


Chemical Composition, Biomolecular Analysis, and Nuclear Magnetic Resonance Spectroscopic Fingerprinting of Posidonia oceanica and Ascophyllum nodosum Extracts

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Harvesting and Preparation
2.3. Chemical Characterization
2.4. Extraction
2.5. Carbohydrate Composition
2.6. Aminoacidic Composition
2.7. Total Phenolic Content
2.8. NMR Analysis
2.9. Statistical Analysis
3. Results and Discussions
3.1. Chemical Features
3.2. Bio(macro)molecules
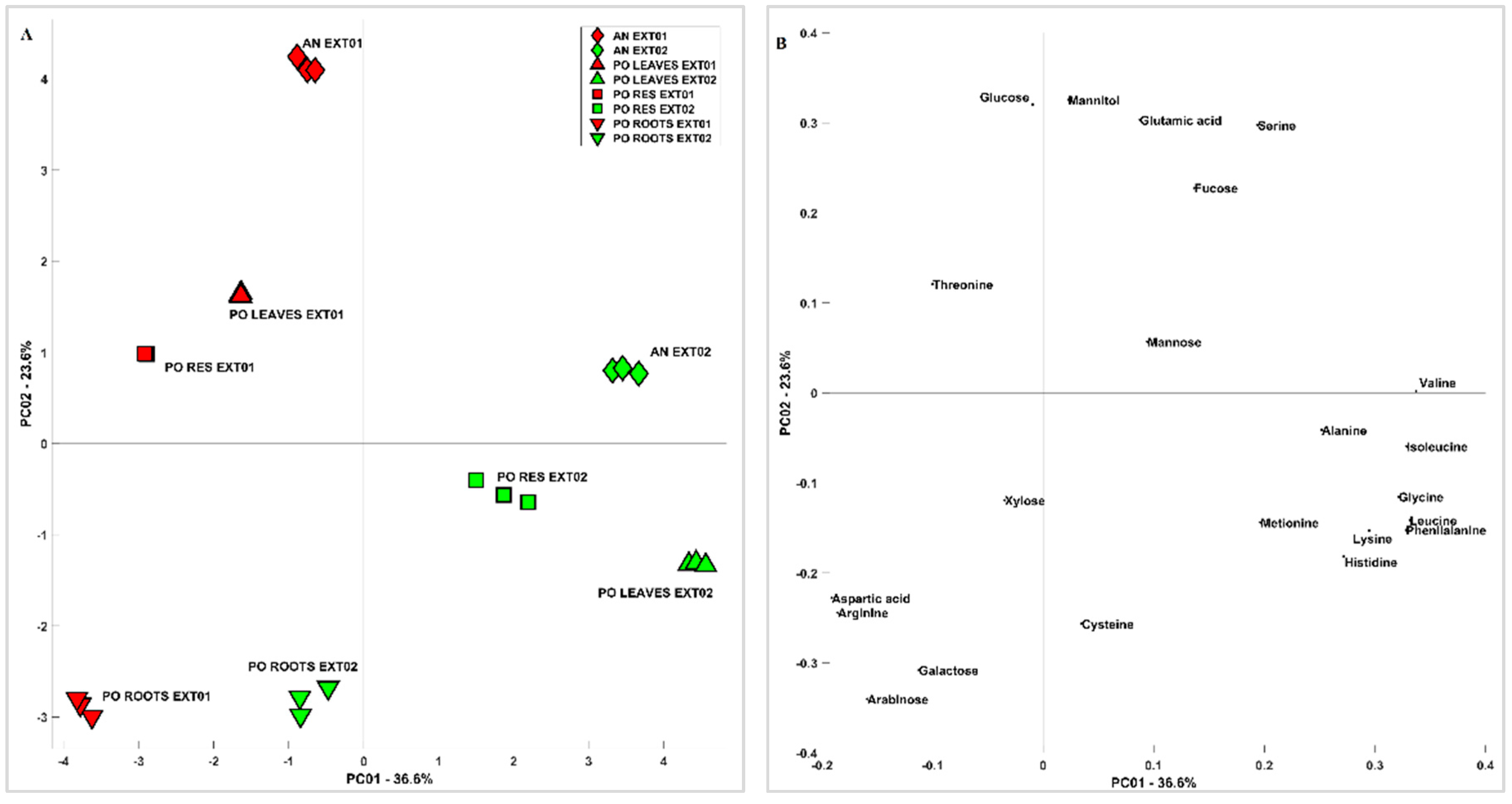
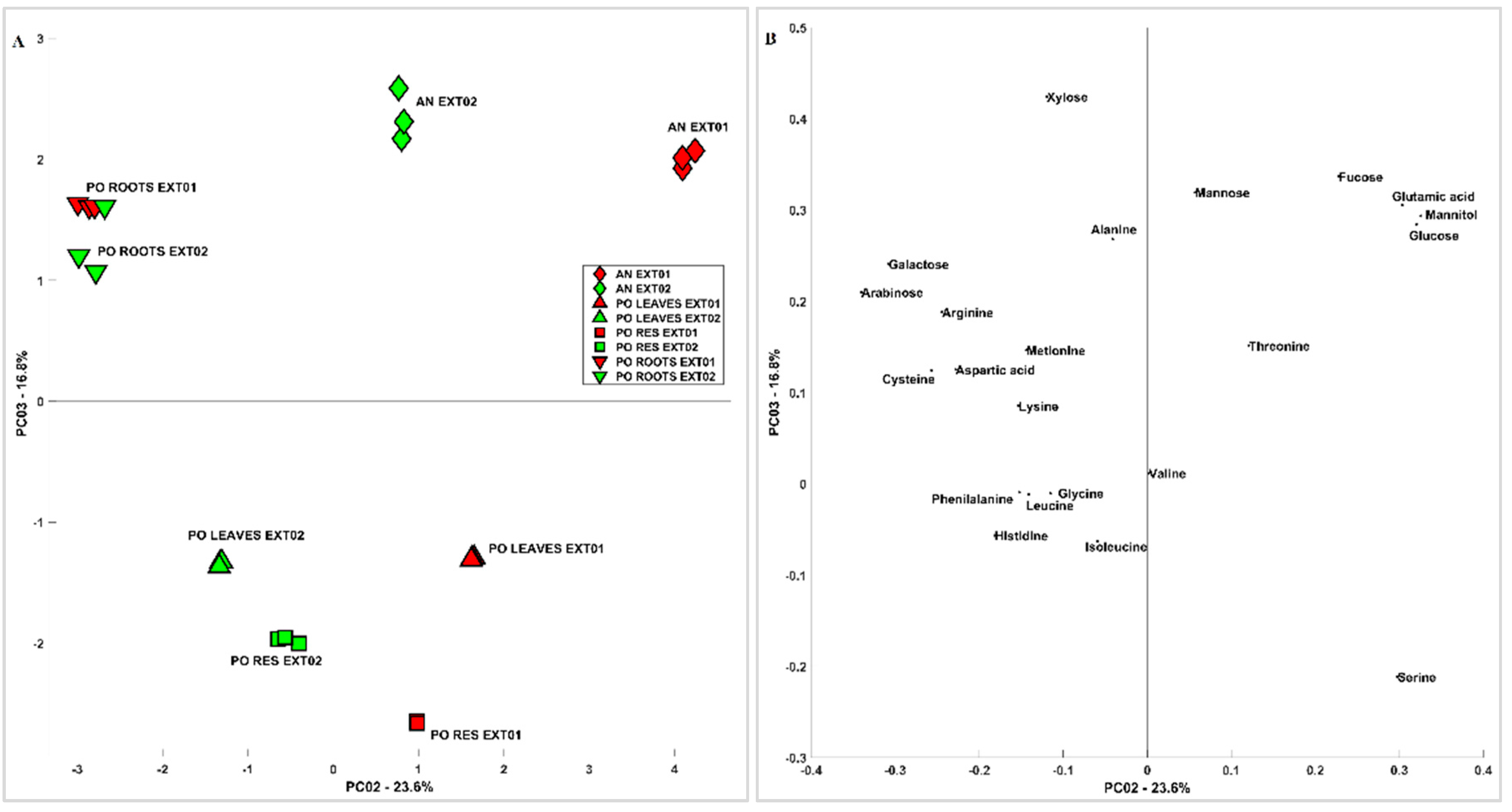
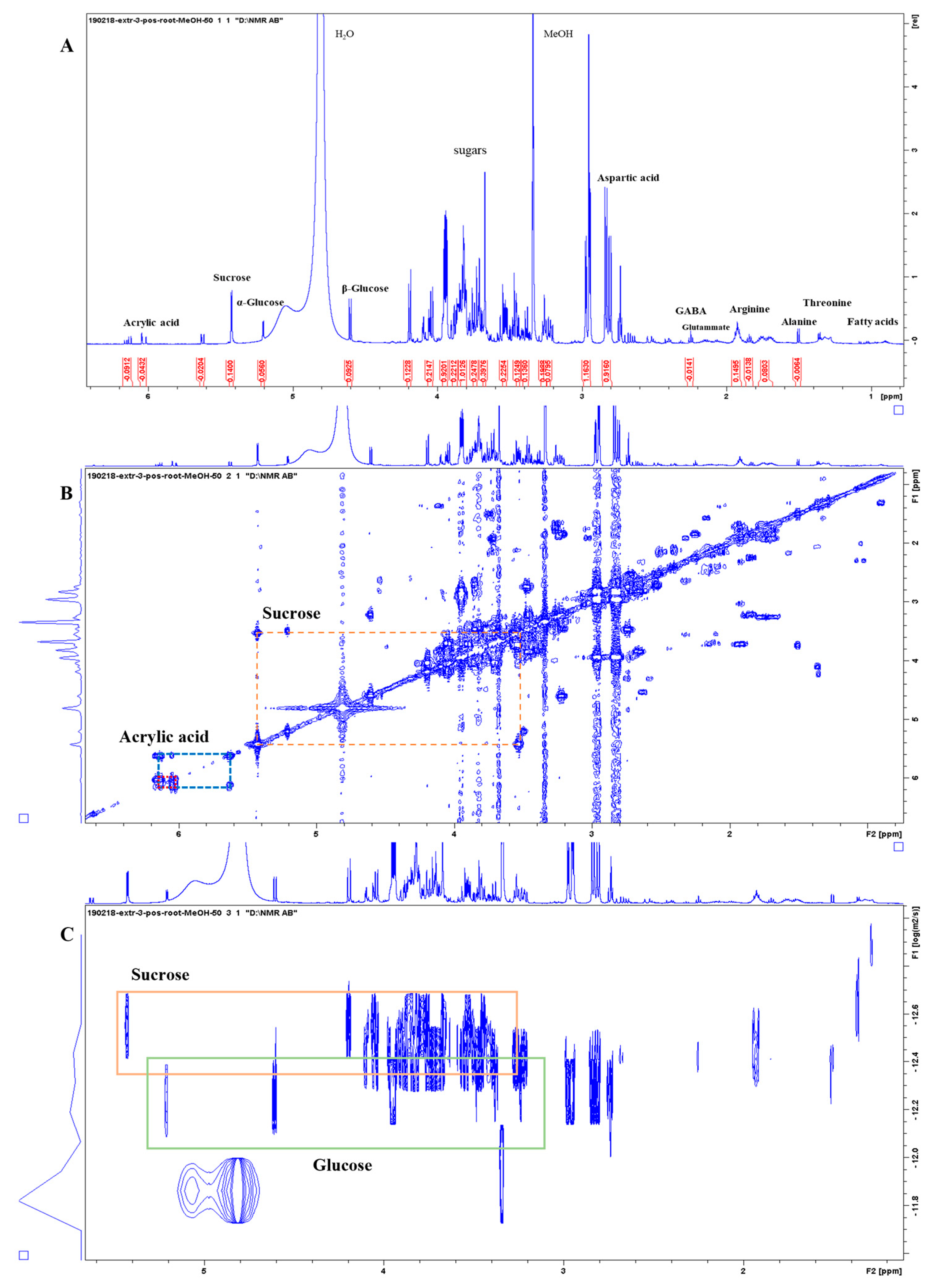
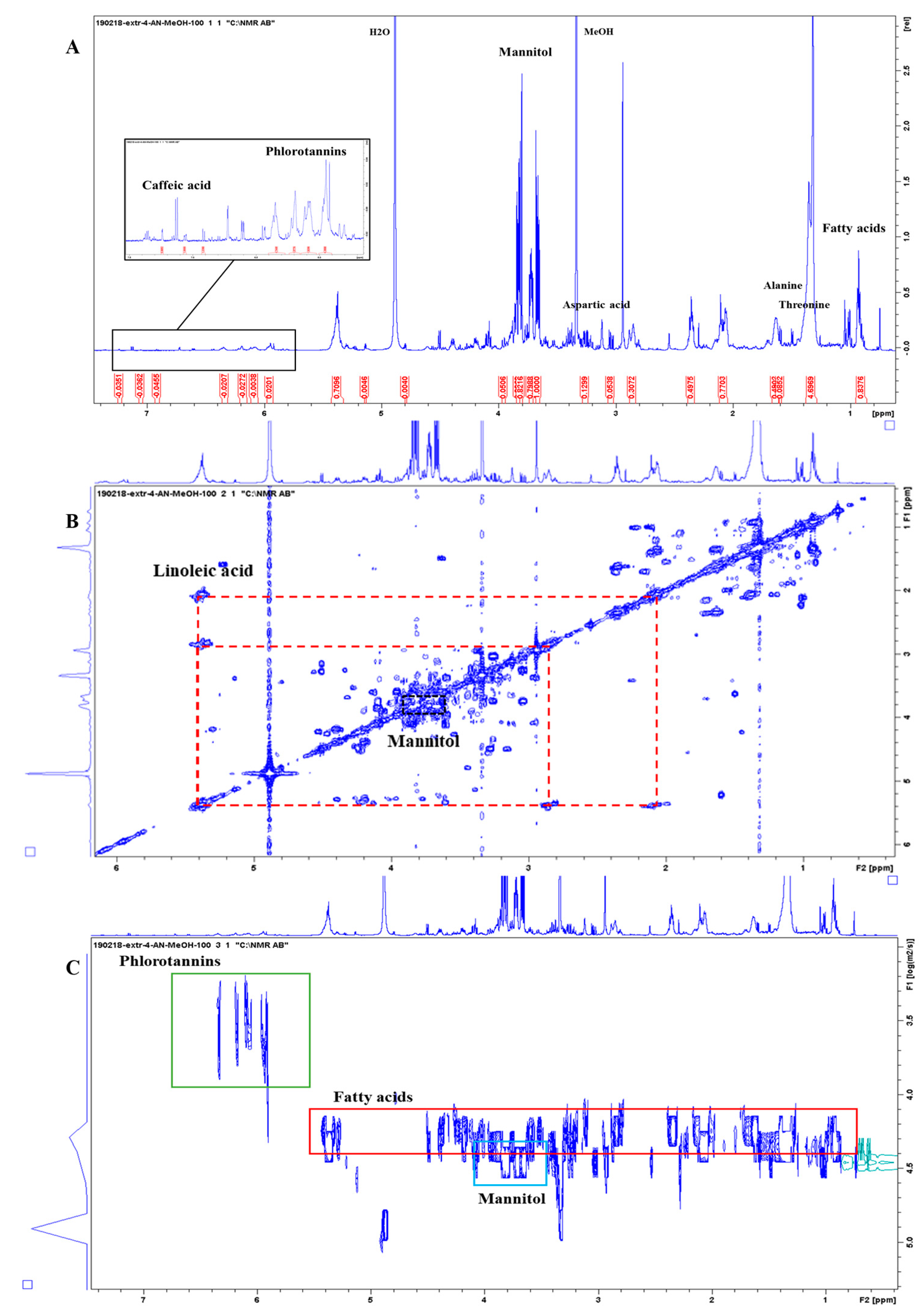
3.3. 1D and 2D 1H-NMR Analyses of PO and AN
3.3.1. Molecular Characterization of Posidonia oceanica and Ascophyllum nodosum
3.3.2. Metabolite Identification of Posidonia oceanica and Ascophyllum nodosum
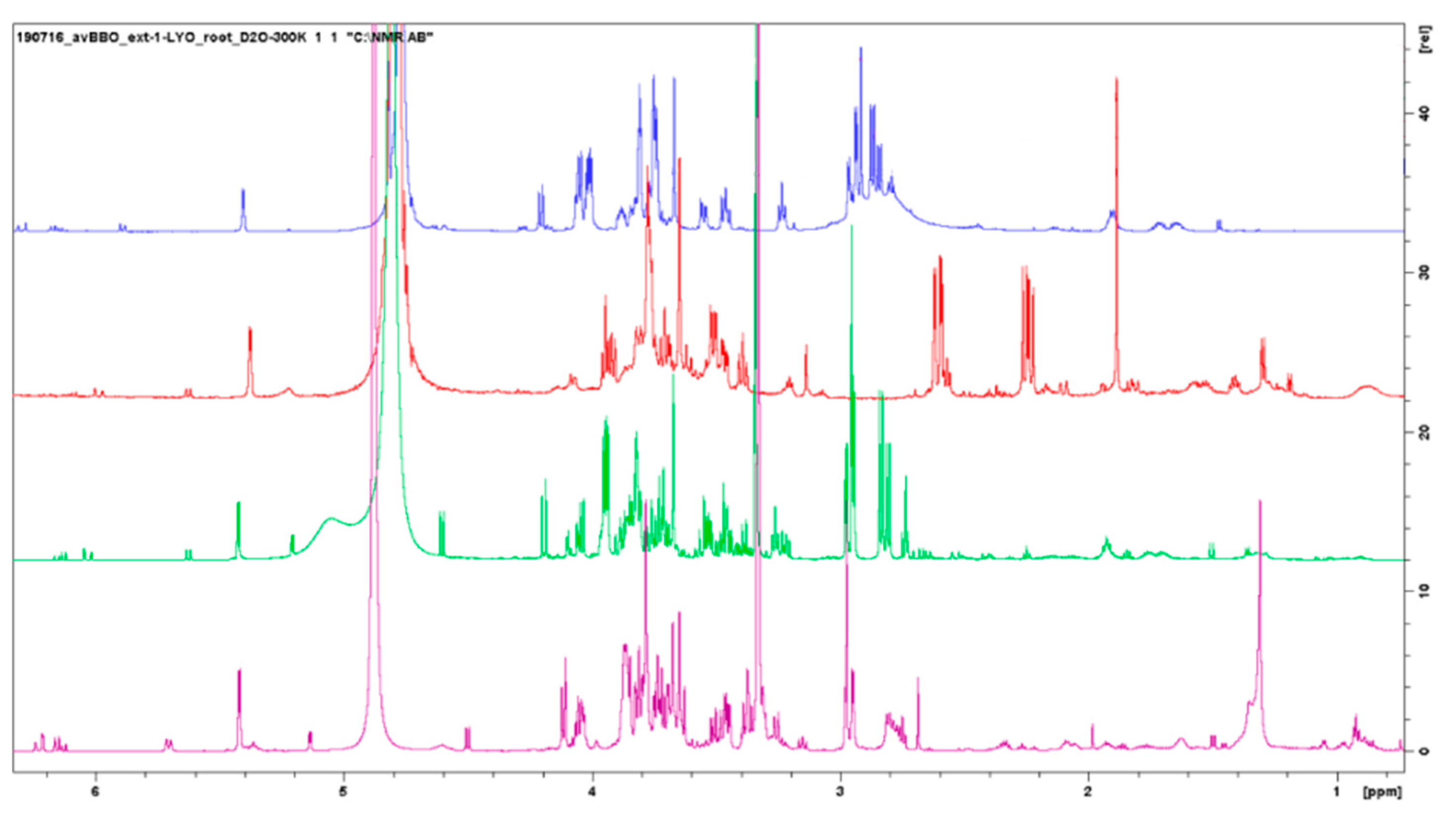
3.3.3. Comparison between Posidonia oceanica Roots, Leaves, and Residues
3.3.4. Evaluation of Extraction Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duarte, C.M. How can beaches be managed with respect to seagrass litter. In European Seagrasses: An Introduction to Monitoring and Management; M&MS Project: Brussels, Belgium, 2004; pp. 83–84. [Google Scholar]
- Cebrian, J.; Duarte, C.M. Detrital stocks and dynamics of the seagrass Posidonia oceanica (L.) Delile in the Spanish Mediterranean. Aquat. Bot. 2001, 70, 295–309. [Google Scholar]
- Jeudy de Grissac, A. Effets des herbiers à Posidonia oceanica sur la dynamique marine et la sédimentologie littorale. In Proceedings of the International Workshop on Posidonia oceanica Beds, Porquerolles, France, 12–15 October 1983; Volume 1, pp. 437–443. [Google Scholar]
- Cocozza, C.; Parente, A.; Zaccone, C.; Mininni, C.; Santamaria, P.; Miano, T. Chemical, physical and spectroscopic characterization of Posidonia oceanica (L.) Del. residues and their possible recycle. Biomass Bioenergy 2011, 35, 799–807. [Google Scholar] [CrossRef]
- Benito-González, I.; López-Rubio, A.; Martínez-Sanz, M. Potential of lignocellulosic fractions from Posidonia oceanica to improve barrier and mechanical properties of bio-based packaging materials. Int. J. Biol. Macromol. 2018, 118, 542–551. [Google Scholar] [PubMed]
- Hamdaoui, O.; Ibos, L.; Mazioud, A.; Safi, M.; Limam, O. Thermophysical characterization of Posidonia oceanica marine fibers intended to be used as an insulation material in Mediterranean buildings. Constr. Build. Mater. 2018, 180, 68–76. [Google Scholar]
- Vasarri, M.; Leri, M.; Barletta, E.; Ramazzotti, M.; Marzocchini, R.; Degl’Innocenti, D. Anti-inflammatory properties of the marine plant Posidonia oceanica (L.) Delile. J. Ethnopharmacol. 2020, 247, 112252. [Google Scholar] [PubMed]
- Leri, M.; Ramazzotti, M.; Vasarri, M.; Peri, S.; Barletta, E.; Pretti, C.; Degl’Innocenti, D. Bioactive compounds from Posidonia oceanica (L.) delile impair malignant cell migration through autophagy modulation. Mar. Drugs 2018, 16, 137. [Google Scholar] [PubMed] [Green Version]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Posidonia oceanica (L.) delile ethanolic extract modulates cell activities with skin health applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [PubMed] [Green Version]
- Salvi, L.; Brunetti, C.; Cataldo, E.; Niccolai, A.; Centritto, M.; Ferrini, F.; Mattii, G.B. Effects of Ascophyllum nodosum extract on Vitis vinifera: Consequences on plant physiology, grape quality and secondary metabolism. Plant Physiol. Biochem. 2019, 139, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Allwood, J.W.; De Vos, R.C.; Moing, A.; Deborde, C.; Erban, A.; Kopka, J.; Goodacre, R.; Hall, R.D. Plant metabolomics and its potential for systems biology research: Background concepts, technology, and methodology. Meth. Enzymol. 2011, 500, 299–336. [Google Scholar]
- Eisenreich, W.; Bacher, A. Advances of high-resolution NMR techniques in the structural and metabolic analysis of plant biochemistry. Phytochemistry 2007, 68, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.V.; Guerra, J.; Velders, A.H.; Crooks, R.M. NMR characterization of fourth generation PAMAM dendrimers in the presence and absence of palladium dendrimer-encapsulated nanoparticles. J. Am. Chem. Soc. 2009, 131, 341–350. [Google Scholar] [PubMed]
- Barjat, H.; Morris, G.A.; Smart, S.; Swanson, A.G.; Williams, S.C. High-resolution diffusion-ordered 2D spectroscopy (HR-DOSY)-a new tool for the analysis of complex mixtures. J. Magn. Reason. Ser. B 1995, 108, 170–172. [Google Scholar] [CrossRef]
- Kjeldahl, J. A new method for the estimation of nitrogen in organic compounds. Z. Anal. Chem. 1883, 22, 366. [Google Scholar] [CrossRef] [Green Version]
- Springer, U.; Klee, J. Prüfung der Leistungsfähigkeit von einigen wichtigeren Verfahren zur Bestimmung des Kohlenstoffs mittels Chromschwefelsäure sowie Vorschlag einer neuen Schnellmethode. J. Plant Nutr. Soil Sci. 1954, 64, 1–26. [Google Scholar] [CrossRef]
- Briceño-Domínguez, D.; Hernández-Carmona, G.; Moyo, M.; Stirk, W.; van Staden, J. Plant growth promoting activity of seaweed liquid extracts produced from Macrocystis pyrifera under different pH and temperature conditions. J. Appl. Phycol. 2014, 26, 2203–2210. [Google Scholar]
- Cui, H.Y.; Niranjana Murthy, H.; Moh, S.H.; Cui, Y.Y.; Lee, E.J.; Paek, K.Y. Comparison of conventional and ultrasound-assisted methods for extraction of nutraceutical compounds from Dendrobium candidum. CYTA J. Food 2014, 12, 355–359. [Google Scholar]
- Zhang, C.; Sanders, J.P.; Bruins, M.E. Critical parameters in cost-effective alkaline extraction for high protein yield from leaves. Biomass Bioenergy 2014, 67, 466–472. [Google Scholar] [CrossRef]
- Manns, D.; Deutschle, A.L.; Saake, B.; Meyer, A.S. Methodology for quantitative determination of the carbohydrate composition of brown seaweeds (Laminariaceae). RSC Adv. 2014, 4, 25736–25746. [Google Scholar] [CrossRef] [Green Version]
- Cataldi, T.R.; Angelotti, M.; Bianco, G. Determination of mono-and disaccharides in milk and milk products by high-performance anion-exchange chromatography with pulsed amperometric detection. Anal. Chim. Acta 2003, 485, 43–49. [Google Scholar] [CrossRef]
- Tsugita, A.; Scheffler, J.J. A rapid method for acid hydrolysis of protein with a mixture of trifluoroacetic acid and hydrochloric acid. Eur. J. Biochem. 1982, 124, 585–588. [Google Scholar] [PubMed]
- Gómez-Alonso, S.; Hermosín-Gutiérrez, I.; García-Romero, E. Simultaneous HPLC analysis of biogenic amines, amino acids, and ammonium ion as aminoenone derivatives in wine and beer samples. J. Agric. Food Chem. 2007, 55, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Cicco, N.; Lanorte, M.T.; Paraggio, M.; Viggiano, M.; Lattanzio, V. A reproducible, rapid, and inexpensive Folin–Ciocalteu micro-method in determining phenolics of plant methanol extracts. Microchem. J. 2009, 91, 107–110. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Jolliffe, I.T. Principal components in regression analysis. In Principal Component Analysis; Springer: New York, NY, USA, 2002; pp. 167–198. [Google Scholar]
- Castillo, C.; Mantecón, A.R.; Sotillo, J.; Benedito, J.L.; Abuelo, A.; Gutiérrez, C.; Hernández, J. The use of banquettes of Posidonia oceanica as a source of fiber and minerals in ruminant nutrition. An observational study. Animal 2014, 8, 1663–1666. [Google Scholar]
- Sanz-Lázaro, C.; Malea, P.; Apostolaki, E.T.; Kalantzi, I.; Marín, A.; Karakassis, I. The role of the seagrass Posidonia oceanica in the cycling of trace elements. Biogeosciences 2012, 9, 2497–2507. [Google Scholar] [CrossRef] [Green Version]
- Tranchina, L.; Brai, M.; D’Agostino, F.; Bartolotta, A.; Rizzo, G. Trace metals in “Posidonia oceanica” seagrass from south-eastern Sicily. Chem. Ecol. 2005, 21, 109–118. [Google Scholar] [CrossRef]
- Ben Salem, Y.; Abdelhamid, A.; Mkadmini Hammi, K.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Microwave-assisted extraction and pharmacological evaluation of polysaccharides from Posidonia oceanica. Biosci. Biotechnol. Biochem. 2017, 81, 1917–1925. [Google Scholar]
- Benito-González, I.; López-Rubio, A.; Martínez-Abad, A.; Ballester, A.R.; Falcó, I.; González-Candelas, L.; Segura-Carretero, A.; Martínez-Sanz, M. In-depth characterization of bioactive extracts from Posidonia oceanica waste biomass. Mar. Drugs 2019, 17, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaal, J.; Serrano, O.; Nierop, K.G.; Schellekens, J.; Cortizas, A.M.; Mateo, M.Á. Molecular composition of plant parts and sediment organic matter in a Mediterranean seagrass (Posidonia oceanica) mat. Aquat. Bot. 2016, 133, 50–61. [Google Scholar]
- Torbatinejad, N.M.; Annison, G.; Rutherfurd-Markwick, K.; Sabine, J.R. Structural constituents of the seagrass Posidonia australis. J. Agric. Food Chem. 2007, 55, 4021–4026. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.; Diop, A.; Rainville, L.C.; Barnabé, S. Bioextracting Polyphenols from the Brown Seaweed Ascophyllum nodosum from Québec’s North Shore Coastline. Ind. Biotechnol. 2019, 15, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Ragan, M.A.; Jensen, A. Quantitative studies on brown algal phenols. II. Seasonal variation in polyphenol content of Ascophyllum nodosum (L.) Le Jol. and Fucus vesiculosus (L.). J. Exp. Mar. Biol. Ecol. 1978, 34, 245–258. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Reed, R.H.; Davison, I.R.; Chudek, J.A.; Foster, R. The osmotic role of mannitol in the Phaeophyta: An appraisal. Phycologia 1985, 24, 35–47. [Google Scholar] [CrossRef]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef]
- Shibata, T.; Nagayama, K.; Tanaka, R.; Yamaguchi, K.; Nakamura, T. Inhibitory effects of brown algal phlorotannins on secretory phospholipase A2s, lipoxygenases and cyclooxygenases. J. Appl. Phycol. 2003, 15, 61–66. [Google Scholar]
- Van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar]
- Rayirath, P.; Benkel, B.; Hodges, D.M.; Allan-Wojtas, P.; MacKinnon, S.; Critchley, A.T.; Prithiviraj, B. Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana. Planta 2009, 230, 135–147. [Google Scholar]
- Sobolev, A.P.; Brosio, E.; Gianferri, R.; Segre, A.L. Metabolic profile of lettuce leaves by high-field NMR spectra. Magn. Reason. Chem. 2005, 43, 625–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannina, L.; Cristinzio, M.; Sobolev, A.P.; Ragni, P.; Segre, A. High-field nuclear magnetic resonance (NMR) study of truffles (Tuber aestivum vittadini). J. Agric. and Food Chem. 2004, 52, 7988–7996. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, A.P.; Segre, A.; Lamanna, R. Proton high-field NMR study of tomato juice. Magn. Reason. Chem. 2003, 41, 237–245. [Google Scholar]
- BMRB. Biological Magnetic Resonance Bank. Available online: http://www.bmrb.wisc.edu/ (accessed on 30 March 2020).
- Human Metabolome Database. Available online: http://www.hmdb.ca/ (accessed on 30 March 2020).
- Grignon-Dubois, M.; Rezzonico, B. Phenolic fingerprint of the seagrass Posidonia oceanica from four locations in the Mediterranean Sea: First evidence for the large predominance of chicoric acid. Bot. Mar. 2015, 58, 379–391. [Google Scholar] [CrossRef]
- Haznedaroglu, M.Z.; Zeybek, U. HPLC Determination of Chicoric Acid in Leaves of Posidonia oceanica. Pharm. Biol. 2007, 45, 745–748. [Google Scholar] [CrossRef]
- Lee, J.; Scagel, C.F. Chicoric acid: Chemistry, distribution, and production. Front. Chem. 2013, 1, 40. [Google Scholar]
- King, P.J.; Robinson, W.E. Resistance to the anti-human immunodeficiency virus type 1 compound L-chicoric acid results from a single mutation at amino acid 140 of integrase. J. Virol. 1998, 72, 8420–8424. [Google Scholar] [CrossRef] [Green Version]
- Viso, A.C.; Pesando, D.; Bernard, P.; Marty, J.C. Lipid components of the Mediterranean seagrass Posidonia oceanica. Phytochemistry 1993, 34, 381–387. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extractions | |||
|---|---|---|---|
| No. 1 (Acid Extraction) | No. 2 (Alkaline Extraction) | No. 3 (Hydroalcoholic Extraction) | No. 4 (Alcoholic Extraction) |
| Citric acid (0.6%), ascorbic acid (0.3%) | NaOH 1.5 M | Methanol 50% | Methanol 100% |
| 40 °C × 4 h (shaker water bath) | 80 °C × 4 h (shaker water bath) | 40 °C × 1.5 h (shaker water bath) | 40 °C × 1.5 h (shaker water bath) |
| Filtration | Filtration | 30 min sonication | 30 min sonication |
| Centrifugation: 5000 rpm × 10 min | Centrifugation: 5000 rpm × 10 min | 30 min, 40 °C (shaker water bath) | 30 min, 40 °C (shaker water bath) |
| Filtration with syringe filter 0.45 µm | Filtration with syringe filter 0.45 µm | ||
| Chemical Features (Mean ± Standard Deviation) | |||||
|---|---|---|---|---|---|
| Posidonia oceanica | Ascophyllum nodosum | ||||
| Residues | Fresh | ||||
| ROOTS | LEAVES | ||||
| pHH2O | 7.9 ± 0.1 | 7.9 ± 0.2 | 7.7 ± 0.1 | 6.3 ± 0.1 | |
| Ash | (%) | 28.2 ± 0.3 | 17.9 ± 0.2 | 25.8 ± 0.1 | 18 ± 0.5 |
| Ntot | (g kg-1 d.m.) | 5.7 ± 0.2 | 14.8 ± 0.7 | 14.8 ± 0.02 | 12.3 ± 0.4 |
| N-NH4 | (g kg-1 d.m.) | 0.12 ± 0.02 | 0.1 ± 0.01 | 0.08 ± 0.01 | 0.3 ± 0.01 |
| Norg | (g kg-1 d.m.) | 5.58 ± 0.2 | 14.7 ± 0.7 | 14.72 ± 0.02 | 12 ± 0.4 |
| Corg | (g kg-1 d.m.) | 359 ± 9 | 410.4 ± 14 | 378 ± 8 | 409.6 ± 12 |
| C/N | 63 | 27.7 | 25 | 32.5 | |
| Ptot | (g kg-1 d.m.) | 0.3 ± 0.01 | 0.34 ± 0.01 | 0.6 ± 0.02 | 0.61 ± 0.01 |
| Catot | (g kg-1 d.m.) | 33.4 ± 0.6 | 15.9 ± 0.2 | 45.8 ± 1 | 8 ± 0.05 |
| Ktot | (g kg-1 d.m.) | 2.1 ± 0.01 | 4.7 ± 0.02 | 1.6 ± 0.01 | 11.4 ± 0.03 |
| Mgtot | (g kg-1 d.m.) | 7 ± 0.08 | 5 ± 0.05 | 10 ± 0.1 | 6 ± 0.01 |
| Natot | (g kg-1 d.m.) | 20.7 ± 0.2 | 15.2 ± 0.2 | 4 ± 0.1 | 17 ± 0.1 |
| Btot | (mg kg-1 d.m.) | 1613.4 ± 24 | 852.3 ± 17 | 1427.9 ± 22 | 95 ± 1.6 |
| Fetot | (mg kg-1 d.m.) | 4758 ± 68 | 2438.4 ± 36 | 489.9 ± 9 | 372.8 ± 8 |
| Mntot | (mg kg-1 d.m.) | 130.1 ± 1.2 | 108.9 ± 1 | 369.3 ± 3.1 | 48.2 ± 0.6 |
| Motot | (mg kg-1 d.m.) | 36.2 ± 0.8 | 7 ± 0.2 | 0.2 ± 0.01 | 1.3 ± 0.01 |
| Zntot | (mg kg-1 d.m.) | 37.3 ± 0.1 | 51.6 ± 0.1 | 113 ± 0.2 | 100.2 ± 0.22 |
| Cutot | (mg kg-1 d.m.) | 28 ± 0.1 | 33 ± 0.1 | 22.2 ± 0.1 | 10 ± 0.05 |
| Posidonia oceanica | Ascophyllum nodosum | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Residues | Fresh Plant | |||||||||
| ROOTS | LEAVES | |||||||||
| Carbohydrates (%) | ||||||||||
| EXT 1 | EXT 2 | EXT 1 | EXT 2 | EXT 1 | EXT 2 | EXT 1 | EXT 2 | |||
| components | rt | |||||||||
| 1 | Mannitol | 2.45 | 0 | 0 | 0 | 0 | 0 | 0 | 7.05 ± 0.25 | 2.51 ± 0.15 |
| 2 | Fucose | 4.27 | 0.06 ± 0.00 | 0.46 ± 0.02 | 0.05 ± 0.00 | 0.20 ± 0.01 | 0 | 0 | 2.00 ± 0.03 | 2.94 ± 0.12 |
| 3 | Arabinose | 8.6 | 0 | 0.17 ± 0.01 | 1.38 ± 0.01 | 1.14 ± 0.05 | 0.02 ± 0.01 | 0.08 ± 0.01 | 0 | 0 |
| 4 | Galactose | 10.5 | 0.16 ± 0.00 | 0.26 ± 0.01 | 1.52 ± 0.03 | 2.39 ± 0.39 | 0.12 ± 0.00 | 0.20 ± 0.00 | 0.24 ± 0.01 | 0.38 ± 0.01 |
| 5 | Glucose | 12.2 | 0.10 ± 0.00 | 0.12 ± 0.00 | 0.26 ± 0.01 | 1.23 ± 0.14 | 1.84 ± 0.04 | 0.44 ± 0.02 | 3.98 ± 0.04 | 1.54 ± 0.06 |
| 6 | Xylose | 14.4 | 0.24 ± 0.01 | 0.1± 0.00 | 1.65 ± 0.01 | 0.80 ± 0.14 | 0.03 ± 0.00 | 0.05 ± 0.02 | 0.57 ± 0.01 | 1.72 ± 0.07 |
| 7 | Mannose | 15.3 | 0.11 ± 0.00 | 0.09± 0.00 | 0.06 ± 0.01 | 0.24 ± 0.11 | 0 | 0 | 0.18 ± 0.02 | 0.47 ± 0.04 |
| Tot | 0.68 ± 0.02 | 1.21 ± 0.04 | 4.93 ± 0.04 | 6.01 ± 0.19 | 2.01 ± 0.04 | 0.77 ± 0.06 | 14.02 ± 0.35 | 9.55 ± 0.44 | ||
| Amino acids (%) | ||||||||||
| components | rt | |||||||||
| 1 | Aspartic acid | 2.78 | 0.31 ± 0.00 | 0.51 ± 0.03 | 13.94 ± 0.10 | 4.36 ± 0.18 | 7.25 ± 0.03 | 2.44 ± 0.02 | 0.42 ± 0.01 | 0.73 ± 0.00 |
| 2 | Glutamic acid | 3.78 | 0.27 ± 0.00 | 0.48 ± 0.03 | 0.76 ± 0.01 | 0.51 ± 0.02 | 1.27 ± 0.01 | 1.05 ± 0.01 | 2.28 ± 0.06 | 1.65 ± 0.03 |
| 3 | Serine | 8.79 | 0.18 ± 0.00 | 0.23 ± 0.01 | 0 | 0 | 0.27 ± 0.00 | 0.27 ± 0.01 | 0.23 ± 0.00 | 0.23 ± 0.00 |
| 4 | Histidine | 12.3 | 0.21 ± 0.00 | 0.29 ± 0.01 | 0.20 ± 0.02 | 0.29 ± 0.01 | 0.21 ± 0.00 | 0.30 ± 0.01 | 0.22 ± 0.01 | 0.26 ± 0.01 |
| 5 | Glycine | 12.65 | 0 | 0.39 ± 0.03 | 0 | 0.35 ± 0.01 | 0.12 ± 0.01 | 0.51 ± 0.02 | 0.14 ± 0.00 | 0.35 ± 0.00 |
| 6 | Threonine | 13.87 | 0 | 0 | 0 | 0.23 ± 0.01 | 0.20 ± 0.00 | 0 | 0.20 ± 0.00 | 0 |
| 7 | Arginine | 15.69 | 0 | 0 | 3.67 ± 0.08 | 0.47 ± 0.01 | 0 | 0 | 0 | 0 |
| 8 | Alanine | 16.66 | 0 | 0.37 ± 0.02 | 0.39 ± 0.00 | 0.34 ± 0.01 | 0.31 ± 0.00 | 0.59 ± 0.02 | 0.44 ± 0.01 | 0.54 ± 0.01 |
| 9 | Tyrosine | 19.52 | nd | nd | nd | nd | nd | nd | nd | nd |
| 10 | Valine | 21.55 | 0 | 0.32 ± 0.01 | 0 | 0.24 ± 0.01 | 0.23 ± 0.00 | 0.48 ± 0.00 | 0.20 ± 0.00 | 0.39 ± 0.01 |
| 11 | Metionine | 22.1 | 0 | 0 | 0.05 ± 0.00 | 0 | 0 | 0.08 ± 0.00 | 0 | 0.07 ± 0.00 |
| 12 | Isoleucine | 24.31 | 0 | 0.24 ± 0.01 | 0 | 0 | 0 | 0.32 ± 0.01 | 0 | 0.28 ± 0.01 |
| 13 | Leucine | 24.87 | 0 | 0.37 ± 0.02 | 0 | 0.26 ± 0.02 | 0 | 0.65 ± 0.01 | 0 | 0.50 ± 0.00 |
| 14 | Phenilalanine | 25.31 | 0 | 0.30 ± 0.01 | 0 | 0.22 ± 0.00 | 0 | 0.45 ± 0.01 | 0 | 0.36 ± 0.00 |
| 15 | Cysteine | 26.9 | 0 | 0.19 ± 0.01 | 0.09 ± 0.01 | 1.39 ± 0.04 | 0 | 0.31 ± 0.00 | 0 | 0.17 ± 0.01 |
| 16 | Lysine | 28.57 | 0 | 0.41 ± 0.03 | 0.29 ± 0.00 | 0.33 ± 0.00 | 0.25 ± 0.00 | 0.63 ± 0.00 | 0.24 ± 0.01 | 0.44 ± 0.00 |
| 17 | Proline | nd | nd | nd | nd | nd | nd | nd | nd | |
| Tot | 0.97 ± 0.08 | 4.10 ± 0.25 | 19.39 ± 0.19 | 9.00 ± 0.29 | 10.11 ± 0.08 | 8.08 ± 0.14 | 4.36 ± 0.22 | 5.97 ± 0.10 | ||
| Total phenols (mgGAE/g DW) | ||||||||||
| EXT 3 | EXT 4 | EXT 3 | EXT 4 | EXT 3 | EXT 4 | EXT 3 | EXT 4 | |||
| 0.49 ± 0.01 | 0.72 ± 0.01 | 46.9 ± 0.19 | 40.8 ± 0.55 | 48.2 ± 0.67 | 29.7 ± 0.18 | 115 ± 0.8 | 15.6 ± 0.18 | |||
| Posidonia oceanica | ||||
|---|---|---|---|---|
| Compounds | Assignment | δ 1H (ppm) | Multiplicity | Connectivity |
| Carbohydrates | ||||
| α-D-glucose (α-Gluc) | CH-4 | 3.39 | m | |
| CH-2 | 3.49 | dd | ||
| CH2-6 | 3.73 | m | ||
| CH-3 | 3.74 | m | ||
| CH-5, CH2-6 | 3.83 | m | ||
| CH-1 | 5.21 | d | 1–2 | |
| β-D-glucose (β-Gluc) | CH-2 | 3.22 | dd | 2–3 |
| CH-4 | 3.39 | m | 4–5 | |
| CH-3,5 | 3.46 | m | 3–4, 5–6 | |
| CH2-6 | 3.76 | m | ||
| CH2-6 | 3.85 | dd | ||
| CH-1 | 4.61 | d | 1–2 | |
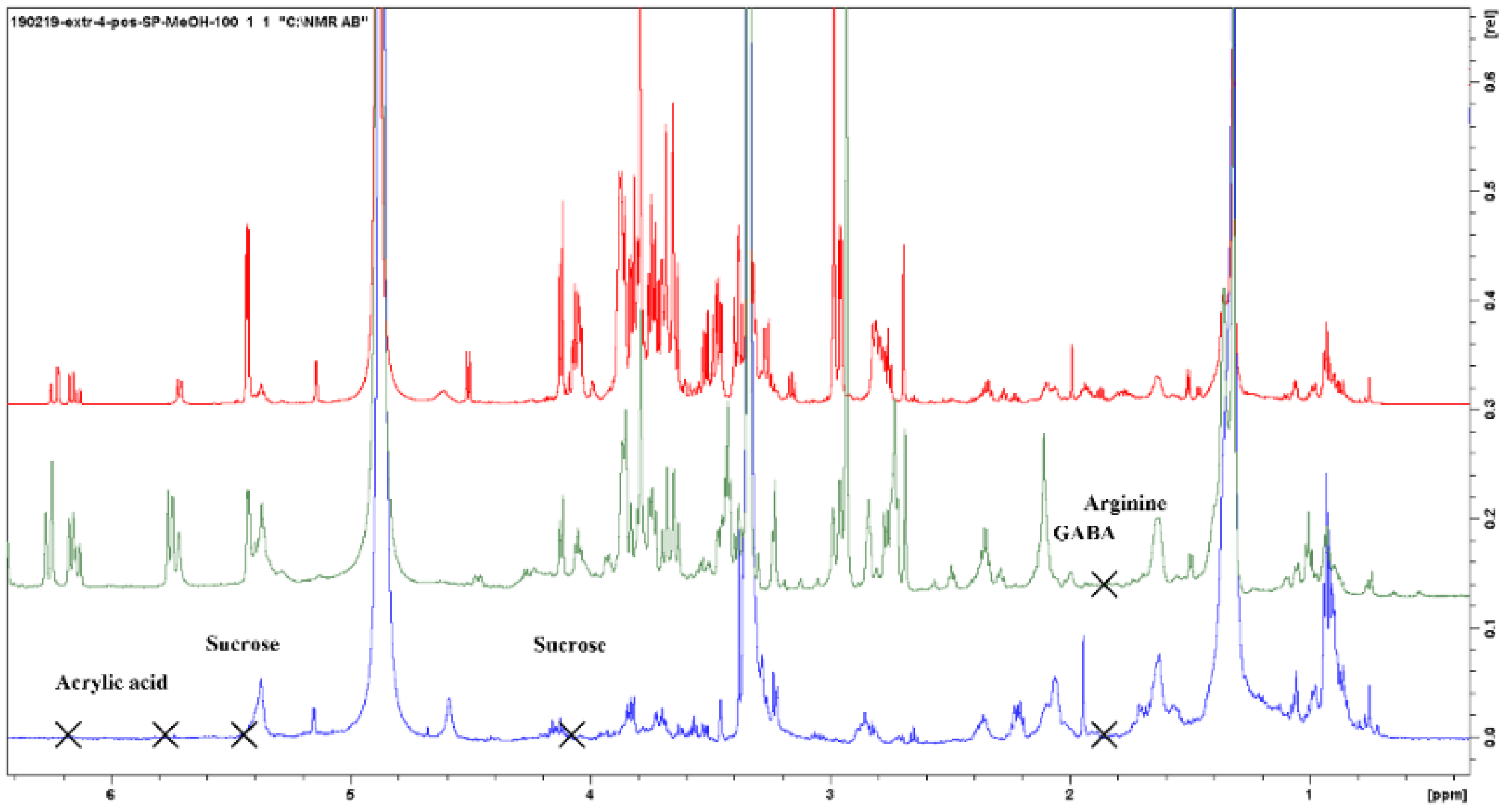
| Sucrose (Suc) | CH-4 | 3.46 | t | 4–5 |
| CH-2 | 3.53 | dd | 2–3 | |
| CH2-1′ (Fru) | 3.67 | s | ||
| CH-3 | 3.74 | t | 3–4 | |
| CH2-6,6′ | 3.81 | m | ||
| CH-5 | 3.85 | dd | ||
| CH-5′ | 3.87 | dd | ||
| CH-4′ | 4.05 | t | 4′–5′ | |
| CH-3′ | 4.19 | d | 3′–4′ | |
| CH-1 (Gluc) | 5.42 | d | 1–2 | |
| Organic acids | ||||
| Acrylic acid | CH2-3 | 5.61 | dd | 3–3 |
| CH2-3 | 6.02 | dd | ||
| CH-2 | 6.13 | dd | 2–3 | |
| Amino acids | ||||
| Alanine (Ala) | CH3-3 | 1.51 | d | |
| CH-2 | 3.75 | q | 2–3 | |
| Arginine (Arg) | CH2-4 | 1.7 | m | 4–3,5 |
| CH2-3 | 1.93 | m | 3–4,2 | |
| CH2-5 | 3.26 | t | ||
| CH2-2 | 3.71 | t | ||
| Aspartic acid (Asp) | CH-3 | 2.82 | dd | 3–3 |
| CH-3 | 2.96 | dd | ||
| CH-2 | 3.95 | dd | 2–3 | |
| Aminobutyric acid (GABA) | CH2-3 | 1.85 | m | 3–2,4 |
| CH2-2 | 2.25 | t | ||
| CH2-4 | 3.22 | t | ||
| Glutamic acid (Glu) * | CH2-6 | 2.05 | m | |
| CH2-6 | 2.15 | m | 6–7, 6–4 | |
| CH2-7 | 2.4 | m | ||
| CH2-4 | 3.72 | dd | ||
| Threonine (Thr) | CH-3 | 1.36 | d | |
| CH3-4 | 4.12 | m | ||
| Lipids: fatty acids | ||||
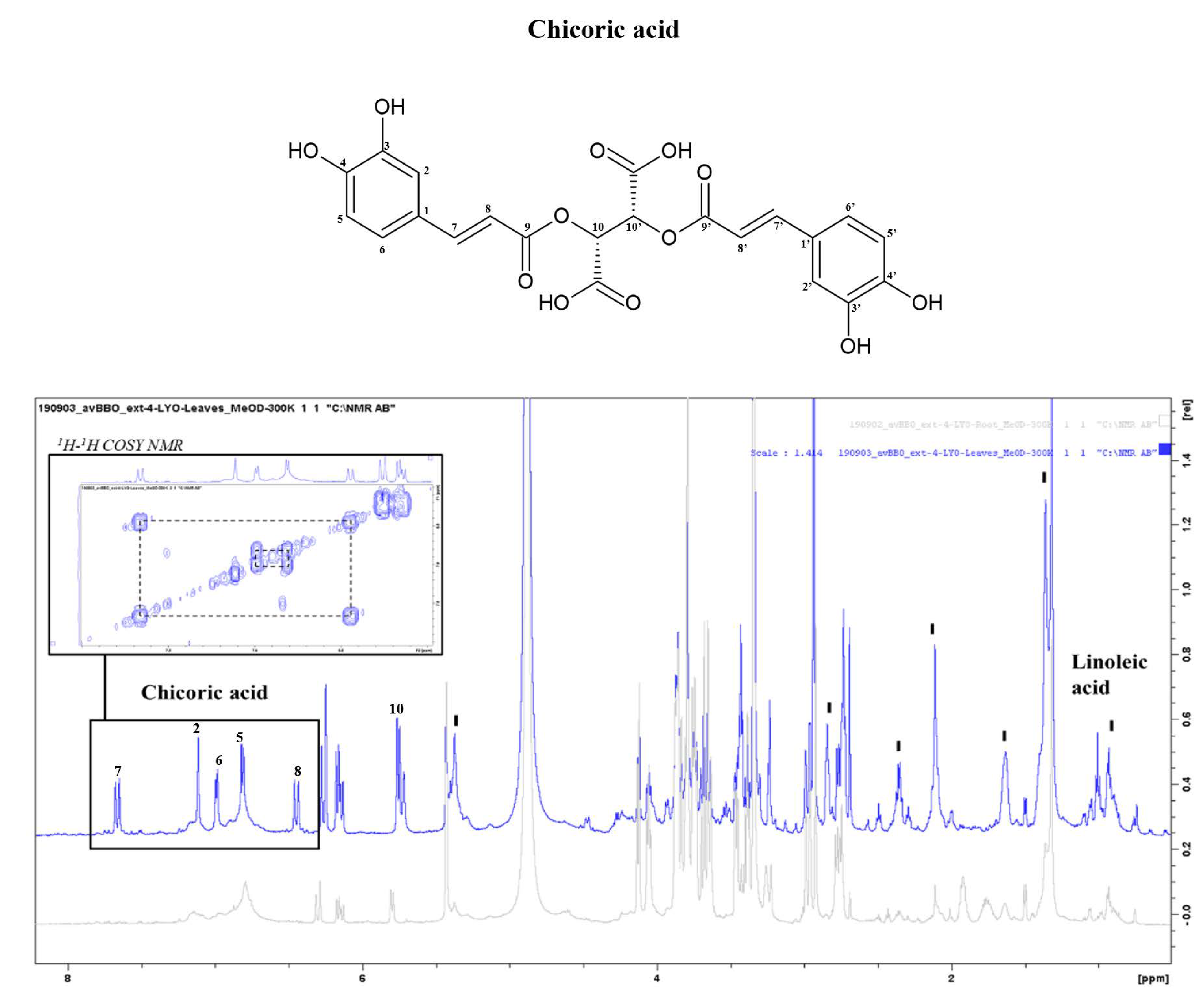
| α-linoleic acid (C18:2 Δ 9,12) | CH3-18 | 0.9 | t | 18–17 |
| CH2-4,5,6,7,15,16,17 | 1.32 | m | 4,7–3,8; 15,17–14,18 | |
| CH2-3 | 1.58 | m | 3–2,4 | |
| CH2-8,14 | 2.06 | m | 8–7,9;14–13,15 | |
| CH2-2 | 2.17 | t | ||
| CH2-11 | 2.78 | t | 11–10,12 | |
| =CH-9,10,12,13 | 5.39 | m | 9,10–8,11; 12,13–11,14 | |
| Oleic acid (C18:1 Δ 9) | CH3-18 | 0.92 | t | 18–17 |
| CH2-4,5,6,7,12,13,14, 15,16,17 | 1.34 | m | 4,7–3,8; 12–11;17–18 | |
| CH2-3 | 1.64 | m | 3–2,4 | |
| CH2-8,11 | 2.09 | m | 8–7,9;11–10,12 | |
| CH2-2 | 2.34 | t | ||
| =CH-9,10 | 5.37 | m | 9,10–8,11; | |
| Phenols | ||||
| Chicoric acid * | CH-10,10′ | 5.57 | s | |
| CH-8,8′ | 6.49 | d | 8–7 | |
| CH-5,5′ | 6.97 | d | 5–6 | |
| CH-6,6′ | 7.17 | d | ||
| CH-2,2′ | 7.25 | d | ||
| CH-7,7′ | 7.73 | d | ||
| Ascophyllum nodosum | ||||
| Carbohydrates | ||||
| Mannitol (Mann) | CH2-1 | 3.66 | dd | 11 |
| CH-2 | 3.72 | m | 2–3 | |
| CH-3 | 3.8 | d | ||
| CH2-1 | 3.83 | dd | ||
| α-D-glucose (α-Gluc) | CH-4 | m | ||
| CH-2 | 3.39 | dd | ||
| CH2-6 | m | |||
| CH-3 | m | |||
| CH-5, CH2-6 | m | |||
| CH-1 | 5.14 | d | 1–2 | |
| β-D-glucose (β-Gluc) | CH-2 | 3.27 | dd | 2–3 |
| CH-4 | m | 4–5 | ||
| CH-3,5 | 3.59 | m | 3–4, 5–6 | |
| CH2-6 | m | |||
| CH2-6 | 3.95 | dd | ||
| CH-1 | 4.5 | d | 1–2 | |
| Amino acids | ||||
| Alanine (Ala) | CH3-3 | 1.5 | ||
| CH-2 | 3.62 | 3–4 | ||
| Threonine (Thr) | CH-3 | 1.43 | d | |
| CH3-4 | 4.23 | m | ||
| Glutamic acid (Glu) | CH2-6 | 2.03 | m | |
| CH2-6 | 2.17 | m | 6–7, 6–4 | |
| CH2-7 | 2.37 | m | ||
| CH2-4 | 3.81 | dd | ||
| Lipids: fatty acids | ||||
| α-Linoleic acid (C18:2 Δ 9,12) | CH3-18 | 0.92 | t | 18–17 |
| CH2-4,5,6,7,15,16,17 | 1.35 | m | 4,7–3,8; 15,17–14,18 | |
| CH2-3 | 1.63 | m | 3–2,4 | |
| CH2-8,14 | 2.06 | m | 8–7,9;14–13,15 | |
| CH2-2 | 2.35 | t | ||
| CH2-11 | 2.81 | t | 11–10,12 | |
| =CH-9,10,12,13 | 5.35 | m | 9,10–8,11; 12,13–11,14 | |
| Oleic acid (C18:1 Δ 9) | CH3-18 | 0.92 | t | 18-17 |
| CH2-4,5,6,7,12,13,14, 15,16,17 | 1.35 | m | 4,7–3,8; 12–11;17–18 | |
| CH2-3 | 1.62 | m | 3–2,4 | |
| CH2-8,11 | 2.06 | m | 8–7,9;11–10,12 | |
| CH2-2 | 2.34 | t | ||
| =CH-9,10 | 5.37 | m | 9,10–8,11; | |
| Phenols | ||||
| Phlorotannin | 5.94 | |||
| 6.08 | ||||
| 6.19 | ||||
| 6.35 | ||||
| Caffeic acid | =CH-10 | 6.33 | d | |
| CH-3 | 6.91 | d | 3–4 | |
| CH-4 | 7.05 | dd | ||
| CH-6 | 7.15 | d | ||
| =CH-9 | 7.24 | d | 4–9 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruno, A.; Velders, A.H.; Biasone, A.; Li Vigni, M.; Mondelli, D.; Miano, T. Chemical Composition, Biomolecular Analysis, and Nuclear Magnetic Resonance Spectroscopic Fingerprinting of Posidonia oceanica and Ascophyllum nodosum Extracts. Metabolites 2023, 13, 170. https://doi.org/10.3390/metabo13020170
Bruno A, Velders AH, Biasone A, Li Vigni M, Mondelli D, Miano T. Chemical Composition, Biomolecular Analysis, and Nuclear Magnetic Resonance Spectroscopic Fingerprinting of Posidonia oceanica and Ascophyllum nodosum Extracts. Metabolites. 2023; 13(2):170. https://doi.org/10.3390/metabo13020170
Chicago/Turabian StyleBruno, Angelica, Aldrik H. Velders, Alessandro Biasone, Mario Li Vigni, Donato Mondelli, and Teodoro Miano. 2023. "Chemical Composition, Biomolecular Analysis, and Nuclear Magnetic Resonance Spectroscopic Fingerprinting of Posidonia oceanica and Ascophyllum nodosum Extracts" Metabolites 13, no. 2: 170. https://doi.org/10.3390/metabo13020170
APA StyleBruno, A., Velders, A. H., Biasone, A., Li Vigni, M., Mondelli, D., & Miano, T. (2023). Chemical Composition, Biomolecular Analysis, and Nuclear Magnetic Resonance Spectroscopic Fingerprinting of Posidonia oceanica and Ascophyllum nodosum Extracts. Metabolites, 13(2), 170. https://doi.org/10.3390/metabo13020170