. Investigating the Selectivity of Metalloenzyme Inhibitors. J. Med. Chem. 2013, 56, 7997–8007. [Google Scholar] [CrossRef]
- Emri, T.; Tóth, V.; Nagy, C.T.; Nagy, G.; Pócsi, I.; Gyémánt, G.; Antal, K.; Balla, J.; Balla, G.; Román, G.; et al. Towards High-Siderophore-Content Foods: Optimisation of Coprogen Production in Submerged Cultures of Penicillium Nalgiovense: Foods with High Siderophore Contents. J. Sci. Food Agric. 2013, 93, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Pócsi, I.; Jeney, V.; Kertai, P.; Pócsi, I.; Emri, T.; Gyémánt, G.; Fésüs, L.; Balla, J.; Balla, G. Fungal Siderophores Function as Protective Agents of LDL Oxidation and Are Promising Anti-Atherosclerotic Metabolites in Functional Food. Mol. Nutr. Food Res. 2008, 52, 1434–1447. [Google Scholar] [CrossRef]
- Tonziello, G.; Caraffa, E.; Pinchera, B.; Granata, G.; Petrosillo, N. Present and Future of Siderophore-Based Therapeutic and Diagnostic Approaches in Infectious Diseases. Infect. Dis. Rep. 2019, 11, 8208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, E.; Holmström, S.J.M. Siderophores in Environmental Research: Roles and Applications: Siderophores in Environmental Research. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.; Patry, S.; Dobson, A. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- ACUF—Algal Collection University Federico II—Naples, I. Anabaena Flos-Aquae Healey. Available online: http://www.Acuf.Net/Index.Php?Option=com_content&view=article&id=43:Anabaena-Flos-Aquae-Healey&catid=2&Itemid=127&lang=en&Itemid=113 (accessed on 10 July 2022).
- UTEX. UTEX 1444 Anabaena Flos-Aquae. Available online: Https://Utex.Org/Products/Utex-1444?Variant=30991204089946 (accessed on 1 November 2022).
- BioCyc. Summary of Trichormus Variabilis, Strain ATCC 29413, Version 26.1, Tier 3 Uncurated Database. Available online: Https://Biocyc.Org/Organism-Summary?Object=GCF_000204075 (accessed on 15 August 2022).
- Esposito, G.; Teta, R.; Marrone, R.; De Sterlich, C.; Casazza, M.; Anastasio, A.; Lega, M.; Costantino, V. A Fast Detection Strategy for Cyanobacterial Blooms and Associated Cyanotoxins (FDSCC) Reveals the Occurrence of Lyngbyatoxin A in Campania (South Italy). Chemosphere 2019, 225, 342–351. [Google Scholar] [CrossRef]
- Medicine, N.L.o. Anabaena Variabilis ATCC 29413, Complete Genome. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Nuccore/CP000117.1 (accessed on 1 November 2022).
- Salamov, V.; Salamov, A. Automatic Annotation of Microbial Genomes and Metagenomic Sequences. In Metagenomics and Its Applications in Agriculture, Biomedicine and Environmental Studies; Nova Science Publishers: Hauppauge, NY, USA, 2020. [Google Scholar]
- Prediction, B.-B.P. Anabaena Variabilis. Available online: Http://Www.Bacpp.Bioinfoucs.Com/Home (accessed on 17 August 2022).
- Riccio, G.; Bottone, S.; La Regina, G.; Badolati, N.; Passacantilli, S.; Rossi, G.B.; Accardo, A.; Dentice, M.; Silvestri, R.; Novellino, E.; et al. A Negative Allosteric Modulator of WNT Receptor Frizzled 4 Switches into an Allosteric Agonist. Biochemistry 2018, 57, 839–851. [Google Scholar] [CrossRef]
- Oliviero, G.; Stornaiuolo, M.; D’Atri, V.; Nici, F.; Yousif, A.M.; D’Errico, S.; Piccialli, G.; Mayol, L.; Novellino, E.; Marinelli, L.; et al. Screening Platform toward New Anti-HIV Aptamers Set on Molecular Docking and Fluorescence Quenching Techniques. Anal. Chem. 2016, 88, 2327–2334. [Google Scholar] [CrossRef]
- Britstein, M.; Devescovi, G.; Handley, K.M.; Malik, A.; Haber, M.; Saurav, K.; Teta, R.; Costantino, V.; Burgsdorf, I.; Gilbert, J.A.; et al. A New N-Acyl Homoserine Lactone Synthase in an Uncultured Symbiont of the Red Sea Sponge Theonella Swinhoei. Appl. Environ. Microbiol. 2016, 82, 1274–1285. [Google Scholar] [CrossRef] [Green Version]
- Katajamaa, M.; Miettinen, J.; Oresic, M. MZmine: Toolbox for Processing and Visualization of Mass Spectrometry Based Molecular Profile Data. Bioinformatics 2006, 22, 634–636. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Teta, R.; Esposito, G.; De Sterlich, C.; Lega, M.; Costantino, V. Early Detection of Cyanobacterial Blooms and Associated Cyanotoxins Using Fast Detection Strategy. J. Vis. Exp. 2021, 168, e61889. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Teta, R.; Esposito, G.; Kundu, K.; Stornaiuolo, M.; Scarpato, S.; Pollio, A.; Costantino, V. A Glimpse at Siderophores Production by Anabaena Flos-aquae UTEX 1444. Mar. Drugs 2022, 20, 256. [Google Scholar] [CrossRef]
- Goldman, S.J.; Lammers, P.J.; Berman, M.S.; Sanders-Loehr, J. Siderophore-Mediated Iron Uptake in Different Strains of Anabaena sp. J. Bacteriol. 1983, 156, 1144–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.; Butler, A. Structure of Synechobactins, New Siderophores of the Marine Cyanobacterium Synechococcus Sp. PCC 7002. Limnol. Oceanogr. 2005, 50, 1918–1923. [Google Scholar] [CrossRef]
- Guerinot, M.L.; Meidl, E.J.; Plessner, O. Citrate as a Siderophore in Bradyrhizobium Japonicum. J. Bacteriol. 1990, 172, 3298–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.; O’Brien, J.; Welch, T.; Clarke, P.; ÓCuív, P.; Crosa, J.H.; O’Connell, M. Genetic Organization of the Region Encoding Regulation, Biosynthesis, and Transport of Rhizobactin 1021, a Siderophore Produced by Sinorhizobium Meliloti. J. Bacteriol. 2001, 183, 2576–2585. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.S.; Moore, M.M. Ironing out Siderophore Biosynthesis: A Review of Non-Ribosomal Peptide Synthetase (NRPS)-Independent Siderophore Synthetases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 356–381. [Google Scholar] [CrossRef]
- Yang, J.; Banas, V.S.; Patel, K.D.; Rivera, G.S.M.; Mydy, L.S.; Gulick, A.M.; Wencewicz, T.A. An Acyl-Adenylate Mimic Reveals the Structural Basis for Substrate Recognition by the Iterative Siderophore Synthetase DesD. J. Biol. Chem. 2022, 298, 102166. [Google Scholar] [CrossRef] [PubMed]
- Raina, S.; Missiakas, D.; Baird, L.; Kumar, S.; Georgopoulos, C. Identification and Transcriptional Analysis of the Escherichia Coli HtrE Operon Which Is Homologous to Pap and Related Pilin Operons. J. Bacteriol. 1993, 175, 5009–5021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challis, G.L. A Widely Distributed Bacterial Pathway for Siderophore Biosynthesis Independent of Nonribosomal Peptide Synthetases. ChemBioChem 2005, 6, 601–611. [Google Scholar] [CrossRef] [PubMed]
- de Avila e Silva, S.; Echeverrigaray, S.; Gerhardt, G.J.L. BacPP: Bacterial Promoter Prediction—A Tool for Accurate Sigma-Factor Specific Assignment in Enterobacteria. J. Theor. Biol. 2011, 287, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Riediger, M.; Hernández-Prieto, M.A.; Song, K.; Hess, W.R.; Futschik, M.E. Genome-Wide Identification and Characterization of Fur-Binding Sites in the Cyanobacteria Synechocystis Sp. PCC 6803 and PCC 6714. DNA Res. 2021, 28, dsab023. [Google Scholar] [CrossRef]
- Winnikoff, J.R.; Glukhov, E.; Watrous, J.; Dorrestein, P.C.; Gerwick, W.H. Quantitative Molecular Networking to Profile Marine Cyanobacterial Metabolomes. J. Antibiot. 2014, 67, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Networking, G.-G.N.P.S.M. Feature Based Molecular Networking. Available online: Https://Gnps.Ucsd.Edu/ProteoSAFe/Status.Jsp?Task=53fbad6b425540ceb5eae088aac6ae32 (accessed on 17 October 2022).
- Soares, E.V. Perspective on the Biotechnological Production of Bacterial Siderophores and Their Use. Appl. Microbiol. Biotechnol. 2022, 106, 3985–4004. [Google Scholar] [CrossRef]
- Yu, S.; Teng, C.; Bai, X.; Liang, J.; Song, T.; Dong, L.; Jin, Y.; Qu, J. Optimization of Siderophore Production by Bacillus Sp. PZ-1 and Its Potential Enhancement of Phytoextration of Pb from Soi. J. Microbiol. Biotechnol. 2017, 27, 1500–1512. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Soliman, N.A.-M.; Abdullah, A.S.; El-Sharouny, E.E.; Matar, S.M.; Sabry, S.A.-F. Maximization of Siderophores Production from Biocontrol Agents, Pseudomonas Aeruginosa F2 and Pseudomonas Fluorescens JY3 Using Batch and Exponential Fed-Batch Fermentation. Processes 2020, 8, 455. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore Production by Bacillus Subtilis MF497446 and Pseudomonas Koreensis MG209738 and Their Efficacy in Controlling Cephalosporium Maydis in Maize Plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef]
- Sasirekha, B.; Srividya, S. Siderophore Production by Pseudomonas Aeruginosa FP6, a Biocontrol Strain for Rhizoctonia Solani and Colletotrichum Gloeosporioides Causing Diseases in Chilli. Agric. Nat. Resour. 2016, 50, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Murakami, C.; Tanaka, A.R.; Sato, Y.; Kimura, Y.; Morimoto, K. Easy Detection of Siderophore Production in Diluted Growth Media Using an Improved CAS Reagent. J. Microbiol. Methods 2021, 189, 106310. [Google Scholar] [CrossRef] [PubMed]
- Vindeirinho, J.M.; Soares, H.M.V.M.; Soares, E.V. Modulation of Siderophore Production by Pseudomonas Fluorescens Through the Manipulation of the Culture Medium Composition. Appl. Biochem. Biotechnol. 2021, 193, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Lemare, M.; Puja, H.; David, S.R.; Mathieu, S.; Ihiawakrim, D.; Geoffroy, V.A.; Rigouin, C. Engineering Siderophore Production in Pseudomonas to Improve Asbestos Weathering. Microb. Biotechnol. 2022, 15, 2351–2363. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Luria Broth (LB) Medium | |||
|---|---|---|---|
| Parameter | Range Tested | Optimal Value | Fold Induction at Optimal Value 1 |
| Temperature (°C) | 23–37 | 29 | 9.1 ± 0.1 * |
| pH | 6.0–8.0 | 7.5 | 2.7 ± 0.8 ** |
| NaCl g/L | 8.0–10.0 | 9.5 | 2.8 ± 0.4 ** |
| Fe3+ (μM) | 0–52 | 37 | 41.3 ± 0.9 ** |
| Fe2+ (μM) | 0–92 | 39.5 | 6.5 ± 0.1 ** |
| Citrate (mM) | 0–52 | 2.6 | 44.7 ± 0.1 ** |
| Glucose (g/L) | 0–10 | 2 | 25.9 ± 0.3 * |
| Lysine (mg/L) | 0–5 | 0.75 | 5.3 ± 0.4 ** |
| Minimal Medium | |||
| Parameter | Range tested | Optimal value | Fold Induction at optimal Value 1 |
| Temperature (°C) | 23–37 | 29 | 9.2 ± 0.8 * |
| pH | 6.0–8.0 | 7.5 | 3.3 ± 0.8 ** |
| NaCl g/L | 8.0–10.0 | 9.5 | 2.2 ± 0.2 * |
| Fe3+ (μM) | 0–52 | 30 | 20.0 ± 0.1 ** |
| Fe2+ (μM) | 0–92 | 92 | 20.7 ± 0.1 ** |
| Citrate (mM) | 0–52 | 2 | 8.3 ± 0.1 ** |
| Glucose (g/L) | 0–10 | 10 | 43.6 ± 0.1 ** |
| Lysine (mg/L) | 0–5 | 0.75 | 3.1 ± 0.1 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundu, K.; Teta, R.; Esposito, G.; Stornaiuolo, M.; Costantino, V. A Four-Step Platform to Optimize Growth Conditions for High-Yield Production of Siderophores in Cyanobacteria. Metabolites 2023, 13, 154. https://doi.org/10.3390/metabo13020154
Kundu K, Teta R, Esposito G, Stornaiuolo M, Costantino V. A Four-Step Platform to Optimize Growth Conditions for High-Yield Production of Siderophores in Cyanobacteria. Metabolites. 2023; 13(2):154. https://doi.org/10.3390/metabo13020154
Chicago/Turabian StyleKundu, Karishma, Roberta Teta, Germana Esposito, Mariano Stornaiuolo, and Valeria Costantino. 2023. "A Four-Step Platform to Optimize Growth Conditions for High-Yield Production of Siderophores in Cyanobacteria" Metabolites 13, no. 2: 154. https://doi.org/10.3390/metabo13020154
APA StyleKundu, K., Teta, R., Esposito, G., Stornaiuolo, M., & Costantino, V. (2023). A Four-Step Platform to Optimize Growth Conditions for High-Yield Production of Siderophores in Cyanobacteria. Metabolites, 13(2), 154. https://doi.org/10.3390/metabo13020154