


An Egg White-Derived Peptide Enhances Systemic Insulin Sensitivity and Modulates Markers of Non-Alcoholic Fatty Liver Disease in Obese, Insulin Resistant Mice

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Body Weight, Body Composition and Sample Collection
2.3. Adipose Tissue Organ Culture
2.4. Preadipocyte Cell Culture
2.5. Hyperinsulinemic-Euglycemic Clamp
2.6. Oral Glucose Tolerance Test (OGTT)
2.7. Liver Triglyceride and Cholesterol Content
2.8. PPARγ DNA Binding Activity
2.9. Plasma Biochemical Analysis
2.10. Protein Extraction and Western Blot
2.11. Histology
2.12. Quantitative PCR (qPCR)
2.13. Statistical Analysis
3. Results
3.1. EHW Effects on WAT Lipolytic Pathway from Obese, Insulin Resistant Rats
3.2. Peptide 2 and Rosiglitazone Effects in HFD Induced Obese and Insulin Resistant Mice
3.2.1. Food Intake, Body Composition and Tissue Weight
3.2.2. Plasma Biochemical Parameters
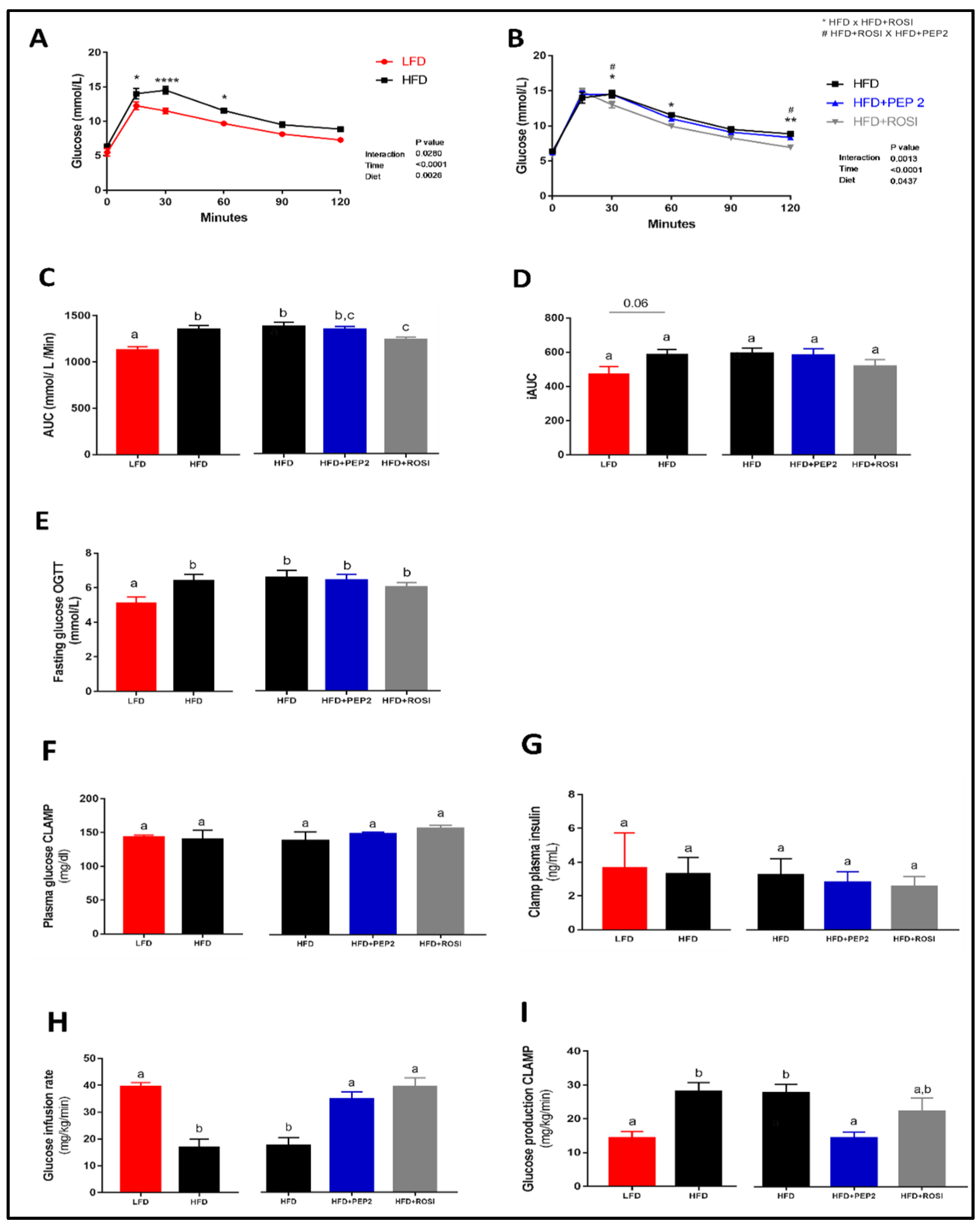
3.2.3. Glucose Homeostasis and Systemic Insulin Sensitivity
3.2.4. WAT Regulation by Insulin: Lipolysis and AKT
3.2.5. PPARγ Activation, Adipocyte Size and Adipogenesis Markers
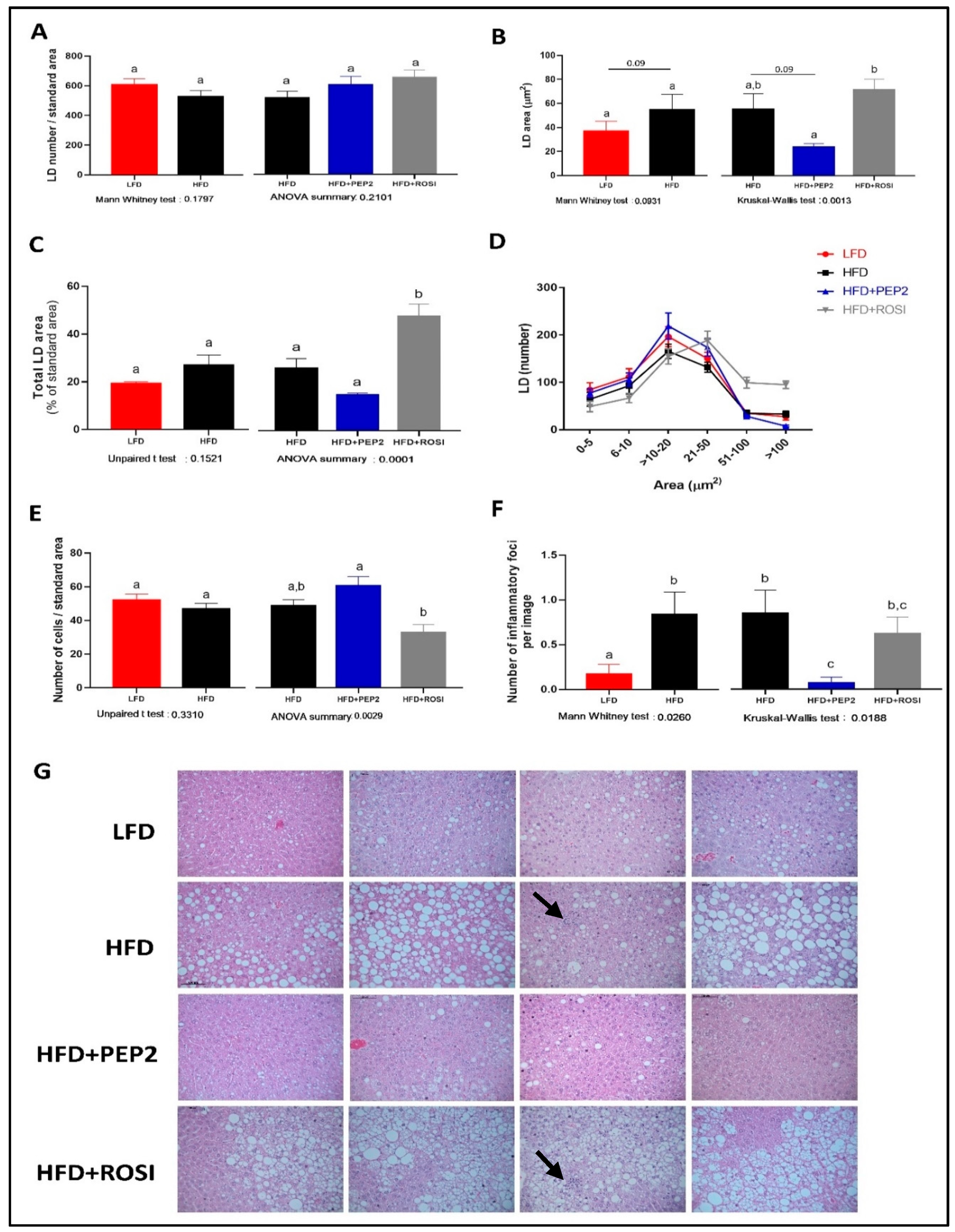
3.2.6. Liver Characterization
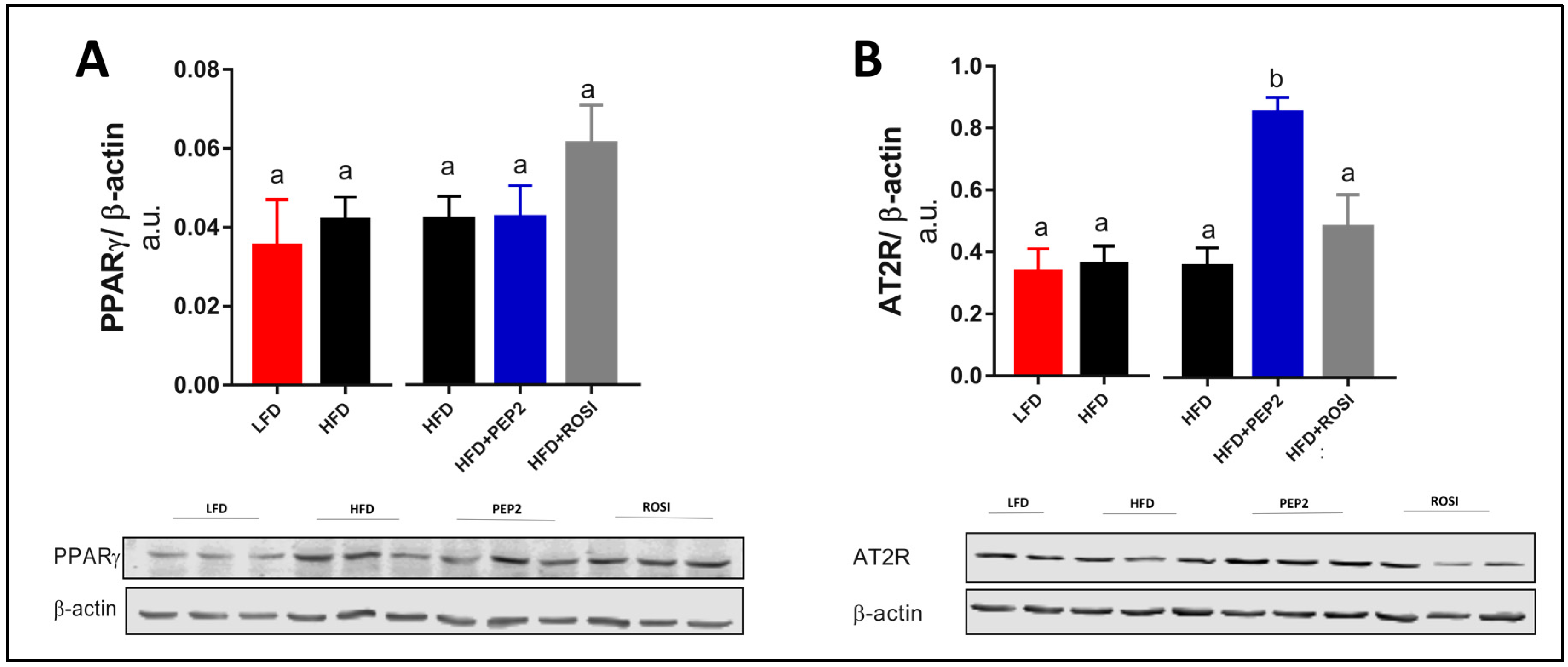
3.2.7. Liver PPARγ and AT2R and Insulin Signaling
3.2.8. Lipid Metabolism, Inflammation, and Fibrosis Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Shankaran, M.; Yoshino, M.; Schweitzer, G.G.; Chondronikola, M.; Beals, J.W.; Okunade, A.L.; Patterson, B.W.; Nyangau, E.; Field, T.; et al. Insulin resistance drives hepatic de novo lipogenesis in nonalcoholic fatty liver disease. J. Clin. Investig. 2020, 130, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Softic, S.; Cohen, D.E.; Kahn, C.R. Role of Dietary Fructose and Hepatic De Novo Lipogenesis in Fatty Liver Disease. Dig. Dis. Sci. 2016, 61, 1282–1293. [Google Scholar] [CrossRef] [Green Version]
- Godoy-Matos, A.F.; Silva Junior, W.S.; Valerio, C.M. NAFLD as a continuum: From obesity to metabolic syndrome and diabetes. Diabetol. Metab. Syndr. 2020, 12, 60. [Google Scholar] [CrossRef]
- Colca, J.R.; Scherer, P.E. The metabolic syndrome, thiazolidinediones, and implications for intersection of chronic and inflammatory disease. Mol. Metab. 2022, 55, 101409. [Google Scholar] [CrossRef]
- Boettcher, E.; Csako, G.; Pucino, F.; Wesley, R.; Loomba, R. Meta-analysis: Pioglitazone improves liver histology and fibrosis in patients with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2012, 35, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Cassader, M.; Paschetta, E.; Gambino, R. Thiazolidinediones and Advanced Liver Fibrosis in Nonalcoholic Steatohepatitis: A Meta-analysis. JAMA Intern. Med. 2017, 177, 633–640. [Google Scholar] [CrossRef]
- FUFOSE. Scientific concepts of functional foods in Europe-consensus document. Br. J. Nutr. 1999, 81, S1–S27. [Google Scholar] [CrossRef] [Green Version]
- Jahandideh, F.; Chakrabarti, S.; Majumder, K.; Li, Q.; Panahi, S.; Morton, J.S.; Davidge, S.T.; Wu, J. Egg white protein hydrolysate reduces blood pressure, improves vascular relaxation and modifies aortic angiotensin II receptors expression in spontaneously hypertensive rats. J. Funct. Foods 2016, 27, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Jahandideh, F.; Zani, S.C.C.; Son, M.; Proctor, S.D.; Davidge, S.T.; Chan, C.B.; Wu, J. Egg white hydrolysate enhances insulin sensitivity in high-fat diet-induced insulin-resistant rats via Akt activation. Br. J. Nutr. 2019, 122, 14–24. [Google Scholar] [CrossRef]
- Jahandideh, F.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Egg white hydrolysate shows insulin mimetic and sensitizing effects in 3T3-F442A pre-adipocytes. PLoS ONE 2017, 12, e0185653. [Google Scholar] [CrossRef] [Green Version]
- Majumder, K.; Chakrabarti, S.; Morton, J.S.; Panahi, S.; Kaufman, S.; Davidge, S.T.; Wu, J. Egg-derived tri-peptide IRW exerts antihypertensive effects in spontaneously hypertensive rats. PLoS ONE 2013, 8, e82829. [Google Scholar] [CrossRef] [Green Version]
- de Campos Zani, S.C.; Son, M.; Bhullar, K.S.; Chan, C.B.; Wu, J. IRW (Isoleucine-Arginine-Tryptophan) Improves Glucose Tolerance in High Fat Diet Fed C57BL/6 Mice via Activation of Insulin Signaling and AMPK Pathways in Skeletal Muscle. Biomedicines 2022, 10, 1235. [Google Scholar] [CrossRef]
- Jahandideh, F.; Liu, P.; Wu, J. Purification and identification of adipogenic-differentiating peptides from egg white hydrolysate. Food Chem. 2018, 259, 25–30. [Google Scholar] [CrossRef]
- Rocha, A.L.; de Lima, T.I.; de Souza, G.P.; Correa, R.O.; Ferrucci, D.L.; Rodrigues, B.; Lopes-Ramos, C.; Nilsson, D.; Knittel, T.L.; Castro, P.R.; et al. Enoxacin induces oxidative metabolism and mitigates obesity by regulating adipose tissue miRNA expression. Sci. Adv. 2020, 6, eabc6250. [Google Scholar] [CrossRef]
- Mighiu, P.I.; Yue, J.T.; Filippi, B.M.; Abraham, M.A.; Chari, M.; Lam, C.K.; Yang, C.S.; Christian, N.R.; Charron, M.J.; Lam, T.K. Hypothalamic glucagon signaling inhibits hepatic glucose production. Nat. Med. 2013, 19, 766–772. [Google Scholar] [CrossRef]
- Yue, J.T.; Abraham, M.A.; Bauer, P.V.; LaPierre, M.P.; Wang, P.; Duca, F.A.; Filippi, B.M.; Chan, O.; Lam, T.K. Inhibition of glycine transporter-1 in the dorsal vagal complex improves metabolic homeostasis in diabetes and obesity. Nat. Commun. 2016, 7, 13501. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ferri, F.; Carotti, S.; Carpino, G.; Mischitelli, M.; Cantafora, A.; Molinaro, A.; Argenziano, M.E.; Parisse, S.; Corsi, A.; Riminucci, M.; et al. The Propensity of the Human Liver to Form Large Lipid Droplets Is Associated with PNPLA3 Polymorphism, Reduced INSIG1 and NPC1L1 Expression and Increased Fibrogenetic Capacity. Int. J. Mol. Sci. 2021, 22, 6100. [Google Scholar] [CrossRef]
- Yersiz, H.; Lee, C.; Kaldas, F.M.; Hong, J.C.; Rana, A.; Schnickel, G.T.; Wertheim, J.A.; Zarrinpar, A.; Agopian, V.G.; Gornbein, J.; et al. Assessment of hepatic steatosis by transplant surgeon and expert pathologist: A prospective, double-blind evaluation of 201 donor livers. Liver Transpl. 2013, 19, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M. Pathology of fatty liver disease. Mod. Pathol. 2007, 20 (Suppl. S1), S40–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdouse, A.; Agrawal, R.R.; Gao, M.A.; Jiang, H.; Blaner, W.S.; Clugston, R.D. Alcohol induced hepatic retinoid depletion is associated with the induction of multiple retinoid catabolizing cytochrome P450 enzymes. PLoS ONE 2022, 17, e0261675. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wai-Sun Wong, V.; Dufour, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Sakurai, Y.; Kubota, N.; Yamauchi, T.; Kadowaki, T. Role of Insulin Resistance in MAFLD. Int. J. Mol. Sci. 2021, 22, 4156. [Google Scholar] [CrossRef]
- Garces-Rimon, M.; Gonzalez, C.; Vera, G.; Uranga, J.A.; Lopez-Fandino, R.; Lopez-Miranda, V.; Miguel, M. Pepsin egg white hydrolysate improves glucose metabolism complications related to metabolic syndrome in Zucker fatty rats. Nutrients 2018, 10, 441. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Ma, Y.; Alsaggar, M.; Liu, D. Dual Outcomes of Rosiglitazone Treatment on Fatty Liver. AAPS J. 2016, 18, 1023–1031. [Google Scholar] [CrossRef]
- Lee, S.M.; Muratalla, J.; Diaz-Ruiz, A.; Remon-Ruiz, P.; McCann, M.; Liew, C.W.; Kineman, R.D.; Cordoba-Chacon, J. Rosiglitazone Requires Hepatocyte PPARgamma Expression to Promote Steatosis in Male Mice with Diet-Induced Obesity. Endocrinology 2021, 162, bqab175. [Google Scholar] [CrossRef]
- Vidal-Puig, A.; Jimenez-Linan, M.; Lowell, B.B.; Hamann, A.; Hu, E.; Spiegelman, B.; Flier, J.S.; Moller, D.E. Regulation of PPAR gamma gene expression by nutrition and obesity in rodents. J. Clin. Investig. 1996, 97, 2553–2561. [Google Scholar] [CrossRef] [Green Version]
- Rosenstengel, S.; Stoeppeler, S.; Bahde, R.; Spiegel, H.U.; Palmes, D. Type of steatosis influences microcirculation and fibrogenesis in different rat strains. J. Investig. Surg. 2011, 24, 273–282. [Google Scholar] [CrossRef]
- Najt, C.P.; Senthivinayagam, S.; Aljazi, M.B.; Fader, K.A.; Olenic, S.D.; Brock, J.R.; Lydic, T.A.; Jones, A.D.; Atshaves, B.P. Liver-specific loss of Perilipin 2 alleviates diet-induced hepatic steatosis, inflammation, and fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G726–G738. [Google Scholar] [CrossRef] [Green Version]
- Garces-Rimon, M.; Gonzalez, C.; Uranga, J.A.; Lopez-Miranda, V.; Lopez-Fandino, R.; Miguel, M. Pepsin egg white hydrolysate ameliorates obesity-related oxidative stress, inflammation and steatosis in Zucker fatty rats. PLoS ONE 2016, 11, e0151193. [Google Scholar] [CrossRef] [Green Version]
- Lemus-Conejo, A.; Grao-Cruces, E.; Toscano, R.; Varela, L.M.; Claro, C.; Pedroche, J.; Millan, F.; Millan-Linares, M.C.; Montserrat-de la Paz, S. A lupine (Lupinus angustifolious L.) peptide prevents non-alcoholic fatty liver disease in high-fat-diet-induced obese mice. Food Funct. 2020, 11, 2943–2952. [Google Scholar] [CrossRef] [Green Version]
- Dumeus, S.; Shibu, M.A.; Lin, W.T.; Wang, M.F.; Lai, C.H.; Shen, C.Y.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Bioactive Peptide Improves Diet-Induced Hepatic Fat Deposition and Hepatocyte Proinflammatory Response in SAMP8 Ageing Mice. Cell Physiol. Biochem. 2018, 48, 1942–1952. [Google Scholar] [CrossRef]
- Song, C.; Lv, W.; Li, Y.; Nie, P.; Lu, J.; Geng, Y.; Heng, Z.; Song, L. Alleviating the effect of quinoa and the underlying mechanism on hepatic steatosis in high-fat diet-fed rats. Nutr. Metab. 2021, 18, 106. [Google Scholar] [CrossRef]
- Steckelings, U.M.; Widdop, R.E.; Sturrock, E.D.; Lubbe, L.; Hussain, T.; Kaschina, E.; Unger, T.; Hallberg, A.; Carey, R.M.; Sumners, C. The Angiotensin AT(2) Receptor: From a Binding Site to a Novel Therapeutic Target. Pharmacol. Rev. 2022, 74, 1051–1135. [Google Scholar] [CrossRef]
- Goh, G.B.; Pagadala, M.R.; Dasarathy, J.; Unalp-Arida, A.; Sargent, R.; Hawkins, C.; Sourianarayanane, A.; Khiyami, A.; Yerian, L.; Pai, R.; et al. Renin-angiotensin system and fibrosis in non-alcoholic fatty liver disease. Liver Int. 2015, 35, 979–985. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, J.; Li, P.; Wang, Y.; Zhang, Y. Up-regulation of components of the renin-angiotensin system in liver fibrosis in the rat induced by CCL(4). Res. Vet. Sci. 2013, 95, 54–58. [Google Scholar] [CrossRef]
- Nabeshima, Y.; Tazuma, S.; Kanno, K.; Hyogo, H.; Iwai, M.; Horiuchi, M.; Chayama, K. Anti-fibrogenic function of angiotensin II type 2 receptor in CCl4-induced liver fibrosis. Biochem. Biophys. Res. Commun. 2006, 346, 658–664. [Google Scholar] [CrossRef]
- Chai, W.; Wang, W.; Dong, Z.; Cao, W.; Liu, Z. Angiotensin II receptors modulate muscle microvascular and metabolic responses to insulin in vivo. Diabetes 2011, 60, 2939–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrion, D.; Kubis, N.; Levy, B.I. Physiological and pathophysiological functions of the AT(2) subtype receptor of angiotensin II: From large arteries to the microcirculation. Hypertension 2001, 38, 1150–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Suzuki, J.; Tsujioka, S.; Sasaki, M.; Gomori, A.; Shirakura, T.; Hirose, H.; Ito, M.; Ishihara, A.; Iwaasa, H.; et al. Longitudinal analysis of murine steatohepatitis model induced by chronic exposure to high-fat diet. Hepatol. Res. 2007, 37, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, M.N.B.; Veidal, S.S.; Christoffersen, C.; Jelsing, J.; Rigbolt, K.T.G. Molecular Characterization of Microvesicular and Macrovesicular Steatosis Shows Widespread Differences in Metabolic Pathways. Lipids 2019, 54, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, Y.; Suemitsu, E.; Kajimoto, K.; Sato, Y.; Akhter, A.; Sakurai, Y.; Hatakeyama, H.; Hyodo, M.; Kaji, N.; Baba, Y.; et al. Hepatic Monoacylglycerol O-acyltransferase 1 as a Promising Therapeutic Target for Steatosis, Obesity, and Type 2 Diabetes. Mol. Ther. Nucleic Acids 2014, 3, e154. [Google Scholar] [CrossRef]
- Lee, Y.J.; Ko, E.H.; Kim, J.E.; Kim, E.; Lee, H.; Choi, H.; Yu, J.H.; Kim, H.J.; Seong, J.K.; Kim, K.S.; et al. Nuclear receptor PPARgamma-regulated monoacylglycerol O-acyltransferase 1 (MGAT1) expression is responsible for the lipid accumulation in diet-induced hepatic steatosis. Proc. Natl. Acad. Sci. USA 2012, 109, 13656–13661. [Google Scholar] [CrossRef] [Green Version]
- Lutkewitte, A.J.; Singer, J.M.; Shew, T.M.; Martino, M.R.; Hall, A.M.; He, M.; Finck, B.N. Multiple antisense oligonucleotides targeted against monoacylglycerol acyltransferase 1 (Mogat1) improve glucose metabolism independently of Mogat1. Mol. Metab. 2021, 49, 101204. [Google Scholar] [CrossRef]
- Lutkewitte, A.J.; McCommis, K.S.; Schweitzer, G.G.; Chambers, K.T.; Graham, M.J.; Wang, L.; Patti, G.J.; Hall, A.M.; Finck, B.N. Hepatic monoacylglycerol acyltransferase 1 is induced by prolonged food deprivation to modulate the hepatic fasting response. J. Lipid Res. 2019, 60, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Carreres, L.; Jilkova, Z.M.; Vial, G.; Marche, P.N.; Decaens, T.; Lerat, H. Modeling Diet-Induced NAFLD and NASH in Rats: A Comprehensive Review. Biomedicines 2021, 9, 378. [Google Scholar] [CrossRef]
- Charlton, M.; Krishnan, A.; Viker, K.; Sanderson, S.; Cazanave, S.; McConico, A.; Masuoko, H.; Gores, G. Fast food diet mouse: Novel small animal model of NASH with ballooning, progressive fibrosis, and high physiological fidelity to the human condition. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G825–G834. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Lao, F.; Pan, X.; Wu, J. Food Protein-Derived Antioxidant Peptides: Molecular Mechanism, Stability and Bioavailability. Biomolecules 2022, 12, 1622. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide 2 | |
|---|---|
| Amino acid sequence | QAMPFRVTEQE |
| Number of amino acids | 11 |
| Theoretical molecular weight (g/mol) * | 1335.50 |
| Observed molecular weight (g/mol) | 1335.8 |
| Theoretical isoelectric point * | 4.53 |
| Grand average of hydropathicity (GRAVY) * | −0.918 |
| Hydrophobicity * | 22 |
| Terminus modifications | None |
| Net charge at pH 7.0 * | −1 |
| HFD | HFD + 4% EWH | |
|---|---|---|
| Plasma (fasting) | ||
| Resistin (pg/mL) | 1047 ± 84.55 | 1011 ± 82.99 |
| Adiponectin (ng/mL) | 55,853 ± 2832 | 60,270 ± 7367 |
| eWAT | ||
| Resistin (pg/mL) | 175.1 ± 15.67 | 166.5 ± 11.14 |
| Adiponectin (ng/mL) | 1002 ± 28.44 | 1044 ± 19.54 |
| rWAT | ||
| Resistin (pg/mL) | 172.3 ± 20.14 | 175.1 ± 15.67 |
| Adiponectin (ng/mL) | 1033 ± 25 | 1026 ± 28.73 |
| LFD | HFD | HFD + PEP2 | HFD + ROSI | |
|---|---|---|---|---|
| Body composition (not fasted) | ||||
| Initial BW (g) (week 6) | 29.1 ± 0.6 # | 33.9 + 0.7 a | 32.9 + 0.6 a | 33.5 + 0.7 a |
| Final BW (g) | 33.4 ± 0.7 # | 42.5 ± 0.7 a | 41.4 ± 0.5 a.b | 39.4 ± 0.8 b |
| BW gain (%) (week 6–13) | 15.4 ± 1.0 # | 25.0 ± 1.3 a | 26.2 ± 1.2 a | 17.7 ± 1.5 b |
| Final fat mass (% BW) | 25.6 ± 1.3 # | 38.5 ± 1.1 a | 38.6 ± 0.6 a | 36.1 ± 1.1 a |
| Final lean mass(% BW) | 65.9 ± 1.2 # | 54.2 ± 1.0 a | 54.3 ± 0.6 a | 56.5 ± 1.0 a |
| Tissue weight (g/BW) | ||||
| eWAT | 0.039 ± 0.0029 # | 0.059 ± 0.0029 a | 0.061 ± 0.0014 a | 0.057 ± 0.0025 a |
| rWAT | 0.017 ± 0.00079 # | 0.028 ± 0.00084 a | 0.025 ± 0.00096 a,b | 0.022 ± 0.0014 b |
| iWAT | 0.046 ± 0.0070 # | 0.073 ± 0.0049 a | 0.065 ± 0.0027 a | 0.062 ± 0.0040 a |
| Liver | 0.036 ± 0.0019 | 0.033 ± 0.0015 a | 0.032 ± 0.0007 a | 0.044 ± 0.0015 b |
| Plasma (fasting) | ||||
| Glucose (mmo/L) | 4.5 ± 0.3 # | 5.9 ± 0.3 a | 5.3 ± 0.3 a | 5.6 ± 0.2 a |
| Insulin (ng/mL) | 0.5 ± 0.1 ^ | 1.0 ± 0.1 a | 0.9 ± 0.1 a | 0.6 ± 0.1 a |
| HOMA-IR | 0.8 ± 0.2 # | 2.4 ± 0.6 a | 1.8 ± 0.3 a | 1.9 ± 0.3 a |
| NEFA (mEq/L) | 0.4 ± 0.02 | 0.4 ± 0.02 a | 0.5 ± 0.03 a | 0.5 ± 0.04 a |
| TG (mg/dL) | 39.4 ± 5.6 | 52.6 ± 5.8 a | 43.2 ± 4.2 a | 41.9 ± 4.6 a |
| Plasma ALT (mU/mL) | 8.4 ± 4.0 # | 28.4 ± 27.1 a | 24.9 ± 15.1 a | 20.9 ± 13.5 a |
| Liver content (mg/g tissue) | ||||
| TG | 46.8 ± 2.3 # | 71.5 ± 11.0 a | 57.6 ± 6.7 a | 132.7 ± 12.1 b |
| Cholesterol | 2.4 ± 0.2 | 2.1 ± 0.2 a | 1.3 ± 0.1 b | 1.8 ± 0.1 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Campos Zani, S.C.; Wang, R.; Veida-Silva, H.; Clugston, R.D.; Yue, J.T.Y.; Mori, M.A.; Wu, J.; Chan, C.B. An Egg White-Derived Peptide Enhances Systemic Insulin Sensitivity and Modulates Markers of Non-Alcoholic Fatty Liver Disease in Obese, Insulin Resistant Mice. Metabolites 2023, 13, 174. https://doi.org/10.3390/metabo13020174
de Campos Zani SC, Wang R, Veida-Silva H, Clugston RD, Yue JTY, Mori MA, Wu J, Chan CB. An Egg White-Derived Peptide Enhances Systemic Insulin Sensitivity and Modulates Markers of Non-Alcoholic Fatty Liver Disease in Obese, Insulin Resistant Mice. Metabolites. 2023; 13(2):174. https://doi.org/10.3390/metabo13020174
Chicago/Turabian Stylede Campos Zani, Stepheny C., Ren Wang, Hellen Veida-Silva, Robin D. Clugston, Jessica T. Y. Yue, Marcelo A. Mori, Jianping Wu, and Catherine B. Chan. 2023. "An Egg White-Derived Peptide Enhances Systemic Insulin Sensitivity and Modulates Markers of Non-Alcoholic Fatty Liver Disease in Obese, Insulin Resistant Mice" Metabolites 13, no. 2: 174. https://doi.org/10.3390/metabo13020174
APA Stylede Campos Zani, S. C., Wang, R., Veida-Silva, H., Clugston, R. D., Yue, J. T. Y., Mori, M. A., Wu, J., & Chan, C. B. (2023). An Egg White-Derived Peptide Enhances Systemic Insulin Sensitivity and Modulates Markers of Non-Alcoholic Fatty Liver Disease in Obese, Insulin Resistant Mice. Metabolites, 13(2), 174. https://doi.org/10.3390/metabo13020174