


Changes in the Urine Metabolomic Profile in Patients Recovering from Severe COVID-19

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Controls
2.3. Urine Sampling
2.4. NMR Data Acquisition
2.5. Data Normalization
2.6. Data Analysis
3. Results
4. Discussion
4.1. Metabolic Changes in Urine
4.2. Multivariate and Discriminatory Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adil, M.T.; Rahman, R.; Whitelaw, D.; Jain, V.; Al-Taan, O.; Rashid, F.; Munasinghe, A.; Jambulingam, P. SARS-CoV-2 and the pandemic of COVID-19. Postgrad. Med. J. 2021, 97, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, H.K.; Libby, P.; Ridker, P.M. COVID-19—A vascular disease. Trends Cardiovasc. Med. 2021, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Galanopoulos, M.; Gkeros, F.; Doukatas, A.; Karianakis, G.; Pontas, C.; Tsoukalas, N.; Viazis, N.; Liatsos, C.; Mantzaris, G.J. COVID-19 pandemic: Pathophysiology and manifestations from the gastrointestinal tract. World J. Gastroenterol. 2020, 26, 4579. [Google Scholar] [CrossRef] [PubMed]
- Liptak, P.; Duricek, M.; Rosolanka, R.; Ziacikova, I.; Kocan, I.; Uhrik, P.; Grendar, M.; Hrnciarova, M.; Bucova, P.; Galo, D.; et al. Gastrointestinal sequalae months after severe acute respiratory syndrome corona virus 2 infection: A prospective, observational study. Eur. J. Gastroenterol. Hepatol. 2022, 34, 925–932. [Google Scholar] [CrossRef]
- Liptak, P.; Nosakova, L.; Rosolanka, R.; Skladany, L.; Banovcin, P. Acute-on-chronic liver failure in patients with severe acute respiratory syndrome coronavirus 2 infection. World J. Hepatol. 2023, 15, 41–51. [Google Scholar] [CrossRef]
- Shen, T.; Wang, T. Metabolic Reprogramming in COVID-19. Int. J. Mol. Sci. 2021, 22, 11475. [Google Scholar] [CrossRef]
- Xiao, N.; Nie, M.; Pang, H.; Wang, B.; Hu, J.; Meng, X.; Li, K.; Ran, X.; Long, Q.; Deng, H.; et al. Integrated cytokine and metabolite analysis reveals immunometabolic reprogramming in COVID-19 patients with therapeutic implications. Nat. Commun. 2021, 12, 1618. [Google Scholar] [CrossRef]
- Miller, L.; Berber, E.; Sumbria, D.; Rouse, B.T. Controlling the Burden of COVID-19 by Manipulating Host Metabolism. Viral Immunol. 2022, 35, 24–32. [Google Scholar] [CrossRef]
- Bujak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Kaliszan, R. Metabolomics for laboratory diagnostics. J. Pharm. Biomed. Anal. 2015, 113, 108–120. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Reiman, D.; Layden, B.T.; Dai, Y. MiMeNet: Exploring microbiome-metabolome relationships using neural networks. PLoS Comput. Biol. 2021, 17, e1009021. [Google Scholar] [CrossRef]
- Goyal, A.; Wang, T.; Dubinkina, V.; Maslov, S. Ecology-guided prediction of cross-feeding interactions in the human gut microbiome. Nat. Commun. 2021, 12, 1335. [Google Scholar] [CrossRef]
- Mallick, H.; Franzosa, E.A.; Mclver, L.J.; Banerjee, S.; Sirota-Madi, A.; Kostic, A.D.; Clish, C.B.; Vlamakis, H.; Xavier, R.J.; Huttenhower, C. Predictive metabolomic profiling of microbial communities using amplicon or metagenomic sequences. Nat. Commun. 2019, 10, 3136. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Goyal, A.; Dubinkina, V.; Maslov, S. Evidence for a multi-level trophic organization of the human gut microbiome. PLoS Comput. Biol. 2019, 15, e1007524. [Google Scholar] [CrossRef] [Green Version]
- Ke, S.; Weiss, S.T.; Liu, Y.Y. Dissecting the role of the human microbiome in COVID-19 via metagenome-assembled genomes. Nat. Commun. 2022, 13, 5235. [Google Scholar] [CrossRef]
- Hasan, M.R.; Suleiman, M.; Pérez-López, A. Metabolomics in the Diagnosis and Prognosis of COVID-19. Front. Genet. 2021, 12, 1358. [Google Scholar] [CrossRef]
- Bi, X.; Liu, W.; Ding, X.; Liang, S.; Zheng, Y.; Zhu, X.; Quan, S.; Yi, X.; Xiang, N.; Du, J.; et al. Proteomic and metabolomic profiling of urine uncovers immune responses in patients with COVID-19. Cell Rep. 2022, 38, 110271. [Google Scholar] [CrossRef]
- Dewulf, J.P.; Martin, M.; Marie, S.; Oguz, F.; Belkhir, L.; De Greef, J.; Yombi, J.C.; Wittebole, X.; Laterre, P.-F.; Jadoul, M.; et al. Urine metabolomics links dysregulation of the tryptophan-kynurenine pathway to inflammation and severity of COVID-19. Sci. Rep. 2022, 12, 9959. [Google Scholar] [CrossRef]
- Liptak, P.; Baranovicova, E.; Rosolanka, R.; Simekova, K.; Bobcakova, A.; Vysehradsky, R.; Duricek, M.; Dankova, Z.; Kapinova, A.; Dvorska, D.; et al. Persistence of Metabolomic Changes in Patients during Post-COVID Phase: A Prospective, Observational Study. Metabolites 2022, 12, 641. [Google Scholar] [CrossRef]
- Ryan, D.; Robards, K.; Prenzler, P.D.; Kendall, M. Recent and potential developments in the analysis of urine: A review. Anal. Chim. Acta 2011, 684, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Mervant, L.; Tremblay-Franco, M.; Jamin, E.L.; Kesse-Guyot, E.; Galan, P.; Martin, J.-F.; Guéraud, F.; Debrauwer, L. Osmolality-based normalization enhances statistical discrimination of untargeted metabolomic urine analysis: Results from a comparative study. Metabolomics 2021, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50(D1), D622–D631. [Google Scholar] [CrossRef]
- Görling, B.; Bräse, S.; Luy, B. NMR Chemical Shift Ranges of Urine Metabolites in Various Organic Solvents. Metabolites 2016, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tredwell, G.D.; Bundy, J.G.; De Iorio, M.; Ebbels, T.M.D. Modelling the acid/base 1H NMR chemical shift limits of metabolites in human urine. Metabolomics 2016, 12, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulido, N.; Guevara-Morales, J.M.; Rodriguez-López, A.; Pulido, A.; Díaz, J.; Edrada-Ebel, R.A.; Echeverri-Peña, O.Y. 1H-Nuclear Magnetic Resonance Analysis of Urine as Diagnostic Tool for Organic Acidemias and Aminoacidopathies. Metabolites 2021, 11, 891. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49(W1), W388–W396. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, R.; Fernández, L.; Marco, S. Overoptimism in cross-validation when using partial least squares-discriminant analysis for omics data: A systematic study. Anal. Bioanal. Chem. 2018, 410, 5981–5992. [Google Scholar] [CrossRef]
- Chen, T.; Cao, Y.; Zhang, Y.; Liu, J.; Bao, Y.; Wang, C.; Jia, W.; Zhao, A. Random Forest in Clinical Metabolomics for Phenotypic Discrimination and Biomarker Selection. Evid. Based Complement. Alternat. Med. 2013, 2013, e298183. [Google Scholar] [CrossRef] [Green Version]
- Baranovicova, E.; Bobcakova, A.; Vysehradsky, R.; Dankova, Z.; Halasova, E.; Nosal, V.; Lehotsky, J. The Ability to Normalise Energy Metabolism in Advanced COVID-19 Disease Seems to Be One of the Key Factors Determining the Disease Progression—A Metabolomic NMR Study on Blood Plasma. Appl. Sci. 2021, 11, 4231. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Chen, J.; Zuo, X.; Zhang, H.; Deng, A. COVID-19 infection may cause ketosis and ketoacidosis. Diabetes Obes. Metab. 2020, 22, 1935–1941. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Berry-Kravis, E.; Booth, G.; Sanchez, A.C.; Woodbury-Kolb, J. Carnitine Levels and the Ketogenic Diet. Epilepsia 2001, 42, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, J.; Thomsen, J.A.; Olesen, J.H.; Lund, T.M.; Mohr, M.; Clementsen, J.; Nielsen, O.W.; Lund, A.M.; Zschocke, J. Carnitine Levels in Skeletal Muscle, Blood, and Urine in Patients with Primary Carnitine Deficiency During Intermission of l-Carnitine Supplementation. In JIMD Reports; Zschocke, J., Baumgartner, M., Morava, E., Patterson, M., Rahman, S., Peters, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 20, pp. 103–111. Available online: https://doi.org/10.1007/8904_2014_398 (accessed on 1 November 2022).
- Sahajwalla, C.G.; Helton, E.D.; Purich, E.D.; Hoppel, C.L.; Cabana, B.E. Multiple-dose pharmacokinetics and bioequivalence of L-carnitine 330-mg tablet versus 1-g chewable tablet versus enteral solution in healthy adult male volunteers. J. Pharm. Sci. 1995, 84, 627–633. [Google Scholar] [CrossRef]
- Sahajwalla, C.G.; Helton, E.D.; Purich, E.D.; Hoppel, C.L.; Cabana, B.E. Comparison of L-carnitine pharmacokinetics with and without baseline correction following administration of single 20-mg/kg intravenous dose. J. Pharm. Sci. 1995, 84, 634–639. [Google Scholar] [CrossRef]
- Lees, H.J.; Swann, J.R.; Wilson, I.D.; Nicholson, J.K.; Holmes, E. Hippurate: The natural history of a mammalian-microbial cometabolite. J. Proteome Res. 2013, 12, 1527–1546. [Google Scholar] [CrossRef]
- Elliott, P.; Posma, J.M.; Chan, Q.; Garcia-Perez, I.; Wijeyesekera, A.; Bictash, M.; Ebbels, T.M.D.; Ueshima, H.; Zhao, L.; van Horn, L.; et al. Urinary metabolic signatures of human adiposity. Sci. Transl. Med. 2015, 7, 285ra62. [Google Scholar] [CrossRef] [Green Version]
- Zuppi, C.; Messana, I.; Tapanainen, P.; Knip, M.; Vincenzoni, F.; Giardina, B.; Nuutinen, M. Proton nuclear magnetic resonance spectral profiles of urine from children and adolescents with type 1 diabetes. Clin. Chem. 2002, 48, 660–662. [Google Scholar] [CrossRef] [Green Version]
- van Doorn, M.; Vogels, J.; Tas, A.; Van Hoogdalem, E.J.; Burggraaf, J.; Cohen, A.; Van Der Greef, J. Evaluation of metabolite profiles as biomarkers for the pharmacological effects of thiazolidinediones in Type 2 diabetes mellitus patients and healthy volunteers. Br. J. Clin. Pharmacol. 2007, 63, 562–574. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.E.; Eyton-Jones, H.W.; Farnworth, M.J.; Gallagher, R.; Provan, W.M. Effect of intestinal microflora on the urinary metabolic profile of rats: A (1)H-nuclear magnetic resonance spectroscopy study. Xenobiotica Fate Foreign Compd. Biol. Syst. 2002, 32, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Doğan, H.O.; Şenol, O.; Bolat, S.; Yıldız, S.N.; Büyüktuna, S.A.; Sarıismailoğlu, R.; Doğan, K.; Hasbek, M.; Hekim, S.N. Understanding the pathophysiological changes via untargeted metabolomics in COVID-19 patients. J. Med. Virol. 2021, 93, 2340–2349. [Google Scholar] [CrossRef] [PubMed]
- Fisher, O.; Benson, R.A.; Imray ChHE. The Clinical Application of Purine Nucleosides as Biomarkers of Tissue Ischemia and Hypoxia in Humans in Vivo. Biomarkers Med. 2019, 13, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Turgan, N.; Habif, S.; Gülter, C.; Şenol, B.; Mutaf, I.; Özmen, D.; Bayindir, O.; Boydak, B. Urinary hypoxanthine and xanthine levels in acute coronary syndromes. Int. J. Clin. Lab. Res. 1999, 29, 162. [Google Scholar] [CrossRef]
- Hamm, L.L. Renal handling of citrate. Kidney Int. 1990, 38, 728–735. [Google Scholar] [CrossRef] [Green Version]
- Hering-Smith, K.S.; Hamm, L.L. Acidosis and citrate: Provocative interactions. Ann. Transl. Med. 2018, 6, 29. [Google Scholar] [CrossRef]
- Zuckerman, J.M.; Assimos, D.G. Hypocitraturia: Pathophysiology and medical management. Rev. Urol. 2009, 11, 134–144. [Google Scholar]
- Zhu, H.; Bi, D.; Zhang, Y.; Kong, C.; Du, J.; Wu, X.; Wei, Q.; Qin, H. Ketogenic diet for human diseases: The underlying mechanisms and potential for clinical implementations. Signal Transduct. Target. Ther. 2022, 7, 11. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. Formate: The Neglected Member of One-Carbon Metabolism. Annu. Rev. Nutr. 2016, 36, 369–388. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Median (Interquartile Range) | |
|---|---|
| Age [years] | 58 (21) |
| Sex: Female/Male | 7/18 |
| Weight [kg] | 82.6 (26) |
| Height [cm] | 171 (8) |
| BMI | 29 (9) |
| Chronic liver disease | 3 |
| Chronic kidney disease | 3 |
| Ischemic cardiac disease | 3 |
| Diabetes Mellitus | 3 |
| Thyroidal disease | 4 |
| Rheumatic disease | 0 |
| Other relevant | NA |
| Samples A | Samples B | Samples C | |
|---|---|---|---|
| Na+ | 133 (5) | 140 (6) | 139 (2) |
| K+ | 4.1 (0.6) | 4.2 (0.7) | 4.2 (0.4) |
| Cl− | 101 (5) | 104 (5) | 104 (2) |
| Glucose | 7 (1.4) | 5.8 (29), 1 m | 5.5 (1.1) |
| Creatinine | 71 (29) | 69 (23), 2 m | 68 (24) |
| CRP | 102.2 (124.8) | 16.6 (38.1) | 2.2 (4.4) |
| AST | 0.88 (0.71) | 0.93 (1.07), 7 m | 0.575 (0.49) |
| ALT | 0.805 (0.63) | 1.465 (1.325),7 m | 0.89 (1.09) |
| GMT | 0.93 (2.08) | 1.865 (3.605), 3 m | 1.255 (0.46), 1 m |
| Bilirubin | 10.7 (5.4) | 11.1 (4.9), 8 m | 9.5 (6.3) |
| Leukocytes | 6.5 (2.9) | 7.65 (3.5), 1 m | 8.0 (3.1) |
| Hemoglobin [g/L] | 143 (12) | 136 (17), 1 m | 144 (14) |
| Platelets count | 177 (163) | 355 (198), 1 m | 259 (145) |
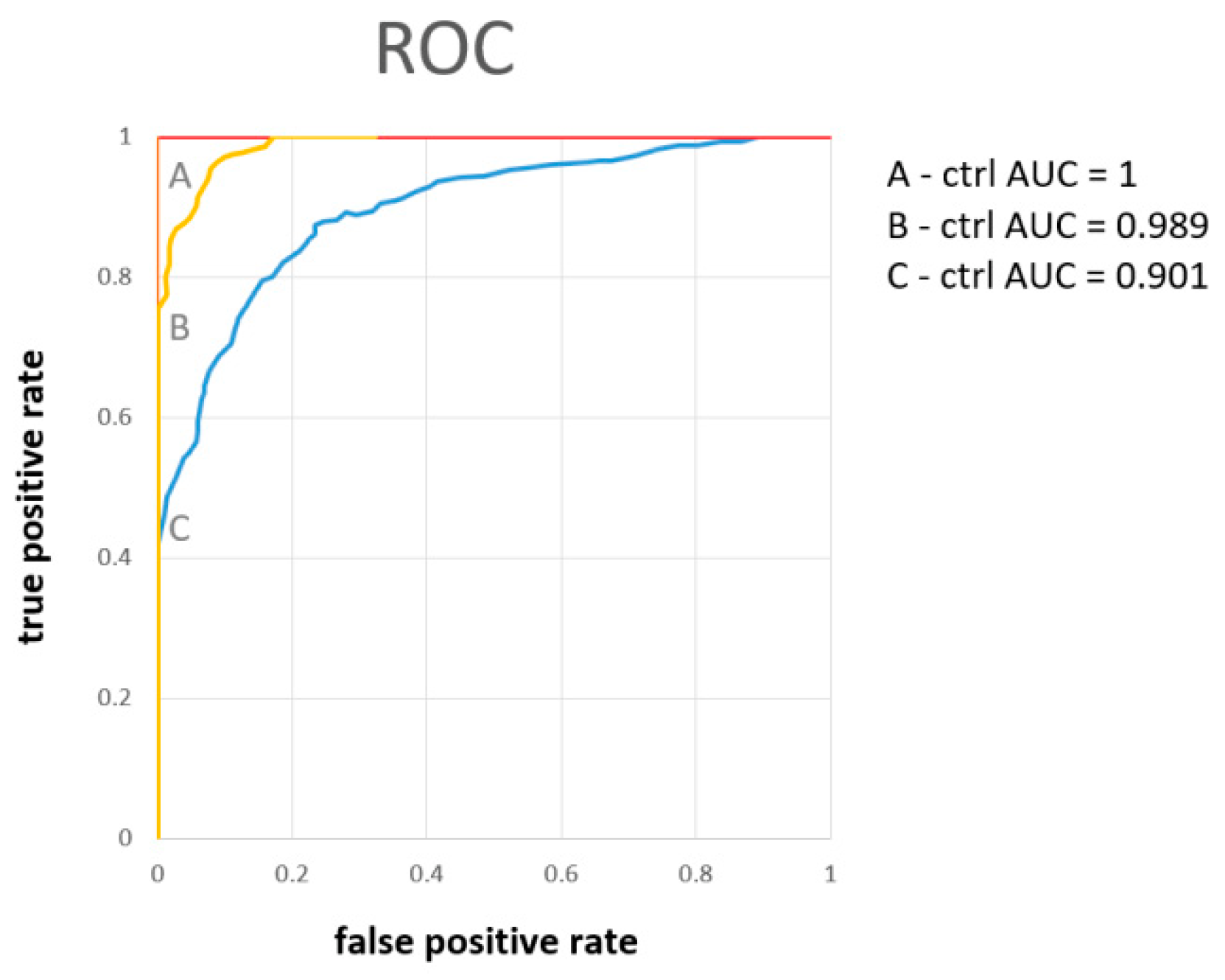
| AUC | Number of Variables (Metabolites) | Metabolites by Importance | Average Accuracy Based on 100 Cross Validation | Oob Error | |
|---|---|---|---|---|---|
| A-ctrl | 1 | 2 | hippurate, citrate | 0.987 | 0 |
| 1 | 5 | hippurate, citrate, pyruvate, alanine, hypoxantine | 0.987 | 0 | |
| B-ctrl | 0.989 | 2 | citrate, hippurate | 0.931 | 0.09 |
| 0.993 | 5 | citrate, hippurate, carnitine, hypoxantine, pyruvate | 0.936 | 0.09 | |
| C-ctrl | 0.844 | 2 | pyruvate, hippurate | 0.784 | 0.32 |
| 0.874 | 5 | pyruvate, hippurate, citrate, glycine, hypoxantine | 0.796 | 0.32 | |
| 0.901 | all | pyruvate, hippurate, citrate, glycine, hypoxanthine, further metabolites were of comparable importance | 0.775 | 0.28 |
| All | A-B | A-C | A-Ctrl | B-C | B-Ctrl | C-Ctrl | |
|---|---|---|---|---|---|---|---|
| acetone | 0.0012 | 0.65 | 0.00078 (152%) | 0.011 | 0.0031 (121%) | 0.035 (80%) | 0.3 |
| alanine | 0.00033 | 0.00003 (−59%) | 0.0026 (−47%) | 0.001 (−46%) | 0.17 | 0.15 | 0.96 |
| acetate | 0.002 | 0.33 | 0.00037 (140%) | 0.016 | 0.0082 (62%) | 0.15 | 0.21 |
| pyruvate | 1.7 × 10−6 | 0.28 | 0.75 | 0.0000034 (−74%) | 0.45 | 0.00036 (−69%) | 0.000016 (−79%) |
| citrate | 3.58 × 10−10 | 0.25 | 0.00036 (−84%) | 0.00000000057 (−91%) | 0.013 | 0.00000041 (−88%) | 0.019 (−49%) |
| carnitine | 3.20 × 10−7 | 0.22 | 0.001 (282%) | 0.00086 (233%) | 0.0000058 (797%) | 0.0000031 (646%) | 0.87 |
| tyrosine | 0.0077 | 0.58 | 0.25 | 0.011 | 0.086 | 0.0015 (65%) | 0.19 |
| hippurate | 1.90 × 10−10 | 0.22 | 0.00075 (−75%) | 0.00000000031 (−85%) | 0.024 | 0.00000022 (−75%) | 0.0068 (−43%) |
| hypoxanthine | 2.40 × 10−5 | 0.021 | 0.41 | 0.018 | 0.0017 (130%) | 0.0000014 (282%) | 0.13 |
| formate | 0.0028 | 0.00042 (−64%) | 0.16 | 0.37 | 0.033 | 0.0034 (139%) | 0.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosolanka, R.; Liptak, P.; Baranovicova, E.; Bobcakova, A.; Vysehradsky, R.; Duricek, M.; Kapinova, A.; Dvorska, D.; Dankova, Z.; Simekova, K.; et al. Changes in the Urine Metabolomic Profile in Patients Recovering from Severe COVID-19. Metabolites 2023, 13, 364. https://doi.org/10.3390/metabo13030364
Rosolanka R, Liptak P, Baranovicova E, Bobcakova A, Vysehradsky R, Duricek M, Kapinova A, Dvorska D, Dankova Z, Simekova K, et al. Changes in the Urine Metabolomic Profile in Patients Recovering from Severe COVID-19. Metabolites. 2023; 13(3):364. https://doi.org/10.3390/metabo13030364
Chicago/Turabian StyleRosolanka, Robert, Peter Liptak, Eva Baranovicova, Anna Bobcakova, Robert Vysehradsky, Martin Duricek, Andrea Kapinova, Dana Dvorska, Zuzana Dankova, Katarina Simekova, and et al. 2023. "Changes in the Urine Metabolomic Profile in Patients Recovering from Severe COVID-19" Metabolites 13, no. 3: 364. https://doi.org/10.3390/metabo13030364
APA StyleRosolanka, R., Liptak, P., Baranovicova, E., Bobcakova, A., Vysehradsky, R., Duricek, M., Kapinova, A., Dvorska, D., Dankova, Z., Simekova, K., Lehotsky, J., Halasova, E., & Banovcin, P. (2023). Changes in the Urine Metabolomic Profile in Patients Recovering from Severe COVID-19. Metabolites, 13(3), 364. https://doi.org/10.3390/metabo13030364