
A Comprehensive Review on the Roles of Metals Mediating Insect–Microbial Pathogen Interactions



Abstract
:1. Introduction
2. Role of Metals in Insects
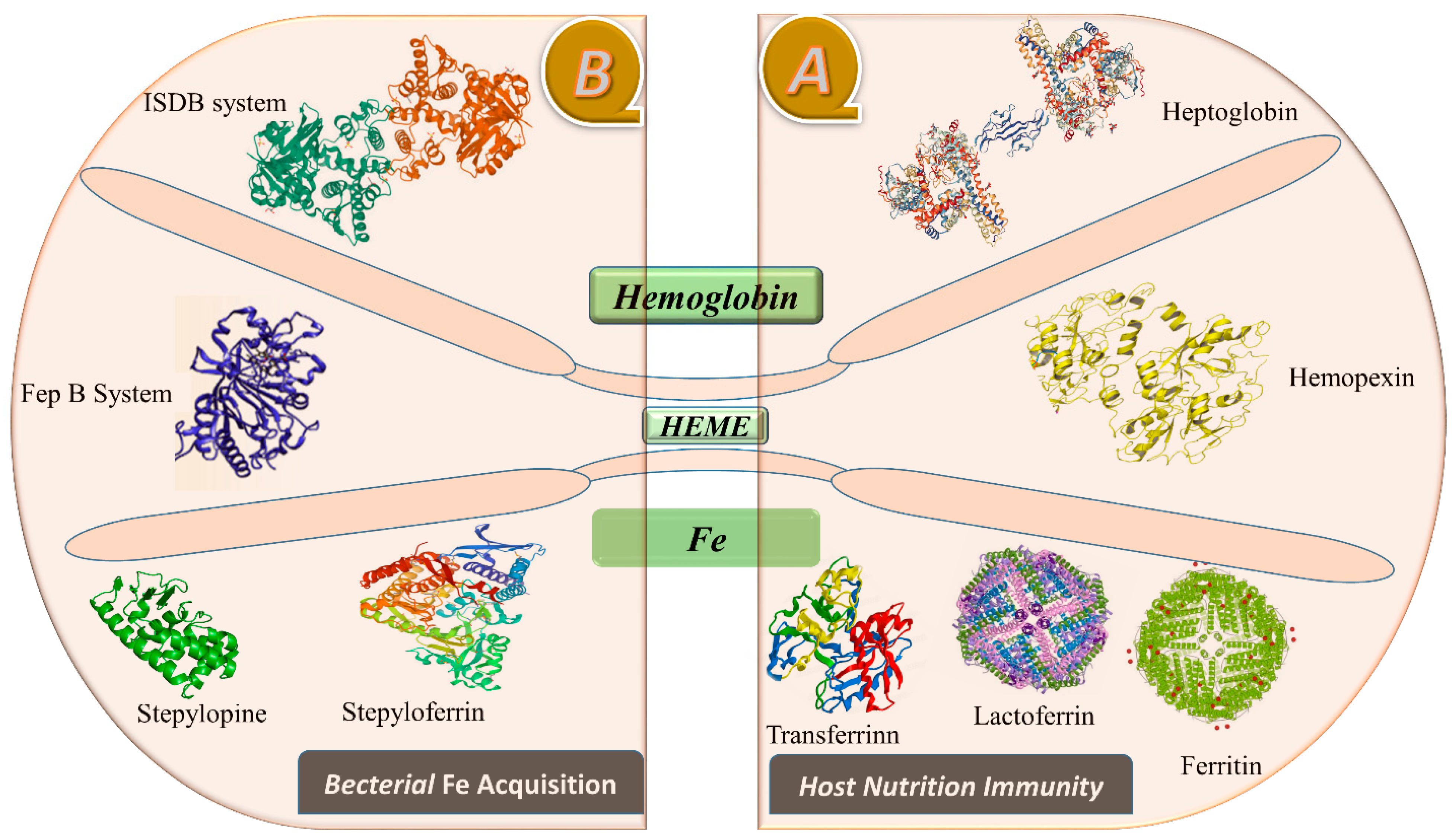
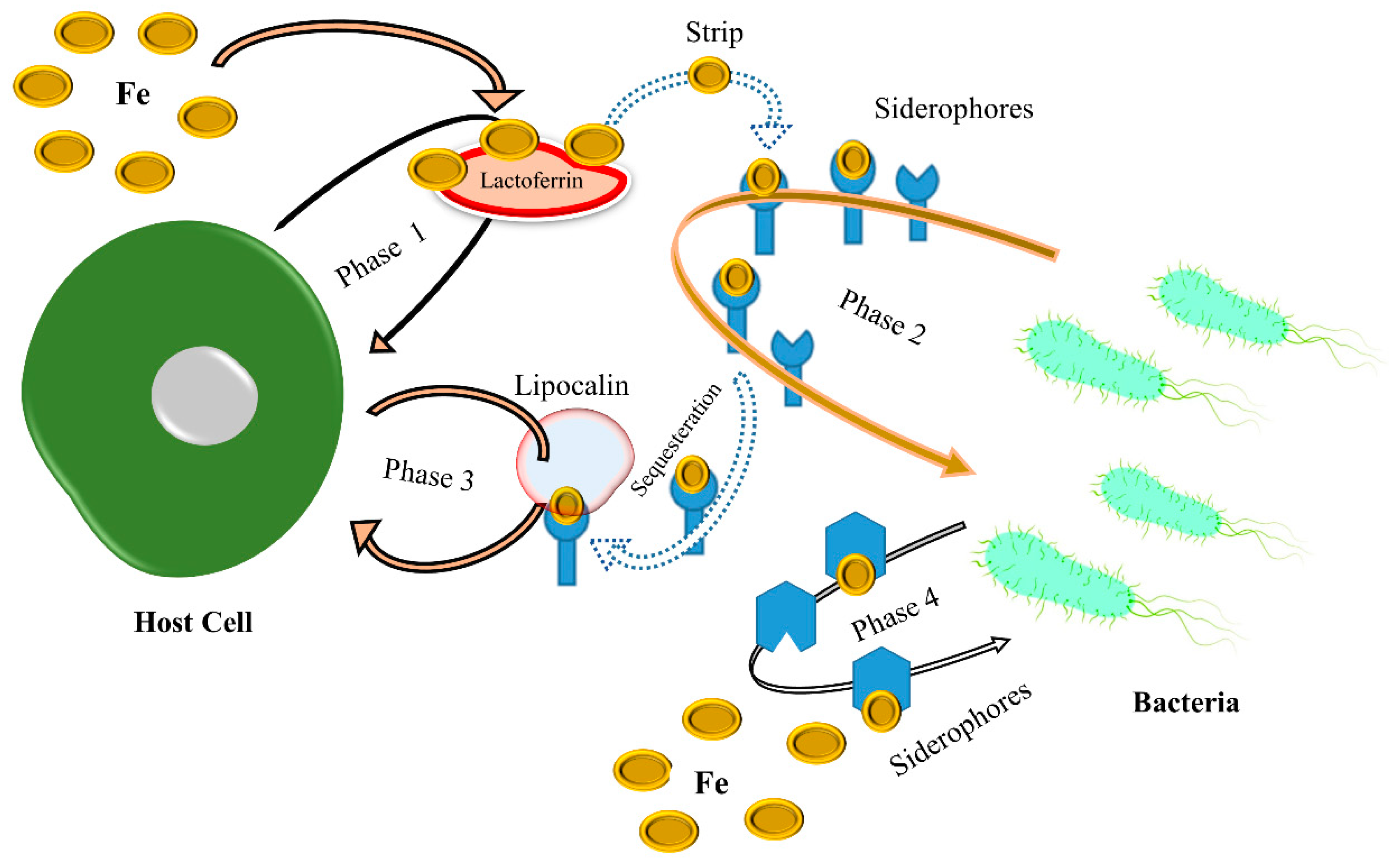
2.1. Iron (Fe)
2.2. Interaction of Plants with Insects and Microbes via Fe
2.3. Zinc (Zn) and Copper (Cu)
2.4. Interaction of Plants with Insects and Microbes through Zn and Cu Metals
2.5. Metals Other Than Fe, Cu, and Zn
3. Role of Metals in Microbial Physiology and Virulence
4. Interactions between Metals and Insects/Microbial Pathogens
5. Mechanisms of Competition between Insects and Microbial Pathogens for Essential Metals
6. Metals That Limit Microbial Growth and Virulence
7. Microorganisms Require Trace Metals for Their Pathogenicity Enhancements
8. Pathogens Use Metals to Evade the Host’s Immune Response
9. Implications and Applications
9.1. Development of Novel Insecticides and Antimicrobial Therapies
9.2. Promoting Agricultural Sustainability
9.3. Potential Applications for Pest and Pathogen Control
10. Critical Observation and Further Research
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, L.; Su, Q.F.; Wang, L.S.; Lv, M.W.; Hou, Y.X.; Li, S.S. Linalool: A ubiquitous floral volatile mediating the communication between plants and insects. J. Syst. Evol. 2022, 61, 538–549. [Google Scholar] [CrossRef]
- Leitão, J.H. Microbial Virulence Factors. IJMS 2020, 21, 5320. [Google Scholar] [CrossRef] [PubMed]
- Biere, A.; Bennett, A.E. Three-way interactions between plants, microbes and insects. Functional 2013, 27, 567–573. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Chen, X.; Xu, C.; Zhao, H.; Zhang, X.; Zeng, G.; Qian, Y.; Liu, R.; Guo, N.; Mi, W. Horizontal gene transfer allowed the emergence of broad host range entomopathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 7982–7989. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Guo, L.; Bai, Y.; Kang, S.; Sun, D.; Qin, J.; Ye, F.; Wang, S.; Wu, Q.; Xie, W. Retrotransposon-mediated evolutionary rewiring of a pathogen response orchestrates a resistance phenotype in an insect host. Proc. Natl. Acad. Sci. USA 2023, 120, e2300439120. [Google Scholar] [CrossRef]
- Anand, U.; Pal, T.; Yadav, N.; Singh, V.K.; Tripathi, V.; Choudhary, K.K.; Shukla, A.K.; Sunita, K.; Kumar, A.; Bontempi, E. Current Scenario and Future Prospects of Endophytic Microbes: Promising Candidates for Abiotic and Biotic Stress Management for Agricultural and Environmental Sustainability. Microb. Ecol. 2023, 1–32. [Google Scholar] [CrossRef]
- Perveen, N.; Muhammad, K.; Muzaffar, S.B.; Zaheer, T.; Munawar, N.; Gajic, B.; Sparagano, O.A.; Kishore, U.; Willingham, A.L. Host-pathogen interaction in arthropod vectors: Lessons from viral infections. Front. Immunol. 2023, 14, 1061899. [Google Scholar] [CrossRef]
- Gebre, S.H. Bio-inspired Synthesis of Metal and Metal Oxide Nanoparticles: The Key Role of Phytochemicals. J. Clust. Sci. 2023, 34, 665–704. [Google Scholar] [CrossRef]
- Page, M.G.P. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin. Infect. Dis. 2019, 69 (Suppl. 7), S529–S537. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Li, X.-S.; Ren, K.-D.; Peng, J.; Luo, X.-J. Restoration of metal homeostasis: A potential strategy against neurodegenerative diseases. Ageing Res. Rev. 2023, 87, 101931. [Google Scholar] [CrossRef] [PubMed]
- Dharmaraja, A.T. Role of Reactive Oxygen Species (ROS) in Therapeutics and Drug Resistance in Cancer and Bacteria. J. Med. Chem. 2017, 60, 3221–3240. [Google Scholar] [CrossRef]
- Schrand, A.M.; Rahman, M.F.; Hussain, S.M.; Schlager, J.J.; Smith, D.A.; Syed, A.F. Metal-based nanoparticles and their toxicity assessment. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2010, 2, 544–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.F.; Aarts, M.G. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. CMLS 2012, 69, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- Thallaj, N. Review of a Few Selected Examples of Intermolecular Dioxygenases Involving Molecular Oxygen and Non-Heme Iron Proteins. Int. J. Adv. Parmacutical Sci. Res. (IJAPSR) 2023, 3, 1–18. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. ROLE OF Iron in Plant Growth and Metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Stijlemans, B.; Beschin, A.; Magez, S.; Van Ginderachter, J.A.; De Baetselier, P. Iron homeostasis and Trypanosoma brucei associated immunopathogenicity development: A battle/quest for iron. BioMed Res. Int. 2015, 2015, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Aziz, D.A.A.; Penyelidikan, P.P. The Development and Optimization of Processes for The expression of Sialylated Recombinant Human Therapeutic Glycoprotein in Insect Cell-Baculovirus System; University Teknologi Malaysia: Johor, Malaysia, 2001. [Google Scholar]
- Macaluso, G.; Grippi, F.; Di Bella, S.; Blanda, V.; Gucciardi, F.; Torina, A.; Guercio, A.; Cannella, V. A Review on the Immunological Response against Trypanosoma cruzi. Pathogens 2023, 12, 282. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin and iron regulation, 10 years later. Blood J. Am. Soc. Hematol. 2011, 117, 4425–4433. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, W.E.; Bazydlo, L.A.; Harris, N.S. The molecular biology of human iron metabolism. Lab. Med. 2014, 45, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinowski, D.S.; Richardson, D.R. Cellular and molecular biology of iron-binding proteins. In Cellular and Molecular Biology of Metals; CRC Press: Boca Raton, FL, USA, 2010; pp. 177–190. [Google Scholar]
- Tandara, L.; Salamunić, I. Iron metabolism: Current facts and future directions. Biochem. Medica 2012, 22, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; McCulloh, R.J. Hemopexin and haptoglobin: Allies against heme toxicity from hemoglobin not contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef]
- Payne, S.M. Iron acquisition in microbial pathogenesis. Trends Microbiol. 1993, 1, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.C.; Garcia-Herrero, A.; Johanson, T.H.; Krewulak, K.D.; Lau, C.K.; Peacock, R.S.; Slavinskaya, Z.; Vogel, H.J. Siderophore uptake in bacteria and the battle for iron with the host; a bird’s eye view. Biometals 2010, 23, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; De Bei, O.; Bettati, S.; Campanini, B.; Kovachka, S.; Gianquinto, E.; Spyrakis, F.; Ronda, L. Iron metabolism at the interface between host and pathogen: From nutritional immunity to antibacterial development. Int. J. Mol. Sci. 2020, 21, 2145. [Google Scholar] [CrossRef] [Green Version]
- Barton, L.L. Transmembrane Movement: Mechanisms and Examples. Struct. Funct. Relatsh. Prokaryotes 2005, 21, 468–526. [Google Scholar]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in iron metabolism: From mechanism to therapy potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lu, Y.; Wang, Y.; Zhang, Y.; Xia, B.; Cao, J. Siderophores induce mitophagy-dependent apoptosis in platelets. Ann. Transl. Med. 2020, 8, 14. [Google Scholar] [CrossRef]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol. 2019, 109, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Scott, Z.W.; Choi, S.-r.; Britigan, B.E.; Narayanasamy, P. Development of Gallium (III) as an Antimicrobial Drug Targeting Pathophysiologic Iron Metabolism of Human Pathogens. ACS Infect. Dis. 2023, 9, 716–738. [Google Scholar] [CrossRef] [PubMed]
- Laranjeira-Silva, M.F.; Hamza, I.; Pérez-Victoria, J.M. Iron and heme metabolism at the leishmania–host interface. Trends Parasitol. 2020, 36, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K. Illuminating Siderophore Transporter Functionality with Thiopeptide Antibiotics. mBio 2023, 14, e0332622. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Jiménez, A.; Marcos-Torres, F.J.; Llamas, M.A. Mechanisms of iron homeostasis in Pseudomonas aeruginosa and emerging therapeutics directed to disrupt this vital process. Microb. Biotechnol. 2023, 16, 1475–1491. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Farhana, A.; Ehtesham, N.Z.; Hasnain, S.E. Iron acquisition, assimilation and regulation in mycobacteria. Infect. Genet. Evol. 2011, 11, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Rementeria, A.; López-Molina, N.; Ludwig, A.; Vivanco, A.B.; Bikandi, J.; Pontón, J.; Garaizar, J. Genes and molecules involved in Aspergillus fumigatus virulence. Rev. Iberoam Micol. 2005, 22, 1–23. [Google Scholar] [CrossRef]
- Kita, K.; Nihei, C.; Tomitsuka, E. Parasite Mitochondria as Drug Target: Diversity and Dynamic Changes During the Life Cycle. Curr. Med. Chem. 2003, 10, 2535–2548. [Google Scholar] [CrossRef]
- Huber, D.; Römheld, V.; Weinmann, M. Relationship between nutrition, plant diseases and pests. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 283–298. [Google Scholar]
- Haschka, D.; Hoffmann, A.; Weiss, G. Iron in immune cell function and host defense. In Seminars in Cell & Developmental Biology, 2021; Elsevier: Amsterdam, The Netherlands, 2021; pp. 27–36. [Google Scholar]
- Khan, S.; Subhan, F.; Haleem, K.S.; Khattak, M.N.K.; Khan, I.; Sultan, T.; Tauseef, I. Impact of plant growth-promoting rhizobacteria on yield and disease control of Nicotiana tabacum. Arch. Biol. Sci. 2018, 70, 717–725. [Google Scholar] [CrossRef]
- Iwama, R.E.; Moran, Y. Origins and diversification of animal innate immune responses against viral infections. Nat. Ecol. Evol. 2023, 7, 1–12. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alias-Villegas, C.; Bellogín, R.A.; del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Vidhyasekaran, P. Plant Hormone Signaling Systems in Plant Innate Immunity; Springer: Berlin, Germany, 2015; Volume 2. [Google Scholar]
- García, M.J.; Lucena, C.; Romera, F.J. Ethylene and Nitric Oxide Involvement in the Regulation of Fe and P Deficiency Responses in Dicotyledonous Plants. Int. J. Mol. Sci. 2021, 22, 4904. [Google Scholar] [CrossRef] [PubMed]
- Maurer, F.; Müller, S.; Bauer, P. Suppression of Fe deficiency gene expression by jasmonate. Plant Physiol. Biochem. 2011, 49, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Brumbarova, T.; Bauer, P.; Ivanov, R. Molecular mechanisms governing Arabidopsis iron uptake. Trends Plant Sci. 2015, 20, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Chen, C.-L.; Cui, M.; Zhou, W.-J.; Wu, H.-L.; Ling, H.-Q. Four IVa bHLH Transcription Factors Are Novel Interactors of FIT and Mediate JA Inhibition of Iron Uptake in Arabidopsis. Mol. Plant 2018, 11, 1166–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hell, R.; Stephan, U.W. Iron uptake, trafficking and homeostasis in plants. Planta 2003, 216, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Mateo, P.; Ye, M.; Zhang, X.; Berset, J.D.; Handrick, V.; Radisch, D.; Grabe, V.; Köllner, T.G.; Gershenzon, J.; et al. Plant iron acquisition strategy exploited by an insect herbivore. Science 2018, 361, 694–697. [Google Scholar] [CrossRef] [Green Version]
- Ratledge, C.; Dover, L.G. Iron Metabolism in Pathogenic Bacteria. Annu. Rev. Microbiol. 2000, 54, 881–941. [Google Scholar] [CrossRef]
- Yadav, B.; Dubey, R.; Gnanasekaran, P.; Narayan, O.P. OMICS approaches towards understanding plant’s responses to counterattack heavy metal stress: An insight into molecular mechanisms of plant defense. Plant Gene 2021, 28, 100333. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Heerman, M.C.; Cook, S.C.; Evans, J.D.; DeGrandi-Hoffman, G.; Banmeke, O.; Zhang, Y.; Huang, S.; Hamilton, M.; Chen, Y.P. Transferrin-mediated iron sequestration suggests a novel therapeutic strategy for controlling Nosema disease in the honey bee, Apis mellifera. PLoS Pathog. 2021, 17, e1009270. [Google Scholar] [CrossRef]
- Pahari, A.; Pradhan, A.; Nayak, S.K.; Mishra, B.B. Bacterial Siderophore as a Plant Growth Promoter. Microb. Biotechnol. 2017, 1, 163–180. [Google Scholar] [CrossRef]
- Schmidt, W. Iron solutions: Acquisition strategies and signaling pathways in plants. Trends Plant Sci. 2003, 8, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Keel, C. Signaling in the Rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Rolfe, M.D.; Smith, L.J. Transcriptional regulation of bacterial virulence gene expression by molecular oxygen and nitric oxide. Virulence 2014, 5, 794–809. [Google Scholar] [CrossRef]
- Rodrigues, W.F.C.; Lisboa, A.B.P.; Lima, J.E.; Ricachenevsky, F.K.; Del-Bem, L.E. Ferrous iron uptake via IRT1/ZIP evolved at least twice in green plants. New Phytol. 2023, 237, 1951–1961. [Google Scholar] [CrossRef]
- Gorman, M.J. Iron Homeostasis in Insects. Annu. Rev. Entomol. 2023, 68, 51–67. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, A.; Rehman, H.U.; Usman, M.; Wakeel, A.; Shahid, M.S.; Alam, S.; Sanaullah, M.; Atiq, M.; Farooq, M. Nanobiotechnology in crop stress management: An overview of novel applications. Discov. Nano 2023, 18, 74. [Google Scholar] [CrossRef]
- Mwangi, M.N.; Oonincx, D.G.; Stouten, T.; Veenenbos, M.; Melse-Boonstra, A.; Dicke, M.; Van Loon, J.J. Insects as sources of iron and zinc in human nutrition. Nutr. Res. Rev. 2018, 31, 248–255. [Google Scholar] [CrossRef]
- Jones, M.W.M.; de Jonge, M.D.; James, S.A.; Burke, R. Elemental mapping of the entire intact Drosophila gastrointestinal tract. JBIC J. Biol. Inorg. Chem. 2015, 20, 979–987. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, J.A.; Schneuwly, S. Copper and Zinc Homeostasis: Lessons from Drosophila melanogaster. Front. Genet. 2017, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Groth, C.; Sasamura, T.; Khanna, M.R.; Whitley, M.; Fortini, M.E. Protein trafficking abnormalities in Drosophila tissues with impaired activity of the ZIP7 zinc transporter Catsup. Development 2013, 140, 3018–3027. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Qin, J.; Zhou, X.; Zhang, Y. Insect Transcription Factors: A Landscape of Their Structures and Biological Functions in Drosophila and beyond. Int. J. Mol. Sci. 2018, 19, 3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2011, 142, 1–16. [Google Scholar] [CrossRef]
- Kanost, M.; Gorman, M. Phenoloxidases in insect immunity. Insect Immunol. 2008, 1, 69–96. [Google Scholar]
- Li, Y.; Wang, Y.; Jiang, H.; Deng, J. Crystal structure of Manduca sexta prophenoloxidase provides insights into the mechanism of type 3 copper enzymes. Proc. Natl. Acad. Sci. USA 2009, 106, 17002–17006. [Google Scholar] [CrossRef]
- Xiao, G. Molecular physiology of zinc in Drosophila melanogaster. Curr. Opin. Insect Sci. 2022, 51, 100899. [Google Scholar] [CrossRef]
- Balamurugan, K.; Egli, D.; Hua, H.; Rajaram, R.; Seisenbacher, G.; Georgiev, O.; Schaffner, W. Copper homeostasis in Drosophila by complex interplay of import, storage and behavioral avoidance. EMBO J. 2007, 26, 1035–1044. [Google Scholar] [CrossRef]
- Selvaraj, A.; Balamurugan, K.; Yepiskoposyan, H.; Zhou, H.; Egli, D.; Georgiev, O.; Thiele, D.J.; Schaffner, W. Metal-responsive transcription factor (MTF-1) handles both extremes, copper load and copper starvation, by activating different genes. Genes Dev. 2005, 19, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Slobodian, M.R.; Petahtegoose, J.D.; Wallis, A.L.; Levesque, D.C.; Merritt, T.J. The effects of essential and non-essential metal toxicity in the Drosophila melanogaster insect model: A review. Toxics 2021, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Bahadorani, S.; Mukai, S.; Egli, D.; Hilliker, A.J. Overexpression of metal-responsive transcription factor (MTF-1) in Drosophila melanogaster ameliorates life-span reductions associated with oxidative stress and metal toxicity. Neurobiol. Aging 2010, 31, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Wintergerst, E.S.; Maggini, S.; Hornig, D.H. Immune-enhancing role of vitamin C and zinc and effect on clinical conditions. Ann. Nutr. Metab. 2006, 50, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the Role of Antimicrobial Peptides in Insects. Int. J. Mol. Sci. 2023, 24, 5753. [Google Scholar] [CrossRef] [PubMed]
- Hrdina, A.; Iatsenko, I. The roles of metals in insect–microbe interactions and immunity. Curr. Opin. Insect Sci. 2022, 49, 71–77. [Google Scholar] [CrossRef] [PubMed]
- McHenry, C.S. DNA replicases from a bacterial perspective. Annu. Rev. Biochem. 2011, 80, 403–436. [Google Scholar] [CrossRef] [Green Version]
- Ishida, T. Antiviral activities of Zn2+ ions for viral prevention, replication, capsid protein in intracellular proliferation of viruses. World Sci. News 2018, 97, 28–50. [Google Scholar]
- Kuznetsova, A.A.; Fedorova, O.S.; Kuznetsov, N.A. Structural and molecular kinetic features of activities of DNA polymerases. Int. J. Mol. Sci. 2022, 23, 6373. [Google Scholar] [CrossRef]
- Lee, J.Y.; Chang, C.; Song, H.K.; Moon, J.; Yang, J.K.; Kim, H.-K.; Kwon, S.-T.; Suh, S.W. Crystal structure of NAD+-dependent DNA ligase: Modular architecture and functional implications. EMBO J. 2000, 19, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- Begg, S.L. The role of metal ions in the virulence and viability of bacterial pathogens. Biochem. Soc. Trans. 2019, 47, 77–87. [Google Scholar] [CrossRef]
- Sharma, K.K.; Singh, D.; Mohite, S.V.; Williamson, P.R.; Kennedy, J.F. Metal manipulators and regulators in human pathogens: A comprehensive review on microbial redox copper metalloenzymes “multicopper oxidases and superoxide dismutases”. Int. J. Biol. Macromol. 2023, 233, 123534. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; de la Lastra, J.M.P.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Superoxide Anion Chemistry—Its Role at the Core of the Innate Immunity. Int. J. Mol. Sci. 2023, 24, 1841. [Google Scholar] [CrossRef] [PubMed]
- Zaminpira, S.; Niknamian, S. How butterfly effect or deterministic chaos theory in theoretical physics explains the main cause of cancer. EC Cancer 2017, 2, 227–238. [Google Scholar]
- Stengel, A. Characterisation of components and mechanisms involved in redox-regulation of protein import into chloroplasts. LMU 2009. [Google Scholar] [CrossRef]
- Isernia, C.; Bucci, E.; Leone, M.; Zaccaro, L.; Di Lello, P.; Digilio, G.; Esposito, S.; Saviano, M.; Di Blasio, B.; Pedone, C. NMR structure of the single QALGGH zinc finger domain from the Arabidopsis thaliana SUPERMAN protein. Chembiochem 2003, 4, 171–180. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, C.; Zeng, J.; Yun, Z.; Liu, Y.; Qu, H.; Jiang, Y.; Duan, X.; Xia, R. Micro RNA 528, a hub regulator modulating ROS homeostasis via targeting of a diverse set of genes encoding copper-containing proteins in monocots. New Phytol. 2020, 225, 385–399. [Google Scholar] [CrossRef]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef]
- Zekri, M.; Obreza, T.A. Plant Nutrients for Citrus Trees; University of Florida Cooperative Extension Service: Panama, FL, USA, 2003. [Google Scholar]
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of heavy metals through terrestrial food webs: A review. Environ. Monit. Assess. 2015, 187, 201. [Google Scholar] [CrossRef] [Green Version]
- Gaspéri, J.; Ayrault, S.; Moreau-Guigon, E.; Alliot, F.; Labadie, P.; Budzinski, H.; Blanchard, M.; Muresan, B.; Caupos, E.; Cladière, M.; et al. Contamination of soils by metals and organic micropollutants: Case study of the Parisian conurbation. Environ. Sci. Pollut. Res. 2018, 25, 23559–23573. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, M. Interactions between phytoplankton and trace metals in the ocean. Advances in Marine Biology. 2001, 41, 1–128. [Google Scholar]
- Aparachita, P. Effects of Water Soluble Zinc on Growth Performance, Immune Response, and Tissue Zinc Transporters in Nursery Pigs; Oklahoma State University: Stillwater, OK, USA, 2020. [Google Scholar]
- Castro, P.H.; Lilay, G.H.; Assunção, A.G. Regulation of micronutrient homeostasis and deficiency response in plants. In Plant Micronutrient Use Efficiency; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–15. [Google Scholar]
- Kuliyev, E. Functional and Biochemical Characterization of ZIP4 Extracellular Domain⎯Implications on Acrodermatitis Enteropathica, a Life-Threatening Genetic Disorder. Michigan State University: East Lansing, MI, USA, 2020. [Google Scholar]
- Murakami, M.; Hirano, T. Intracellular zinc homeostasis and zinc signaling. Cancer Sci. 2008, 99, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- John, E.; Laskow, T.C.; Buchser, W.J.; Pitt, B.R.; Basse, P.H.; Butterfield, L.H.; Kalinski, P.; Lotze, M.T. Zinc in innate and adaptive tumor immunity. J. Transl. Med. 2010, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, M.; Bruno, L.B.; Banu, J.R. Alleviation of environmental stress in plants: The role of beneficial Pseudomonas spp. Crit. Rev. Environ. Sci. Technol. 2017, 47, 372–407. [Google Scholar] [CrossRef]
- Naz, T.; Iqbal, M.M.; Fahad, S.; Akhtar, J.; Saqib, M.; Alamri, S.; Siddiqui, M.H.; Saud, S.; Khattak, J.Z.K.; Ali, S.; et al. Bio-fortification of Two Wheat Cultivars with Iron and Zinc Through Their Soil and Foliar Application in Salt-Factored Soil: Growth, Ionic, Physiological, and Biochemical Modifications. J. Plant Growth Regul. 2023, 1–19. [Google Scholar] [CrossRef]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Curr. Res. Microb. Sci. 2021, 2, 100054. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The impact of microbes in the orchestration of plants’ resistance to biotic stress: A disease management approach. Appl. Microbiol. Biotechnol. 2019, 103, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Kahromi, S.; Khara, J. Chitosan stimulates secondary metabolite production and nutrient uptake in medicinal plantDracocephalum kotschyi. J. Sci. Food Agric. 2021, 101, 3898–3907. [Google Scholar] [CrossRef]
- Khan, T.; Abbasi, B.H.; Khan, M.A. The interplay between light, plant growth regulators and elicitors on growth and secondary metabolism in cell cultures of Fagonia indica. J. Photochem. Photobiol. B Biol. 2018, 185, 153–160. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.; Petropoulos, V.; Molotokaite, E.; Cerullo, G.; Casazza, A.P.; Maiuri, M.; Santabarbara, S. Ultrafast excited-state dynamics in land plants Photosystem I core and whole supercomplex under oxidised electron donor conditions. Photosynth. Res. 2020, 144, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Biological chemistry of superoxide radicals. Chemtexts 2020, 6, 7. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S. Melatonin alleviates cold-induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Song, X.; Zhong, Z.; Gao, L.; Weiss, B.L.; Wang, J. Metabolic interactions between disease-transmitting vectors and their microbiota. Trends Parasitol. 2022, 38, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Soetan, K.; Olaiya, C.; Oyewole, O. The importance of mineral elements for humans, domestic animals and plants: A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Bhatla, S.C.A.; Lal, M.; Kathpalia, R.; Bhatla, S.C. Plant mineral nutrition. Plant Physiol. Dev. Metab. 2018, 37–81. [Google Scholar]
- Jha, C.K.; Saraf, M. Plant growth promoting rhizobacteria (PGPR): A review. J. Agric. Res. Dev. 2015, 5, 108–119. [Google Scholar]
- Rajak, P.; Roy, S.; Ganguly, A.; Mandi, M.; Dutta, A.; Das, K.; Nanda, S.; Sarkar, S.; Khatun, S.; Ghanty, S. Protective potential of vitamin C and E against organophosphate toxicity: Current status and perspective. J. Ecophysiol. Occup. Health 2022, 456, 141–154. [Google Scholar] [CrossRef]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yi, H. Effect of sulfur dioxide on ROS production, gene expression and antioxidant enzyme activity in Arabidopsis plants. Plant Physiol. Biochem. 2012, 58, 46–53. [Google Scholar] [CrossRef]
- Huang, W.; Montroni, D.; Wang, T.; Murata, S.; Arakaki, A.; Nemoto, M.; Kisailus, D. Nanoarchitected Tough Biological Composites from Assembled Chitinous Scaffolds. Acc. Chem. Res. 2022, 55, 1360–1371. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Jiang, L.; Dong, Z. Overflow control for sustainable development by superwetting surface with biomimetic structure. Chem. Rev. 2022, 123, 2276–2310. [Google Scholar] [CrossRef] [PubMed]
- Goetzke, H.H.; Pattrick, J.G.; Federle, W. Froghoppers jump from smooth plant surfaces by piercing them with sharp spines. Proc. Natl. Acad. Sci. USA 2019, 116, 3012–3017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, R. Robin Stewart’s Chemical Free Pest Control: Hundreds of Practical & Inexpensive Ways to Control Pests without Chemicals; Black Inc.: Melbourne Australia, 2002. [Google Scholar]
- Gochfield, M.; Laumbach, R. Chemical hazards. In Occupational and Environemntal Health-Recognizing and Preventing Disease and Injury; Lippincott Williams & Wilkins: Ambler, PA, USA, 2011; Volume 1, pp. 192–226. [Google Scholar]
- Deng, L. Shining the Light: Structure and Function Relationship of Calcium-Regulated Photoproteins; University of Georgia: Athens, GA, USA, 2004. [Google Scholar]
- Waterhouse, D. Studies on the digestion of wool by insects IV. Absorption and elimination of metals by Lepidopterous larvae, with special reference to the clothes moth, Tineola Bisselliella (Humm.). Aust. J. Biol. Sci. 1952, 5, 143–168. [Google Scholar] [CrossRef]
- Mma, A.A.F.; Ahmed, F.; Abada, M.A.M.; Yassen, E. Effect of spraying silicon and selenium on growth, vine nutritional status, berry setting, yield and berries quality of superior grapevines grown under sandy soil conditions i-the effect on growth and vine nutritional status. Fayoum J. Agric. Res. Dev. 2017, 31, 135–144. [Google Scholar]
- Bánfalvi, G. Heavy metals, trace elements and their cellular effects. Cell. Eff. Heavy Met. 2011, 3–28. [Google Scholar]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Wright, J.F. Biological Insights into Therapeutic Protein Modifications throughout Trafficking and Their Biopharmaceutical Applications. Int. J. Cell Biol. 2013, 2013, 273086. [Google Scholar] [CrossRef] [Green Version]
- Moskovitz, J.; Yim, M.B.; Chock, P.B. Free radicals and disease. Arch. Biochem. Biophys. 2002, 397, 354–359. [Google Scholar] [CrossRef]
- Calderon, I.L.; Elías, A.O.; Fuentes, E.L.; Pradenas, G.A.; Castro, M.E.; Arenas, F.A.; Perez, J.M.; Vasquez, C.C. Tellurite-mediated disabling of [4Fe–4S] clusters of Escherichia coli dehydratases. Microbiology 2009, 155, 1840–1846. [Google Scholar] [CrossRef] [Green Version]
- Madkour, L.H. Targeted Drug Delivery. J. Target. Drug Deliv. 2019, 3. [Google Scholar]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Zamudio, R.; Ha, H.C. Environmental epigenetics in metal exposure. Epigenetics 2011, 6, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.Y.; Merzendorfer, H.; Zhang, W.; Zhang, J.; Muthukrishnan, S. Biosynthesis, turnover, and functions of chitin in insects. Annu. Rev. Entomol. 2016, 61, 177–196. [Google Scholar] [CrossRef]
- Tariq, A.; Zeng, F.; Graciano, C.; Ullah, A.; Sadia, S.; Ahmed, Z.; Murtaza, G.; Ismoilov, K.; Zhang, Z. Regulation of Metabolites by Nutrients in Plants. In Plant Ionomics: Sensing, Signaling, and Regulation; Urumqi China Wiley: Hoboken, NJ, USA, 2023; pp. 1–18. [Google Scholar]
- Bretscher, H.; O’connor, M.B. The Role of Muscle in Insect Energy Homeostasis. Front. Physiol. 2020, 11, 580687. [Google Scholar] [CrossRef]
- Mattson, W.J., Jr. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar] [CrossRef]
- Dresen, M.; Valentin-Weigand, P.; Berhanu Weldearegay, Y. Role of Metabolic Adaptation of Streptococcus suis to Host Niches in Bacterial Fitness and Virulence. Pathogens 2023, 12, 541. [Google Scholar] [CrossRef]
- Aggarwal, S.; Kumaraswami, M. Managing Manganese: The Role of Manganese Homeostasis in Streptococcal Pathogenesis. Front. Cell Dev. Biol. 2022, 10, 2022. [Google Scholar] [CrossRef]
- Mitchell, A.; Mitchell, T. Streptococcus pneumoniae: Virulence factors and variation. Clin. Microbiol. Infect. 2010, 16, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Crosby, H.A.; Tiwari, N.; Kwiecinski, J.M.; Xu, Z.; Dykstra, A.; Jenul, C.; Fuentes, E.J.; Horswill, A.R. The Staphylococcus aureus ArlRS two-component system regulates virulence factor expression through MgrA. Mol. Microbiol. 2020, 113, 103–122. [Google Scholar] [CrossRef]
- Mustafa, A.; Zulfiqar, U.; Mumtaz, M.Z.; Radziemska, M.; Haider, F.U.; Holatko, J.; Hammershmiedt, T.; Naveed, M.; Ali, H.; Kintl, A. Nickel (Ni) phytotoxicity and detoxification mechanisms: A review. Chemosphere 2023, 328, 138574. [Google Scholar] [CrossRef] [PubMed]
- Mora, D.; Arioli, S. Microbial urease in health and disease. PLoS Pathog. 2014, 10, e1004472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xu, C.; Qian, D.; Yu, Q.; Huang, M.; Zhou, L.; Qin, J.G.; Chen, L.; Li, E. Growth and health status of Pacific white shrimp, Litopenaeus vannamei, exposed to chronic water born cobalt. Fish Shellfish. Immunol. 2020, 100, 137–145. [Google Scholar] [CrossRef]
- Gopinath, K.; Venclovas, Č.; Ioerger, T.R.; Sacchettini, J.C.; McKinney, J.D.; Mizrahi, V.; Warner, D.F. A vitamin B12 transporter in Mycobacterium tuberculosis. Open Biol. 2013, 3, 120175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duell, B.L.; Su, Y.C.; Riesbeck, K. Host–pathogen interactions of nontypeable Haemophilus influenzae: From commensal to pathogen. FEBS Lett. 2016, 590, 3840–3853. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.; Mendel, R.R.; Ribbe, M.W. Molybdenum cofactors, enzymes and pathways. Nature 2009, 460, 839–847. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.-J.; Khan, F.; Khan, S.; Tabassum, N.; Mehta, S.; Kim, Y.-M. Pseudomonas aeruginosa virulence attenuation by inhibiting siderophore functions. Appl. Microbiol. Biotechnol. 2023, 107, 1019–1038. [Google Scholar] [CrossRef]
- Ladomersky, E.; Petris, M.J. Copper tolerance and virulence in bacteria. Metallomics 2015, 7, 957–964. [Google Scholar] [CrossRef] [Green Version]
- Ogunniyi, A.D.; Mahdi, L.K.; Jennings, M.P.; McEwan, A.G.; McDevitt, C.A.; Van der Hoek, M.B.; Bagley, C.J.; Hoffmann, P.; Gould, K.A.; Paton, J.C. Central role of manganese in regulation of stress responses, physiology, and metabolism in Streptococcus pneumoniae. J. Bacteriol. 2010, 192, 4489–4497. [Google Scholar] [CrossRef] [Green Version]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, J.; Guo, G.; Shen, H. Interactions between macrophages and biofilm during Staphylococcus aureus-associated implant infection: Difficulties and solutions. J. Innate Immun. 2023, 15, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.; Agerer, F.; Hauck, C.R.; Herrmann, M.; Ullrich, J.; Hacker, J.; Ohlsen, K. Global regulatory impact of ClpP protease of Staphylococcus aureus on regulons involved in virulence, oxidative stress response, autolysis, and DNA repair. J. Bacteriol. 2006, 188, 5783–5796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.M.; D’Orazio, S.E. Bacterial ureases: Structure, regulation of expression and role in pathogenesis. Mol. Microbiol. 1993, 9, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Yamaoka, Y. Survival of Helicobacter pylori in gastric acidic territory. Helicobacter 2017, 22, e12386. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Zhu, Y.; Ge, R. Molecular Mechanisms of Bismuth-containing Drugs Against Helicobacter pylori: A Further Update. Curr. Pharmacol. Rep. 2023, 9, 59–65. [Google Scholar] [CrossRef]
- Schalk, I.J.; Cunrath, O. An overview of the biological metal uptake pathways in P seudomonas aeruginosa. Environ. Microbiol. 2016, 18, 3227–3246. [Google Scholar] [CrossRef]
- Ghssein, G.; Ezzeddine, Z. A review of Pseudomonas aeruginosa metallophores: Pyoverdine, pyochelin and pseudopaline. Biology 2022, 11, 1711. [Google Scholar] [CrossRef]
- Kılıç, N.K.; Stensballe, A.; Otzen, D.E.; Dönmez, G. Proteomic changes in response to chromium (VI) toxicity in Pseudomonas aeruginosa. Bioresour. Technol. 2010, 101, 2134–2140. [Google Scholar] [CrossRef]
- Okamoto, S.; Eltis, L.D. The biological occurrence and trafficking of cobalt. Metallomics 2011, 3, 963–970. [Google Scholar] [CrossRef]
- Salam, L.B. Unravelling the antibiotic and heavy metal resistome of a chronically polluted soil. 3 Biotech 2020, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Westman, J.; Plumb, J.; Licht, A.; Yang, M.; Allert, S.; Naglik, J.R.; Hube, B.; Grinstein, S.; Maxson, M.E. Calcium-dependent ESCRT recruitment and lysosome exocytosis maintain epithelial integrity during Candida albicans invasion. Cell Rep. 2022, 38, 110187. [Google Scholar] [CrossRef] [PubMed]
- Jayaseelan, S.; Ramaswamy, D.; Dharmaraj, S. Pyocyanin: Production, applications, challenges and new insights. World J. Microbiol. Biotechnol. 2014, 30, 1159–1168. [Google Scholar] [CrossRef]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D. Cellular effects of pyocyanin, a secreted virulence factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norcross, E.W.; Sanders, M.E.; Moore III, Q.C.; Marquart, M.E. Pathogenesis of a clinical ocular strain of Streptococcus pneumoniae and the interaction of pneumolysin with corneal cells. J. Bacteriol. Parasitol. 2011, 2, 108. [Google Scholar] [CrossRef] [Green Version]
- Schmalstig, A.A.; Benoit, S.L.; Misra, S.K.; Sharp, J.S.; Maier, R.J. Noncatalytic antioxidant role for Helicobacter pylori urease. J. Bacteriol. 2018, 200, e00124-18. [Google Scholar] [CrossRef] [Green Version]
- Garland, M.M. Chemical Tools to Interrogate and Inhibit Bacterial Exotoxins; Stanford University: Stanford, CA, USA, 2021. [Google Scholar]
- Losada, L.; DebRoy, C.; Radune, D.; Kim, M.; Sanka, R.; Brinkac, L.; Kariyawasam, S.; Shelton, D.; Fratamico, P.M.; Kapur, V. Whole genome sequencing of diverse Shiga toxin-producing and non-producing Escherichia coli strains reveals a variety of virulence and novel antibiotic resistance plasmids. Plasmid 2016, 83, 8–11. [Google Scholar] [CrossRef]
- Ko, Y.-P.; Kuipers, A.; Freitag, C.M.; Jongerius, I.; Medina, E.; van Rooijen, W.J.; Spaan, A.N.; van Kessel, K.P.; Höök, M.; Rooijakkers, S.H. Phagocytosis escape by a Staphylococcus aureus protein that connects complement and coagulation proteins at the bacterial surface. PLoS Pathog. 2013, 9, e1003816. [Google Scholar] [CrossRef] [Green Version]
- Haan, L.D.; Hirst, T.R. Cholera toxin: A paradigm for multi-functional engagement of cellular mechanisms. Mol. Membr. Biol. 2004, 21, 77–92. [Google Scholar] [CrossRef]
- Porcheron, G.; Garénaux, A.; Proulx, J.; Sabri, M.; Dozois, C.M. Iron, copper, zinc, and manganese transport and regulation in pathogenic Enterobacteria: Correlations between strains, site of infection and the relative importance of the different metal transport systems for virulence. Front. Cell. Infect. Microbiol. 2013, 3, 90. [Google Scholar] [CrossRef] [Green Version]
- Iatsenko, I.; Marra, A.; Boquete, J.-P.; Peña, J.; Lemaitre, B. Iron sequestration by transferrin 1 mediates nutritional immunity in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2020, 117, 7317–7325. [Google Scholar] [CrossRef] [PubMed]
- Gurung, K.; Wertheim, B.; Falcao Salles, J. The microbiome of pest insects: It is not just bacteria. Entomol. Exp. Appl. 2019, 167, 156–170. [Google Scholar] [CrossRef] [Green Version]
- Loulou, A.; Mastore, M.; Caramella, S.; Bhat, A.H.; Brivio, M.F.; Machado, R.A.; Kallel, S. Entomopathogenic potential of bacteria associated with soil-borne nematodes and insect immune responses to their infection. PLoS ONE 2023, 18, e0280675. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Zhang, L.-j.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Bastakoti, S. Role of zinc in management of plant diseases: A review. Cogent Food Agric. 2023, 9, 2194483. [Google Scholar] [CrossRef]
- Glavinic, U.; Stankovic, B.; Draskovic, V.; Stevanovic, J.; Petrovic, T.; Lakic, N.; Stanimirovic, Z. Dietary amino acid and vitamin complex protects honey bee from immunosuppression caused by Nosema ceranae. PLoS ONE 2017, 12, e0187726. [Google Scholar] [CrossRef] [Green Version]
- Shafeeq, S.; Yesilkaya, H.; Kloosterman, T.G.; Narayanan, G.; Wandel, M.; Andrew, P.W.; Kuipers, O.P.; Morrissey, J.A. The cop operon is required for copper homeostasis and contributes to virulence in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 1255–1270. [Google Scholar] [CrossRef] [Green Version]
- Palmer, L.D.; Skaar, E.P. Transition metals and virulence in bacteria. Annu. Rev. Genet. 2016, 50, 67–91. [Google Scholar] [CrossRef] [Green Version]
- Price, S.L.; Vadyvaloo, V.; DeMarco, J.K.; Brady, A.; Gray, P.A.; Kehl-Fie, T.E.; Garneau-Tsodikova, S.; Perry, R.D.; Lawrenz, M.B. Yersiniabactin contributes to overcoming zinc restriction during Yersinia pestis infection of mammalian and insect hosts. Proc. Natl. Acad. Sci. USA 2021, 118, e2104073118. [Google Scholar] [CrossRef]
- Geiser, D.L.; Winzerling, J.J. Insect transferrins: Multifunctional proteins. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Qi, S.-S.; Gul, F.; Manan, S.; Rono, J.K.; Naz, M.; Shi, X.-N.; Zhang, H.; Dai, Z.-C.; Du, D.-L. A Green Approach Used for Heavy Metals ‘Phytoremediation’Via Invasive Plant Species to Mitigate Environmental Pollution: A Review. Plants 2023, 12, 725. [Google Scholar] [CrossRef] [PubMed]
- Sandy, M.; Butler, A. Microbial iron acquisition: Marine and terrestrial siderophores. Chem. Rev. 2009, 109, 4580–4595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, A.; Wilson, D. Essential metals at the host–pathogen interface: Nutritional immunity and micronutrient assimilation by human fungal pathogens. FEMS Yeast Res. 2015, 15, fov071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.P. Metal uptake in host-pathogen interactions: Role of iron in Porphyromonas gingivalis interactions with host organisms. Periodontol. 2000 2010, 52, 94. [Google Scholar] [CrossRef] [Green Version]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef]
- Vasil, M.L. How we learnt about iron acquisition in Pseudomonas aeruginosa: A series of very fortunate events. Biometals 2007, 20, 587–601. [Google Scholar] [CrossRef]
- Roth, M. Oxidative Stress Response: Lessons from the Gram-Negative Bacterium Escherichia coli; University of Geneva: Geneva, Switzerland, 2023. [Google Scholar]
- Waterman, S.R.; Hacham, M.; Hu, G.; Zhu, X.; Park, Y.-D.; Shin, S.; Panepinto, J.; Valyi-Nagy, T.; Beam, C.; Husain, S. Role of a CUF1/CTR4 copper regulatory axis in the virulence of Cryptococcus neoformans. J. Clin. Investig. 2007, 117, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Pokharel, P.; Dhakal, S.; Dozois, C.M. The Diversity of Escherichia coli Pathotypes and Vaccination Strategies against This Versatile Bacterial Pathogen. Microorganisms 2023, 11, 344. [Google Scholar] [CrossRef]
- Zhao, C.; Jia, X.; Pan, Y.; Liao, S.; Zhang, S.; Ji, C.; Kuang, G.; Wu, X.; Liu, Q.; Tang, Y. Thioredoxin A of Streptococcus suis Serotype 2 Contributes to Virulence by Inhibiting the Expression of Pentraxin 3 to Promote Survival Within Macrophages. J. Microbiol. 2023, 61, 433–448. [Google Scholar] [CrossRef]
- Heindl, J.E.; Hibbing, M.E.; Xu, J.; Natarajan, R.; Buechlein, A.M.; Fuqua, C. Discrete responses to limitation for iron and manganese in Agrobacterium tumefaciens: Influence on attachment and biofilm formation. J. Bacteriol. 2016, 198, 816–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.-H.; Fang, S.-B.; Chang, Y.-C.; Huang, S.-T.; Huang, C.-H.; Chang, P.-R.; Chang, W.-C.; Yang, L.T.-L.; Lin, P.-C.; Cheng, H.-Y. Effects of colonization-associated gene yqiC on global transcriptome, cellular respiration, and oxidative stress in Salmonella typhimurium. J. Biomed. Sci. 2022, 29, 102. [Google Scholar] [CrossRef] [PubMed]
- Ohradanova-Repic, A.; Praženicová, R.; Gebetsberger, L.; Moskalets, T.; Skrabana, R.; Cehlar, O.; Tajti, G.; Stockinger, H.; Leksa, V. Time to Kill and Time to Heal: The Multifaceted Role of Lactoferrin and Lactoferricin in Host Defense. Pharmaceutics 2023, 15, 1056. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.C.; Finlay, B.B. Targeting the type III secretion system to treat bacterial infections. Expert Opin. Ther. Targets 2014, 18, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Herget, T.; Klebl, B.; Ullrich, A. Interplay between mycobacteria and host signalling pathways. Nat. Rev. Microbiol. 2004, 2, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Dobrindt, U.; Hacker, J. Uropathogens and virulence factors. Urogenit. Infect. 2010, 1, 4–22. [Google Scholar]
- Ding, C.; Festa, R.A.; Sun, T.S.; Wang, Z.Y. Iron and copper as virulence modulators in human fungal pathogens. Mol. Microbiol. 2014, 93, 10–23. [Google Scholar] [CrossRef]
- Neyrolles, O.; Wolschendorf, F.; Mitra, A.; Niederweis, M. Mycobacteria, metals, and the macrophage. Immunol. Rev. 2015, 264, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Maurya, R.; Namdeo, M. Superoxide dismutase: A key enzyme for the survival of intracellular pathogens in host. In Reactive Oxygen Species; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Bombaywala, S.; Purohit, H.J.; Dafale, N.A. Mobility of antibiotic resistance and its co-occurrence with metal resistance in pathogens under oxidative stress. J. Environ. Manag. 2021, 297, 113315. [Google Scholar] [CrossRef]
- Brooks, L.R.; Mias, G.I. Streptococcus pneumoniae’s virulence and host immunity: Aging, diagnostics, and prevention. Front. Immunol. 2018, 9, 1366. [Google Scholar] [CrossRef] [Green Version]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Neoh, H.-M.; Nathan, S. Targeting Staphylococcus aureus toxins: A potential form of anti-virulence therapy. Toxins 2016, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, R.R.; Bittner, F. Cell biology of molybdenum. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashburn, L.M.; Jett, A.M.; Akins, D.R.; Whiteley, M. Staphylococcus aureus serves as an iron source for Pseudomonas aeruginosa during in vivo coculture. J. Bacteriol. 2005, 187, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Singh, P.; Srivastava, A. Synthesis, nature and utility of universal iron chelator–Siderophore: A review. Microbiol. Res. 2018, 212, 103–111. [Google Scholar] [CrossRef]
- Porcheron, G.; Dozois, C.M. Interplay between iron homeostasis and virulence: Fur and RyhB as major regulators of bacterial pathogenicity. Vet. Microbiol. 2015, 179, 2–14. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Schroll, A.; Sonnweber, T.; Weiss, G. The struggle for iron–a metal at the host–pathogen interface. Cell. Microbiol. 2010, 12, 1691–1702. [Google Scholar] [CrossRef]
- Kell, D.B.; Heyden, E.L.; Pretorius, E. The biology of lactoferrin, an iron-binding protein that can help defend against viruses and bacteria. Front. Immunol. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Yang, X.; Peng, W.; Wang, Y.; Yan, K.; Liu, Z.; Gao, T.; Yang, K.; Liu, W.; Guo, R.; Li, C. Mutations in troABCD against Copper Overload in a copA Mutant of Streptococcus suis. Appl. Environ. Microbiol. 2023, 89, e01841-22. [Google Scholar] [CrossRef]
- Focarelli, F.; Giachino, A.; Waldron, K.J. Copper microenvironments in the human body define patterns of copper adaptation in pathogenic bacteria. PLoS Pathog. 2022, 18, e1010617. [Google Scholar] [CrossRef]
- Virieux-Petit, M.; Hammer-Dedet, F.; Aujoulat, F.; Jumas-Bilak, E.; Romano-Bertrand, S. From copper tolerance to resistance in pseudomonas aeruginosa towards patho-adaptation and hospital success. Genes 2022, 13, 301. [Google Scholar] [CrossRef] [PubMed]
- Secli, V.; Di Biagio, C.; Martini, A.; Michetti, E.; Pacello, F.; Ammendola, S.; Battistoni, A. Localized Infections with P. aeruginosa Strains Defective in Zinc Uptake Reveal That Zebrafish Embryos Recapitulate Nutritional Immunity Responses of Higher Eukaryotes. Int. J. Mol. Sci. 2023, 24, 944. [Google Scholar] [CrossRef] [PubMed]
- Gabbianelli, R.; Scotti, R.; Ammendola, S.; Petrarca, P.; Nicolini, L.; Battistoni, A. Role of ZnuABC and ZinT in Escherichia coliO157: H7 zinc acquisition and interaction with epithelial cells. BMC Microbiol. 2011, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Carter, E.L.; Flugga, N.; Boer, J.L.; Mulrooney, S.B.; Hausinger, R.P. Interplay of metal ions and urease. Metallomics 2009, 1, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acheson, D.W.; Luccioli, S. Mucosal immune responses. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, S.; Iwamoto, T.; Takada, K.; Okuda, K.-i.; Tajima, A.; Iwase, T.; Mizunoe, Y. Staphylococcus epidermidis Esp degrades specific proteins associated with Staphylococcus aureus biofilm formation and host-pathogen interaction. J. Bacteriol. 2013, 195, 1645–1655. [Google Scholar] [CrossRef] [Green Version]
- Chhipa, H.; Joshi, P. Nanofertilisers, nanopesticides and nanosensors in agriculture. Nanosci. Food Agric. 1 2016, 20, 247–282. [Google Scholar]
- Gauthier, G.M.; Keller, N.P. Crossover fungal pathogens: The biology and pathogenesis of fungi capable of crossing kingdoms to infect plants and humans. Fungal Genet. Biol. 2013, 61, 146–157. [Google Scholar] [CrossRef]
- Vignesh, K.S.; Deepe, G.S., Jr. Immunological orchestration of zinc homeostasis: The battle between host mechanisms and pathogen defenses. Arch. Biochem. Biophys. 2016, 611, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Rohr, J.R.; Barrett, C.B.; Civitello, D.J.; Craft, M.E.; Delius, B.; DeLeo, G.A.; Hudson, P.J.; Jouanard, N.; Nguyen, K.H.; Ostfeld, R.S. Emerging human infectious diseases and the links to global food production. Nat. Sustain. 2019, 2, 445–456. [Google Scholar] [CrossRef]
- Chassy, B.M. Food safety risks and consumer health. New Biotechnol. 2010, 27, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Frei, A.; Verderosa, A.D.; Elliott, A.G.; Zuegg, J.; Blaskovich, M.A. Metals to combat antimicrobial resistance. Nat. Rev. Chem. 2023, 7, 202–224. [Google Scholar] [CrossRef]
- Sekhon, B.S. Nanotechnology in agri-food production: An overview. Nanotechnol. Sci. Appl. 2014, 7, 31–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, M.; Ingle, A. Role of nanotechnology in agriculture with special reference to management of insect pests. Appl. Microbiol. Biotechnol. 2012, 94, 287–293. [Google Scholar] [CrossRef]
- Morkunas, I.; Woźniak, A.; Mai, V.C.; Rucińska-Sobkowiak, R.; Jeandet, P. The role of heavy metals in plant response to biotic stress. Molecules 2018, 23, 2320. [Google Scholar] [CrossRef] [Green Version]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Karman, S.B.; Diah, S.Z.M.; Gebeshuber, I.C. Raw materials synthesis from heavy metal industry effluents with bioremediation and phytomining: A biomimetic resource management approach. Adv. Mater. Sci. Eng. 2015, 2015, 185071. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fe | Role in Plants | Role in Insects | Role in Microbes |
|---|---|---|---|
| Absorption | Plants take up Fe through their roots and transport it to different plant parts for use in metabolic processes [51]. | Insects acquire Fe from their diet, including plant material [52]. | Microbes acquire Fe from their environment for growth and metabolism [53]. |
| Defense | Fe produces defensive compounds in plants, such as phytochelatin and lignin [54]. | Insects can use Fe-binding proteins to sequester Fe from their diet and prevent it from being used by pathogens [55]. | Some microbes can produce Fe-chelating compounds called siderophores, competing with plants and insects for Fe [56]. |
| Signaling | Fe is involved in signaling pathways in plants, e.g., the regulation of gene expression and responses to stress [57]. | Fe can act as a signaling molecule in insects, affecting behavior and development [58]. | Fe availability can also affect microbial gene expression and behavior [59]. |
| Regulation | Plants regulate Fe acquisition and transport through a complex system of genes and proteins [60]. | Insects require a certain amount of Fe for proper growth and development, but excess Fe can be toxic [61]. | Microbes have evolved mechanisms to sense and respond to Fe availability in their environment [62]. |
| Insects | Metals | Defensive Mechanisms | References |
|---|---|---|---|
| Ironclad beetle | Fe | Hardens exoskeleton for protection against predators. | [120] |
| Bombardier Beetle | Cu | Copper enzymes facilitate the expulsion of toxic sprays. | [121] |
| Leafhopper | Zn | Protection of exoskeleton for defense against predators. | [122] |
| Silverfish | Ag | Antimicrobial defense, silver scales provide camouflage and protection. | [123] |
| Sweat Bees | Hg | It uses mercury to deter parasites and pathogens. | [124] |
| Fireflies | Co | Cobalt ions play a role in the production of bioluminescence. | [125] |
| Nickel-Eating Moth | Ni | Nickel detoxification enables feeding on nickel-rich plants. | [126] |
| Selenium-Superior Fly | Se | Selenium detoxification offers resistance to toxic plants. | [127] |
| Microbes | Virulence Factor | Role of Trace Metals | References |
|---|---|---|---|
| Candida albicans | Phospholipase enzymes | Ca: Activates and stabilizes phospholipase enzymes, also secretes candida lysin. | [165] |
| Pseudomonas aeruginosa | Pyocyanin | Fe: Crucial for pyocyanin synthesis, a virulent pigment. | [166,167] |
| Streptococcus pneumoniae | Pneumolysin | Zn: Activates pneumolysin, causing host cell damage. | [168] |
| Helicobacter pylori | Urease | Ni: Essential for urease activity, aiding survival in an acidic stomach. | [169] |
| Escherichia coli | Shiga toxin | Fe, Zn, Cu, and Mg: Essential for toxin production and stability. | [170,171] |
| Staphylococcus aureus | Coagulase | Ca: Activates coagulase and aids in blood clotting. | [172] |
| Vibrio cholerae | Cholera toxin | Zn: Stabilizes cholera toxin structure for practical function. | [173] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Lang, M. A Comprehensive Review on the Roles of Metals Mediating Insect–Microbial Pathogen Interactions. Metabolites 2023, 13, 839. https://doi.org/10.3390/metabo13070839
Khan S, Lang M. A Comprehensive Review on the Roles of Metals Mediating Insect–Microbial Pathogen Interactions. Metabolites. 2023; 13(7):839. https://doi.org/10.3390/metabo13070839
Chicago/Turabian StyleKhan, Subhanullah, and Minglin Lang. 2023. "A Comprehensive Review on the Roles of Metals Mediating Insect–Microbial Pathogen Interactions" Metabolites 13, no. 7: 839. https://doi.org/10.3390/metabo13070839
APA StyleKhan, S., & Lang, M. (2023). A Comprehensive Review on the Roles of Metals Mediating Insect–Microbial Pathogen Interactions. Metabolites, 13(7), 839. https://doi.org/10.3390/metabo13070839