
Adult Triploid Rainbow Trout Can Adapt to Various Dietary Lipid Levels by Coordinating Metabolism in Different Tissues

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Feed, Feeding, and Sampling
2.2. Chemical Analysis
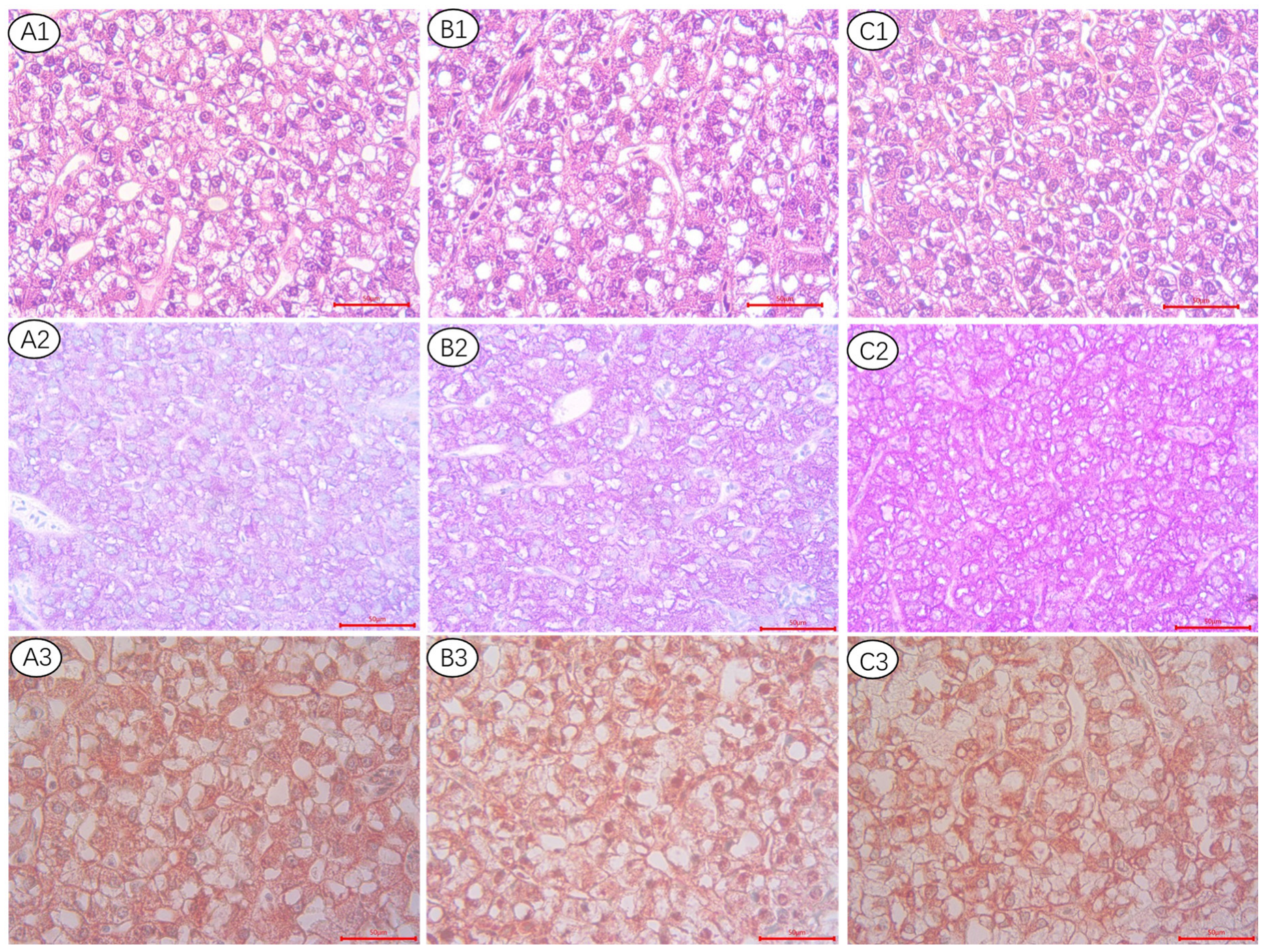
2.3. Hepatic Morphology Analysis
2.4. qRT-PCR Analysis of Lipid-Related Genes
2.5. Calculations
2.6. Statistical Analysis
3. Results
3.1. Growth Performance, Organ Indexes, and Plasma Biochemistry
3.2. Lipid Deposition
3.3. The Histological Appearance of Liver
3.4. The Relative mRNA Expression Levels of Hepatic Metabolism Genes
3.5. The Relative mRNA Expression Levels of Abdominal Adipose Tissue Metabolism Genes
3.6. The Relative mRNA Expression Levels of Muscle Metabolism Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ojolick, E.; Cusack, R.; Benfey, T.; Kerr, S. Survival and growth of all-female diploid and triploid rainbow trout (Oncorhynchus mykiss) reared at chronic high temperature. Aquaculture 1995, 131, 177–187. [Google Scholar] [CrossRef]
- Meng, Y.; Qian, K.; Ma, R.; Liu, X.; Han, B.; Wu, J.; Zhang, L.; Zhan, T.; Hu, X.; Tian, H.; et al. Effects of dietary lipid levels on sub-adult triploid rainbow trout (Oncorhynchus mykiss): 1. Growth performance, digestive ability, health status and expression of growth-related genes. Aquaculture 2019, 513, 734394. [Google Scholar] [CrossRef]
- Meng, Y.; Tian, H.; Hu, X.; Han, B.; Li, X.; Cangzhong, L.; Ma, R.; Kubilay, A. Effects of Dietary Lipid Levels on the Lipid Deposition and Metabolism of Subadult Triploid Rainbow Trout (Oncorhynchus mykiss). Aquacult. Nutr. 2022, 2022, 6924835. [Google Scholar] [CrossRef]
- He, A.Y.; Ning, L.J.; Chen, L.Q.; Chen, Y.L.; Xing, Q.; Li, J.M.; Qiao, F.; Li, D.L.; Zhang, M.L.; Du, Z.Y. Systemic adaptation of lipid metabolism in response to low-and high-fat diet in Nile tilapia (Oreochromis niloticus). Physiol. Rep. 2015, 3, e12485. [Google Scholar] [CrossRef] [PubMed]
- Gélineau, A.; Corraze, G.; Boujard, T.; Larroquet, L.; Kaushik, S. Relation between dietary lipid level and voluntary feed intake, growth, nutrient gain, lipid deposition and hepatic lipogenesis in rainbow trout. Reprod. Nutr. Dev. 2001, 41, 487–503. [Google Scholar] [CrossRef]
- Johnsen, M.H.F. High-energy/low-protein diets for Atlantic salmon: Effects on growth, nutrient retention and slaughter quality. Aquaculture 1994, 124, 109–116. [Google Scholar] [CrossRef]
- Phan, L.T.T.; Kals, J.; Masagounder, K.; Mas-Munoz, J.; La, N.T.H.; Schrama, J.W. Effect of dietary carbohydrate and fat supplementation on the yield and chemical composition of fillet and the location of fat deposition in striped catfish (Pangasius hypophthalmus), African catfish (Clarias gariepinus) and snakehead (Channa striata). Aquac. Rep. 2021, 21, 100806. [Google Scholar] [CrossRef]
- Grahn, T.H.; Zhang, Y.; Lee, M.J.; Sommer, A.G.; Mostoslavsky, G.; Fried, S.K.; Greenberg, A.S.; Puri, V. FSP27 and PLIN1 interaction promotes the formation of large lipid droplets in human adipocytes. Biochem. Biophys. Res. Commun. 2013, 432, 296–301. [Google Scholar] [CrossRef]
- Chen, Q.L.; Gong, Y.; Luo, Z.; Zheng, J.L.; Zhu, Q.L. Differential effect of waterborne cadmium exposure on lipid metabolism in liver and muscle of yellow catfish Pelteobagrus fulvidraco. Aquat. Toxicol. 2013, 142–143, 380–386. [Google Scholar] [CrossRef]
- Albalat, A.; Saera-Vila, A.; Capilla, E.; Gutierrez, J.; Perez-Sanchez, J.; Navarro, I. Insulin regulation of lipoprotein lipase (LPL) activity and expression in gilthead sea bream (Sparus aurata). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 148, 151–159. [Google Scholar] [CrossRef]
- Zhao, M.; Shen, X.; Li, X.; Chen, B.; Fan, L.; Xia, Q.; Zhao, L. Chitooligosaccharide supplementation prevents the development of high fat diet-induced non-alcoholic fatty liver disease (NAFLD) in mice via the inhibition of cluster of differentiation 36 (CD36). J. Funct. Foods 2019, 57, 7–18. [Google Scholar] [CrossRef]
- Islam, A.; Kagawa, Y.; Sharifi, K.; Ebrahimi, M.; Miyazaki, H.; Yasumoto, Y.; Kawamura, S.; Yamamoto, Y.; Sakaguti, S.; Sawada, T.; et al. Fatty Acid Binding Protein 3 Is Involved in n-3 and n-6 PUFA transport in mouse trophoblasts. J. Nutr. 2014, 144, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Kerner, J.; Hoppel, C. Fatty acid import into mitochondria. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1486, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef]
- Meyers, A.; Weiskittel, T.M.; Dalhaimer, P. Lipid droplets: Formation to breakdown. Lipids 2017, 52, 465–475. [Google Scholar] [CrossRef]
- Thiam, A.R.; Beller, M. The why, when and how of lipid droplet diversity. J. Cell Sci. 2017, 130, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Sun, J.; Bian, C.; Ji, S.; Ji, H. Perilipin 1-3 in grass carp Ctenopharyngodon idella: Molecular characterization, gene structure, tissue distribution, and mRNA expression in DHA-induced lipid droplet formation in adipocytes. Fish Physiol. Biochem. 2020, 46, 2311–2322. [Google Scholar] [CrossRef]
- Sztalryd, C.; Brasaemle, D.L. The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1221–1232. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, J.; Zhang, Q. Expression pattern and functions of autophagy-related gene atg5 in zebrafish organogenesis. Autophagy 2011, 7, 1514–1527. [Google Scholar] [CrossRef]
- Scorletti, E.; Carr, R.M. A new perspective on NAFLD: Focusing on lipid droplets. J. Hepatol. 2022, 76, 934–945. [Google Scholar] [CrossRef]
- Meng, Y.; Han, B.; Li, C.; Qian, K.; Liu, X.; Hu, X.; Yang, X.; Tian, H.; Ma, R. Digestive characteristics and blood chemistry profile of triploid rainbow trout Oncorhynchus mykiss: Influence of body size and seasonal variation. Fish. Sci. 2019, 85, 1001–1010. [Google Scholar] [CrossRef]
- Ma, R.; Liu, X.; Meng, Y.; Wu, J.; Zhang, L.; Han, B.; Qian, K.; Luo, Z.; Wei, Y.; Li, C. Protein nutrition on sub-adult triploid rainbow trout (1): Dietary requirement and effect on anti-oxidative capacity, protein digestion and absorption. Aquaculture 2019, 507, 428–434. [Google Scholar] [CrossRef]
- Chen, Q.-L.; Luo, Z.; Pan, Y.-X.; Zheng, J.-L.; Zhu, Q.-L.; Sun, L.-D.; Zhuo, M.-Q.; Hu, W. Differential induction of enzymes and genes involved in lipid metabolism in liver and visceral adipose tissue of juvenile yellow catfish Pelteobagrus fulvidraco exposed to copper. Aquat. Toxicol. 2013, 136, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Nanton, D.A.; Vegusdal, A.; Rørå, A.M.B.; Ruyter, B.; Baeverfjord, G.; Torstensen, B.E. Muscle lipid storage pattern, composition, and adipocyte distribution in different parts of Atlantic salmon (Salmo salar) fed fish oil and vegetable oil. Aquaculture 2007, 265, 230–243. [Google Scholar] [CrossRef]
- Sheridan, M.A.; Kao, Y.-H. Regulation of metamorphosis-associated changes in the lipid metabolism of selected vertebrates. Am. Zool. 1998, 38, 350–368. [Google Scholar] [CrossRef]
- Bjerkeng, B.; Refstie, S.; Fjalestad, K.; Storebakken, T.; Rødbotten, M.; Roem, A. Quality parameters of the flesh of Atlantic salmon (Salmo salar) as affected by dietary fat content and full-fat soybean meal as a partial substitute for fish meal in the diet. Aquaculture 1997, 157, 297–309. [Google Scholar] [CrossRef]
- Mattijssen, F.; Georgiadi, A.; Andasarie, T.; Szalowska, E.; Zota, A.; Krones-Herzig, A.; Heier, C.; Ratman, D.; De Bosscher, K.; Qi, L. Hypoxia-inducible lipid droplet-associated (HILPDA) is a novel peroxisome proliferator-activated receptor (PPAR) target involved in hepatic triglyceride secretion. J. Biol. Chem. 2014, 289, 19279–19293. [Google Scholar] [CrossRef]
- McPherson, P.A.C.; McEneny, J. The biochemistry of ketogenesis and its role in weight management, neurological disease and oxidative stress. J. Physiol. Biochem. 2012, 68, 141–151. [Google Scholar] [CrossRef]
- Sun, Z.; Lazar, M.A. Dissociating fatty liver and diabetes. Trends Endocrinol. Metab. 2013, 24, 4–12. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef]
- Espinosa, A.; Ross, A.; Dovale-Rosabal, G.; Pino-de la Fuente, F.; Uribe-Oporto, E.; Sacristán, C.; Ruiz, P.; Valenzuela, R.; Romero, N.; Aubourg, S.P. EPA/DHA concentrate by urea complexation decreases hyperinsulinemia and increases Plin5 in the liver of mice fed a high-fat diet. Molecules 2020, 25, 3289. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.R.; Shew, T.M.; Schwartz, D.M.; Tzekov, A.; Lepus, C.M.; Abumrad, N.A.; Wolins, N.E. Diacylglycerol enrichment of endoplasmic reticulum or lipid droplets recruits perilipin 3/TIP47 during lipid storage and mobilization. J. Biol. Chem. 2009, 284, 30941–30948. [Google Scholar] [CrossRef] [PubMed]
- Wolins, N.E.; Quaynor, B.K.; Skinner, J.R.; Schoenfish, M.J.; Tzekov, A.; Bickel, P.E. S3-12, Adipophilin, and TIP47 package lipid in adipocytes. J. Biol. Chem. 2005, 280, 19146–19155. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Cuervo, A.M. Autophagy in the cellular energetic balance. Cell Metab. 2011, 13, 495–504. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Tian, J.; Wu, F.; Yang, C.-G.; Jiang, M.; Liu, W.; Wen, H. Dietary lipid levels impact lipoprotein lipase, hormone-sensitive lipase, and fatty acid synthetase gene expression in three tissues of adult GIFT strain of Nile tilapia, Oreochromis niloticus. Fish Physiol. Biochem. 2015, 41, 1–18. [Google Scholar] [CrossRef]
- Zhang, W.; Dan, Z.; Zhuang, Y.; Zheng, J.; Gong, Y.; Liu, Y.; Mai, K.; Ai, Q.; Leng, X. Effects of Dietary Lipid Levels on Growth, Digestive Enzyme Activities, Antioxidant Capacity, and Lipid Metabolism in Turbot (Scophthalmus maximus L.) at Three Different Stages. Aquacult. Nutr. 2022, 2022, 1042263. [Google Scholar] [CrossRef]
- Weisgraber, K.H. Apolipoprotein E: Structure-function relationships. Adv. Protein Chem. 1994, 45, 249–302. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef]
- Kleiner, S.; Nguyen-Tran, V.; Baré, O.; Huang, X.; Spiegelman, B.; Wu, Z. PPARδ agonism activates fatty acid oxidation via PGC-1α but does not increase mitochondrial gene expression and function. J. Biol. Chem. 2009, 284, 18624–18633. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Murray, A.J.; Menassa, D.; Ashmore, T.; Nicholls, A.W.; Griffin, J.L. The contrasting roles of PPARδ and PPARγ in regulating the metabolic switch between oxidation and storage of fats in white adipose tissue. Genome Biol. 2011, 12, R75. [Google Scholar] [CrossRef] [PubMed]
- Bou, M.; Montfort, J.; Le Cam, A.; Rallière, C.; Lebret, V.; Gabillard, J.-C.; Weil, C.; Gutiérrez, J.; Rescan, P.-Y.; Capilla, E. Gene expression profile during proliferation and differentiation of rainbow trout adipocyte precursor cells. BMC Genom. 2017, 18, 347. [Google Scholar] [CrossRef] [PubMed]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Dong, G.-F.; Zou, Q.; Wang, H.; Huang, F.; Liu, X.-C.; Chen, L.; Yang, C.-Y.; Yang, Y.-O. Conjugated linoleic acid differentially modulates growth, tissue lipid deposition, and gene expression involved in the lipid metabolism of grass carp. Aquaculture 2014, 432, 181–191. [Google Scholar] [CrossRef]
- Turchini, G.M.; Ng, W.-K.; Tocher, D.R. Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Tai, C.C.; Ding, S.T. N-3 polyunsaturated fatty acids regulate lipid metabolism through several inflammation mediators: Mechanisms and implications for obesity prevention. J. Nutr. Biochem. 2010, 21, 357–363. [Google Scholar] [CrossRef]
- Huang, D.; Liang, H.; Ren, M.; Ge, X.; Ji, K.; Yu, H.; Maulu, S. Effects of dietary lysine levels on growth performance, whole body composition and gene expression related to glycometabolism and lipid metabolism in grass carp, Ctenopharyngodon idellus fry. Aquaculture 2021, 530, 735806. [Google Scholar] [CrossRef]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Diets1 (20%) | Diets2 (25%) | Diets3 (30%) |
|---|---|---|---|
| Brown fish meal 1 | 51 | 51 | 51 |
| Wheat meal 1 | 15 | 15 | 15 |
| Cassava starch 1 | 5 | 5 | 5 |
| Cellulose | 11.36 | 6.36 | 1.36 |
| Fish oil | 11.8 | 16.8 | 21.8 |
| Soybean oil | 3 | 3 | 3 |
| Vitamin-mineral premix 2 | 1 | 1 | 1 |
| Ca (H2PO4)2 | 0.8 | 0.8 | 0.8 |
| Choline chloride | 0.3 | 0.3 | 0.3 |
| Calcium propionate | 0.1 | 0.1 | 0.1 |
| Ethoxyquin | 0.05 | 0.05 | 0.05 |
| Betaine | 0.5 | 0.5 | 0.5 |
| Y2O3 | 0.05 | 0.05 | 0.05 |
| Astaxanthin 3 | 0.04 | 0.04 | 0.04 |
| Proximate analysis (n = 3, % dry matter) | |||
| Crude protein | 40.6 | 39.7 | 39.9 |
| Crude lipid | 19.8 | 26.1 | 30.5 |
| Gross energy (KJ/g) | 16.3 | 18.5 | 20.2 |
| P/E ratio (mg protein/KJ) | 24.9 | 21.4 | 19.7 |
| Gene Symbol | Gene Name | Primer Sequence (5′-3′) | Genbank Accession No. | |
|---|---|---|---|---|
| Reference gene | ||||
| β-actin | β-actin | F:TACAACGAGCTGAGGGTGGC | R: GGCAGGGGTGTTGAAGGTCT | AJ438158.1 |
| Lipid uptake | ||||
| lpl | Lipoprotein lipase | F:GGACGTTGGGGAGCTGCTTA | R: ATTGAGTCTCCCCGGCCTTG | NM_001124604.1 |
| cd36 | Cluster of differentiation 36 | F:GTCGTGGCTCAAGTCTTCCA | R: TCAAATACTCGGCTCGCCTC | AY606034.1 |
| fatp1 | Fatty acid transport protein 1 | F:GTCCCGTGTTCCTACGCATCT | R:GCCTCATAACGCCCTGCTCT | XM_036941441.1 |
| fatp5 | Fatty acid transport protein 5 | F:TGGGAAGACTTTTGATGGGC | R:CTGGATACGGATGAAATGAGGT | AF072760.1 |
| fabp3 | Fatty acid binding protein 3 | F:ATGGCTGAGGCATTCG | R:CATCTTACCACCGTCTATC | NM_001124713.1 |
| fabp4 | Fatty acid binding protein 4 | F:CGTTGGAACTTGGAAGATGACT | R:TGCCGAGTAGCAAAACCCA | XM 021584874.2 |
| Triacylglycerol synthesis and catabolism | ||||
| accα | Acetyl-CoA carboxylase α | F:CCCATTCGCCTTTTCCTTAC | R:ACCCTGCGTGTCCCCATA | XM_021623129.1 |
| fas | Fatty acid synthase | F:TCTAGAGACGCCACCTTCGA | R:TGCAGTTTCTCCTCAGCCAG | XM_021581290.1 |
| cpt1 | Carnitine palmitoyl transferase 1 | F:TACAGCTGGCCCAATTCAGG | R:TCGCAGTGTTCTTGTCCTCC | AF327058.3 |
| aco | Acetyl-CoA oxidase | F:TTGGGCCTCATCATTGCAGT | R:ACTGGGTCTGGTGCTCAATG | XM_021613038.1 |
| Very low-density lipoprotein protein | ||||
| apoE | apolipoprotein E | F:ACCTGGGAGAGCTCAAGACT | R:CACTGTGTTGCGGATCTCCT | NM_001124346.1 |
| apoB100 | Apolipoprotein B100 | F:GTCTGCCACCATGTTCTCCA | R:CTGGATGGCCTGCTCAAGAA | XM_ 021611526. 1 |
| Transcription factors | ||||
| pparα | Peroxisome proliferator-activated receptor α | F:CACTCCACCCTTCGTCATCC | R:CCTCAGCCTCCTTCTCAAGC | HM536190.1 |
| pparβ | Peroxisome proliferator-activated receptor β | F:GGGGTACACGTGTTCTACCG | R:GTACTTCAGCAGCGTCACCT | HM536191.1 |
| pparγ | Peroxisome proliferator-activated Receptor γ | F:GAGCTGGACATGAACGACCA | R:TGTGCCGTCCTTGTTCATGA | NM_001197212.1 |
| srebp1 | Sterol regulatory element-binding protein 1 | F:TCCTCTCCCTCAATCCCCTG | R:CGAGTCAGCTGCGTTGTCT | KP342261.1 |
| lipid droplets metabolism | ||||
| plin1 | Perilipin1 | F:AAGGTCAGGAACTGGTCACAC | R:TCTGAGGACTGTGCTGTTGTC | XM_021598341.2 |
| plin3 | Perilipin3 | F:GAGAAGGGAGAGGACCTGGA | R:CCTGGGAGACCCTGTACACT | XM_021603908.2 |
| atg7 | autophagy-related gene 7 | F:TACTGAAGGAGGTTATGCG | R:TGATCTGATGAGGGACGAG | XM_036984267.1 |
| atg12 | autophagy-related gene 12 | F:TGGAGGCCAATGAACAGCTG | R:CTTCCCATCGCTGCCAAAAC | XM_021623074.1 |
| Carbohydrate metabolism | ||||
| gk | Glucokinase | F:AGATCACTGTGGGCATCGAC | R:GATGTCACAGTGAGGCGTCA | AF053331.2 |
| pk | Pyruvate kinase | F:GTTCCCTGTCGAGTCTGTGG | R:CAGACGACGAAGCTCCTCAA | XM_021622264.1 |
| g6pase | Glucose-6-phosphatase | F:GCTGACCTGCATACCACCTT | R:CAGCCACCCAGATGAGCTTT | XM_021575943.1 |
| pepck | Phosphoenol pyruvate carboxykinase | F:CGGTGTGTTTGTAGGAGCCT | R:ACGTGGAAGATCTTGGGCAG | NM_001124275.1 |
| Dietary Lipid Levels (% Dry Matter) | F | p | |||
|---|---|---|---|---|---|
| 20 | 25 | 30 | |||
| Growth performance | |||||
| SGR 2 | 0.37 ± 0.01 a | 0.42 ± 0.02 ab | 0.43 ± 0.01 b | 6.91 | 0.03 |
| Organ indexes | |||||
| Condition factor 3 | 2.15 ± 0.04 | 2.19 ± 0.04 | 2.12 ± 0.03 | 0.93 | 0.42 |
| Viscerosomatic index (%) 4 | 13.27 ± 0.13 a | 13.91 ± 0.76 ab | 15.21 ± 0.16 b | 4.75 | 0.03 |
| Hepatosomatic index (%) 5 | 1.25 ± 0.02 | 1.33 ± 0.05 | 1.32 ± 0.03 | 1.26 | 0.31 |
| Liver color | 1.78 ± 0.28 | 1.89 ± 0.26 | 2 ± 0.24 | 0.19 | 0.83 |
| Glycogen (mg/g liver) | 18.95 ± 0.51 a | 22.89 ± 0.90 b | 29.50 ± 0.23 c | 76.13 | 0.00 |
| Lipid content | |||||
| whole fish | |||||
| (g/100g, wet weight) | 20.35 ± 0.39 a | 25.61 ± 0.38 b | 26.32 ± 1.24 b | 17.41 | 0.00 |
| Adipose tissue | |||||
| (g/100g, wet weight) | 74.48 ± 1.26 a | 78.75 ± 0.71 ab | 81.59 ± 1.54 b | 8.59 | 0.02 |
| (g/tissue of a 100 g fish) | 6.57 ± 0.51 a | 6.59 ± 0.32 a | 8.07 ± 0.22 b | 6.70 | 0.02 |
| liver | |||||
| (g/100g, wet weight) | 9.06 ± 1.01 | 8.65 ± 0.59 | 7.49 ± 0.24 | 1.41 | 0.29 |
| (g/tissue of a 100 g fish) | 0.13 ± 0.02 | 0.11 ± 0.01 | 0.1 ± 0.01 | 0.88 | 0.43 |
| Muscle | |||||
| (g/100g, wet weight) | 12.27 ± 0.39 a | 12.82 ± 0.17 ab | 13.85 ± 0.11 b | 10.09 | 0.01 |
| (g/tissue of a 100 g fish) | 7.8 ± 0.25 a | 8.12 ± 0.1 ab | 8.59 ± 0.07 b | 6.11 | 0.04 |
| Dietary Lipid Levels (% Dry Matter) | F | p | |||
|---|---|---|---|---|---|
| 20 | 25 | 30 | |||
| Plasma biochemistry parameters | |||||
| NEFA (umol/L) 2 | 418.27 ± 12.89 ab | 387.83 ± 12.45 a | 482.12 ± 19.73 b | 9.78 | 0.01 |
| Glucose (mg/dL) | 19.82 ± 0 a | 36.94 ± 0.9 ab | 46.4 ± 6.76 b | 11.72 | 0.04 |
| Total cholesterol (mmol/L) | 0.23 ± 0 a | 0.35 ± 0 b | 0.26 ± 0.03 a | 16.02 | 0.03 |
| Triglyceride (mg/dL) | 225.71 ± 17.5 | 262.26 ± 10.63 | 253.84 ± 9.3 | 2.17 | 0.26 |
| HDL-C (mg/dL) 3 | 82.37 ± 1.55 a | 140.18 ± 14.11 b | 97.45 ± 8.12 ab | 10.08 | 0.05 |
| LDL-C (mg/dL) 4 | 112.72 ± 0.19 a | 215.2 ± 4.45 b | 150.43 ± 18.17 a | 23.02 | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Chen, L.; Tian, H.; Sun, G.; Wei, F.; Meng, Y.; Ma, R. Adult Triploid Rainbow Trout Can Adapt to Various Dietary Lipid Levels by Coordinating Metabolism in Different Tissues. Metabolites 2023, 13, 396. https://doi.org/10.3390/metabo13030396
Liu G, Chen L, Tian H, Sun G, Wei F, Meng Y, Ma R. Adult Triploid Rainbow Trout Can Adapt to Various Dietary Lipid Levels by Coordinating Metabolism in Different Tissues. Metabolites. 2023; 13(3):396. https://doi.org/10.3390/metabo13030396
Chicago/Turabian StyleLiu, Gege, Lixia Chen, Haining Tian, Guoliang Sun, Fulei Wei, Yuqiong Meng, and Rui Ma. 2023. "Adult Triploid Rainbow Trout Can Adapt to Various Dietary Lipid Levels by Coordinating Metabolism in Different Tissues" Metabolites 13, no. 3: 396. https://doi.org/10.3390/metabo13030396
APA StyleLiu, G., Chen, L., Tian, H., Sun, G., Wei, F., Meng, Y., & Ma, R. (2023). Adult Triploid Rainbow Trout Can Adapt to Various Dietary Lipid Levels by Coordinating Metabolism in Different Tissues. Metabolites, 13(3), 396. https://doi.org/10.3390/metabo13030396