Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling and Sample Preparation
2.2. NMR Experiments

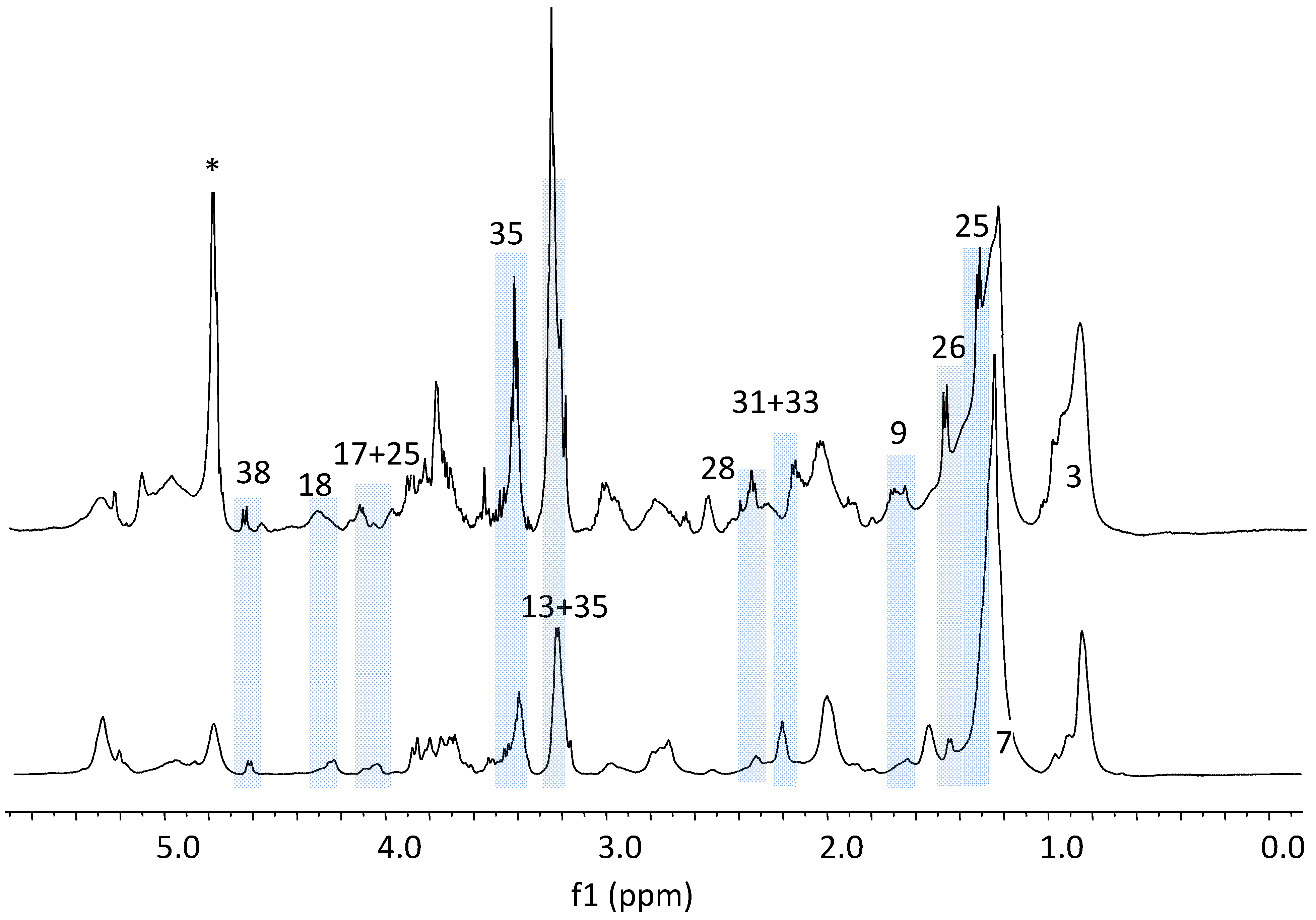{kind=link}
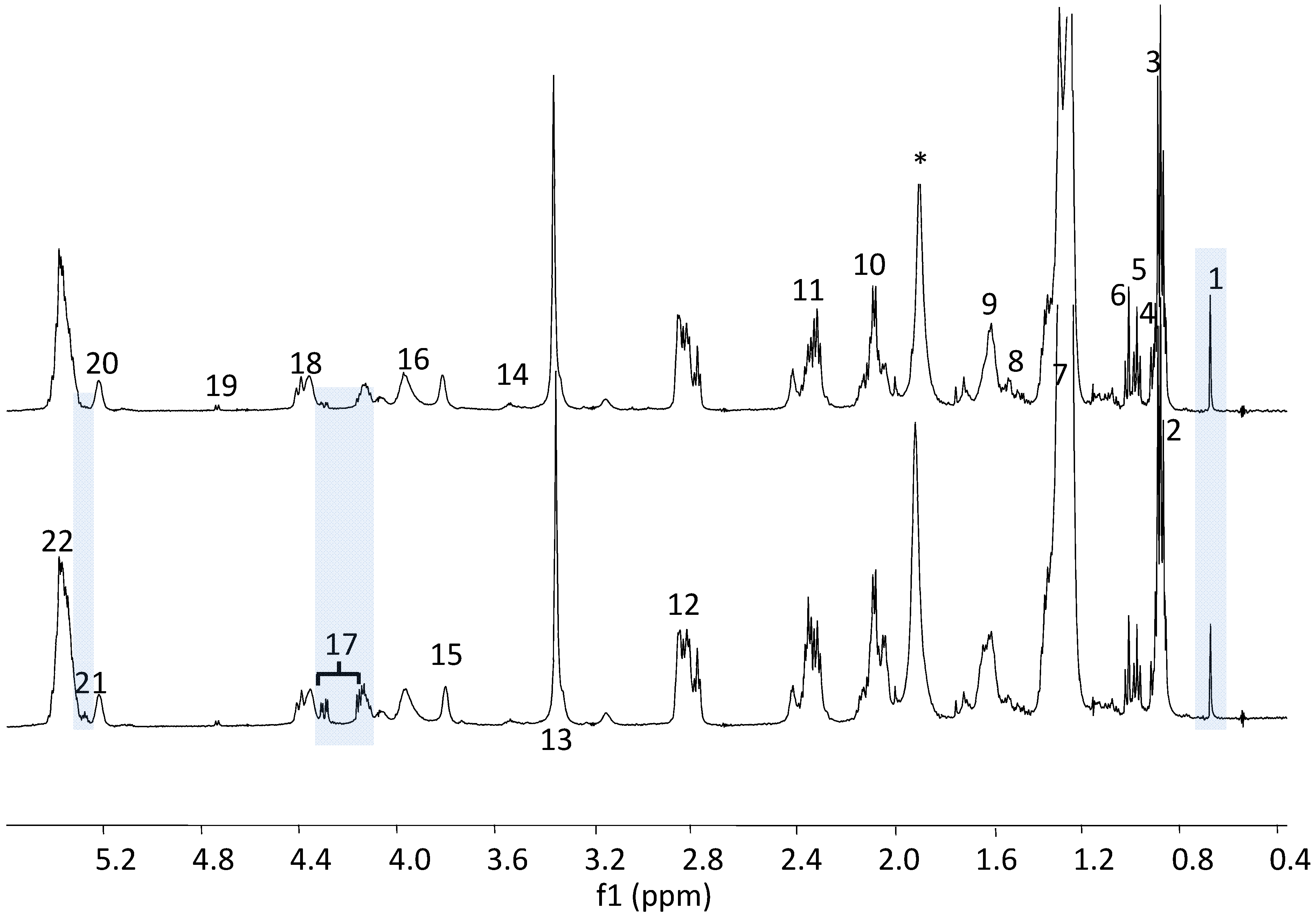{kind=link}
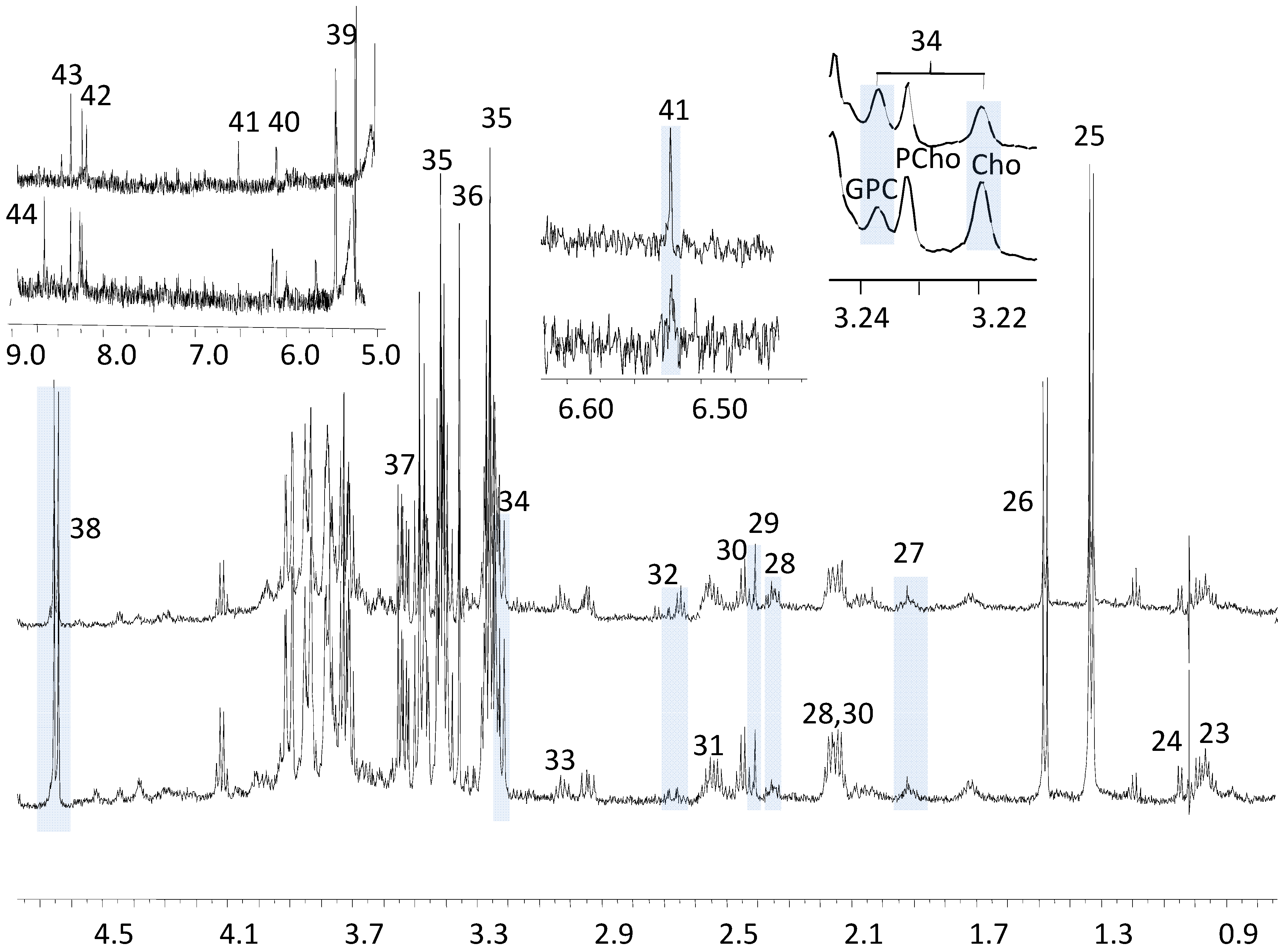{kind=link}
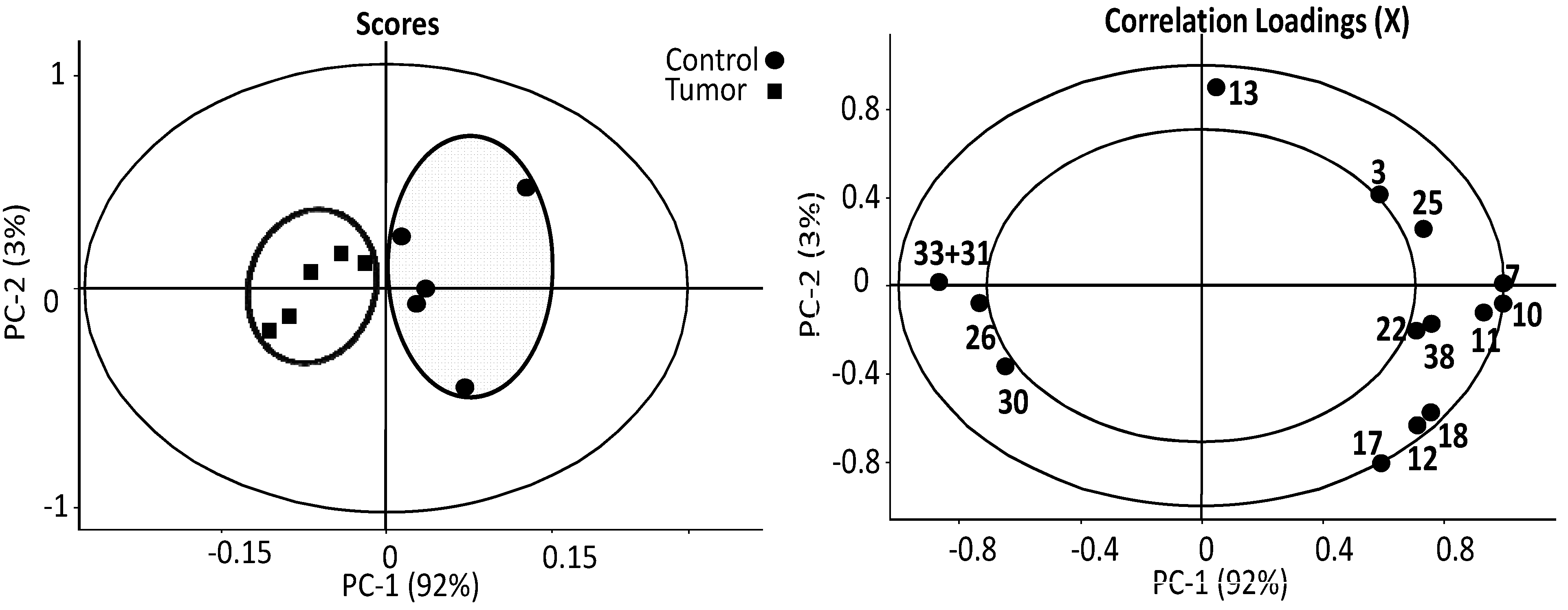{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Peaks’ Signals | Chemical Shifts | Concentrations in NMR tube: (µM/mg) Mean ± SD | Estimates of absolute Concentrations in tissue: (mM) Mean ± SD | ||
|---|---|---|---|---|---|---|
| (ppm) | ||||||
| Tumor | Control | Tumor | Control | |||
| 1 | Cholesterol 18-CH3 | 0.67 (s) | 1.03 ± 0.01 | 0.59 ± 0.09 | 0.62 ± 0.06 | 0.36 ± 0.06 |
| 2 | Cholesterol 26, 27-CH3 | 0.87 (d) | 1.03 ± 0.01 | 0.59 ± 0.09 | 0.62 ± 0.06 | 0.36 ± 0.06 |
| 3 | Lipids CH3 | 0.88 (t) | 1.43 ± 0.17 | 18.36 ± 0.72 | 8.56 ± 1.03 | 11.02 ± 0.43 |
| 4 | Cholesterol 21-CH3 | 0.90 (d) | 1.03 ± 0.10 | 0.59 ± 0.09 | 0.62 ± 0.06 | 0.36 ± 0.06 |
| 5 | Ω3 CH3 | 0.97 (t) | 4.36 ± 0.52 | 3.64 ± 0.26 | 2.62 ± 0.31 | 2.18 ± 0.16 |
| 6 | Free cholesterol 19-CH3 | 1.01 (t) | 1.03 ± 0.10 | 0.594 ± 0.092 | 0.62 ± 0.06 | 0.36 ± 0.06 |
| 7 | Lipids CH2 | 1.27 (m) | 58.66 ± 13.21 | 74.28 ± 2.38 | 35.19 ± 7.92 | 44.57 ± 1.43 |
| 8 | Cholesterol CH2 | 1.45–1.50 (m) | 1.03 ± 0.10 | 0.59 ± 0.09 | 0.62 ± 0.06 | 0.36 ± 0.06 |
| 9 | Lipids CH2CH2CO | 1.52–1.61 (m) | 5.88 ± 1.26 | 7.99 ± 0.44 | 3.53 ± 0.75 | 4.79 ± 0.26 |
| 10 | Lipids CH2CH=C | 1.96–2.15 (m) | 8.28 ± 1.51 | 11.33 ± 0.51 | 4.97 ± 0.90 | 6.80 ± 0.30 |
| 11 | Lipids CH2CO | 2.22–2.42 (m) | 7.59 ± 1.67 | 9.62 ± 0.38 | 4.55 ± 1.00 | 5.77 ± 0.23 |
| 12 | Lipids =CHCH2CH= | 2.74–2.91 (m) | 7.98 ± 1.74 | 8.71 ± 0.62 | 4.79 ± 1.04 | 5.23 ± 0.37 |
| 13 | Choline N(CH3)3 | 3.36 (s) | 28.81 ± 3.78 | 33.36 ± 1.06 | 17.29 ± 2.27 | 20.01 ± 0.64 |
| 14 | Cholesterol 3-CHOH | 3.53 (d) | 0.71 ± 0.15 | 0.73 ± 0.02 | 0.43 ± 0.09 | 0.44 ± 0.01 |
| 15 | Phosphatidylcholine N-CH2 | 3.81 (m) | 3.44 ± 0.54 | 3.48 ± 0.26 | 2.06 ± 0.33 | 2.09 ± 0.16 |
| 16 | Glycerophospholipid backbone 3-CH2 | 3.97 (m) | 1.51 ± 0.44 | 2.05 ± 0.08 | 0.91 ± 0.26 | 1.23 ± 0.05 |
| 17 | Glycerol backbone 1,3-CH2 | 4.16–4.30 (m) | 0.15 ± 0.13 | 0.73 ± 0.09 | 0.09 ± 0.08 | 0.44 ± 0.05 |
| 18 | Phosphatidylcholine PO-CH2 | 4.35 (m) | 2.66 ± 0.44 | 2.70 ± 0.10 | 1.60 ± 0.27 | 1.62 ± 0.06 |
| 19 | Esterified cholesterol 3-CHOH | 4.72 (d) | 0.55 ± 0.13 | 0.48 ± 0.06 | 0.33 ± 0.08 | 0.29 ± 0.04 |
| 20 | Glycerophospholipid backbone 2-CH | 5.22 (m) | 1.51 ± 0.44 | 2.05 ± 0.08 | 0.91 ± 0.26 | 1.23 ± 0.05 |
| 21 | Glycerol backbone 2-CH | 5.28 (m) | 0.037 ± 0.08 | 0.63 ± 0.37 | 0.02 ± 0.05 | 0.38 ± 0.22 |
| 22 | Lipid CH=CH. | 5.37 (m) | 11.36 ± 2.57 | 13.18 ± 0.67 | 6.82 ± 1.54 | 7.91 ± 0.40 |
| 23 | Leucine | 0.99 (d), 1.70 (m), 3.72 (m) | 1.17 ± 0.29 | 0.80 ± 0.27 | 0.70 ± 0.17 | 0.48 ± 0.16 |
| 24 | Valine | 0.97 (d), 1.02 (d), 2.28 (m), 3.61 (d) | 0.84 ± 0.32 | 0.75 ± 0.39 | 0.50 ± 0.19 | 0.45 ± 0.23 |
| 25 | Lactate | 1.32 (d), 4.2 (q) | 11.56 ± 5.00 | 18.58 ± 6.75 | 6.94 ± 3.00 | 11.15 ± 4.05 |
| 26 | Alanine | 1.47 (d), 3.77 (m) | 6.59 ± 1.25 | 6.52 ± 2.23 | 3.95 ± 0.75 | 3.91 ± 1.338 |
| 27 | Acetate | 1.92 (s) | 0.63 ± 0.20 | 0.27 ± 0.09 | 0.38 ± 0.12 | 0.16 ± 0.054 |
| 28 | Glutamate | 2.05 (m), 2.36 (dt), 3.76 (m) | 3.64 ± 1.34 | 1.38 ± 0.33 | 2.18 ± 0.80 | 0.83 ± 0.20 |
| 29 | Succinate | 2.41 (s) | 0.81 ± 0.11 | 1.11 ± 0.41 | 0.49 ± 0.07 | 0.67 ± 0.25 |
| 30 | Glutamine | 3.77 (m), 2.46 (m), 2.14 (m) | 4.08 ± 1.40 | 6.34 ± 2.58 | 2.45 ± 0.84 | 3.80 ± 1.55 |
| 31 | Glutathione | 2.16 (m), 2.54 (m), 2.97 (m), 3.77 (m), 4.58 (dd) | 2.64 ± 1.96 | 2.48 ± 1.42 | 1.58 ± 1.18 | 1.49 ± 0.85 |
| 32 | Malate | 2.35 (dd), 2.65 (dd), 4.26 (d) | 2.17 ± 0.57 | 1.21 ± 0.53 | 1.30 ± 0.34 | 0.73 ± 0.32 |
| 33 | Creatine + Creatinine | 3.08 (s) | 0.33 ± 0.12 | 0.32 ± 0.09 | 0.20 ± 0.072 | 0.19 ± 0.05 |
| 34 | Choline | 3.22–3.24, 3.50(m) | 0.70 ± 0.10 | 0.65 ± 0.26 | 0.42 ± 0.06 | 0.39 ± 0.16 |
| 35 | Taurine | 3.25 (t), 3.40 (t) | 21.95 ± 6.22 | 24.61 ± 6.23 | 13.17 ± 3.73 | 14.77 ± 3.74 |
| 36 | Proline+Inositol | 3.37 (m) | 0.70 ± 0.42 | 0.90 ± 0.36 | 0.42 ± 0.25 | 0.54 ± 0.22 |
| 37 | Glycine | 3.55 (s) | 2.68 ± 1.40 | 3.12 ± 1.78 | 1.61 ± 0.84 | 1.87 ± 1.07 |
| 38 | â-Glucose | 3.41 (dd), 3.47 (dd), 3.92 (m), 4.65 (d) | 37.55 ± 25.83 | 74.75 ± 21.53 | 22.53 ± 15.50 | 44.85 ± 12.92 |
| 39 | á-Glucose | 3.42 (m), 3.55 (t), 3.76 (m) 3.81 (m), 3.92 (m), 5.23 (d) | 0.80 ± 0.40 | 0.87 ± 0.44 | 0.48 ± 0.24 | 0.52 ± 0.26 |
| 40 | Inosine Derivatives | 4.42 (dd), 4.82 (t), 6.16 (d), 4.32 (m), 3.83 (dd), 3.91 (dd), 8.21 (s), 8.32 (s) | ||||
| 41 | Fumarate | 6.75 (s) | 0.23 ± 0.13 | 0.08 ± 0.02 | 0.14 ± 0.08 | 0.05 ± 0.012 |
| 42 | ATP/ADP | 4.22 (m), 4.37 (m), 4.57 (d), 6.13 (d), 8.26 (s), 8.52 (s) | 1.12 ± 0.75 | 2.22 ± 0.56 | 0.67 ± 0.45 | 1.33 ± 0.34 |
| 43 | Formate | 8.34 (s) | 0.63 ± 0.25 | 0.7 ± 0.28 | 0.38 ± 0.15 | 0.42 ± 0.17 |
| 44 | -NH. | 9.01 | ||||
| 45 | sn-Glycero-3-phosphocholine | 3.24, 3.50 (m) | 1.25 ± 0.30 | 0.65 ± 0.16 | 0.75 ± 0.18 | 0.39 ± 0.10 |
| 46 | 2-Oxoglutarate | 2.47 (t), 3.01 (t) | 1.11 ± 0.42 | 0.85 ± 0.40 | 0.67 ± 0.25 | 0.51 ± 0.24 |
| 47 | TMAO | 3.25 (s) | 1.02 ± 0.45 | 0.66 ± 0.25 | 0.61 ± 0.27 | 0.40 ± 0.15 |
| 48 | O-Phosphocholine | 3.23, 3.50 (m) | 0.59 ± 0.34 | 0.44 ± 0.20 | 0.35 ± 0.20 | 0.26 ± 0.12 |
| 49 | Hypoxanthine | 8.15 (s) | 0.70 ± 0.42 | 0.90 ± 0.36 | 0.42 ± 0.25 | 0.54 ± 0.22 |
| 50 | Dimethylamine | 2.71 (s) | 0.12 ± 0.07 | 0.10 ± 0.01 | 0.07 ± 0.04 | 0.06 ± 0.01 |
| 51 | Isoleucine | 0.91 (t),1.00 (d), 1.25 (m), 1.47 (m), 1.97 (m), 3.65 (d) | 0.55 ± 0.31 | 0.65 ± 0.13 | 0.33 ± 0.19 | 0.39 ± 0.08 |
2.3. Statistical Analysis
| Peaks’ Signals | Chemical Shifts | Relative Conc. Mean ± SD | ||
|---|---|---|---|---|
| (ppm) | Tumor | Control | ||
| 3 | Methyl | 0.88 | 0.87 ± 0.20 | 1.00 ± 0.06 |
| 7 | Lipid CH2 | 1.28 (m) | 1.49 ± 0.32 | 3.02 ± 0.71 |
| 13 | Choline | 3.22–3.24 (m) | 0.68 ± 0.10 | 0.73 ± 0.05 |
| 17 | Glycerol | 4.16–4.30 (m) | 0.02 ± 0.01 | 0.04 ± 0.02 |
| 18 | PhosphatidylCholine | 4.35 (m) | 0.04 ± 0.01 | 0.08 ± 0.03 |
| 25 | Lactate | 1.32 (d), 4.2 (q) | 0.16 ± 0.03 | 0.49 ± 0.28 |
| 26 | Alanine | 1.47 (d), 3.77 (m) | 0.09 ± 0.01 | 0.06 ± 0.01 |
| 28 | Glutamate | 2.05 (m) | 0.02 ± 0.01 | 0.01 ± 0.01 |
| 35 | Glycine | 3.55 (s) | 0.32 ± 0.14 | 0.46 ± 0.21 |
3. Results
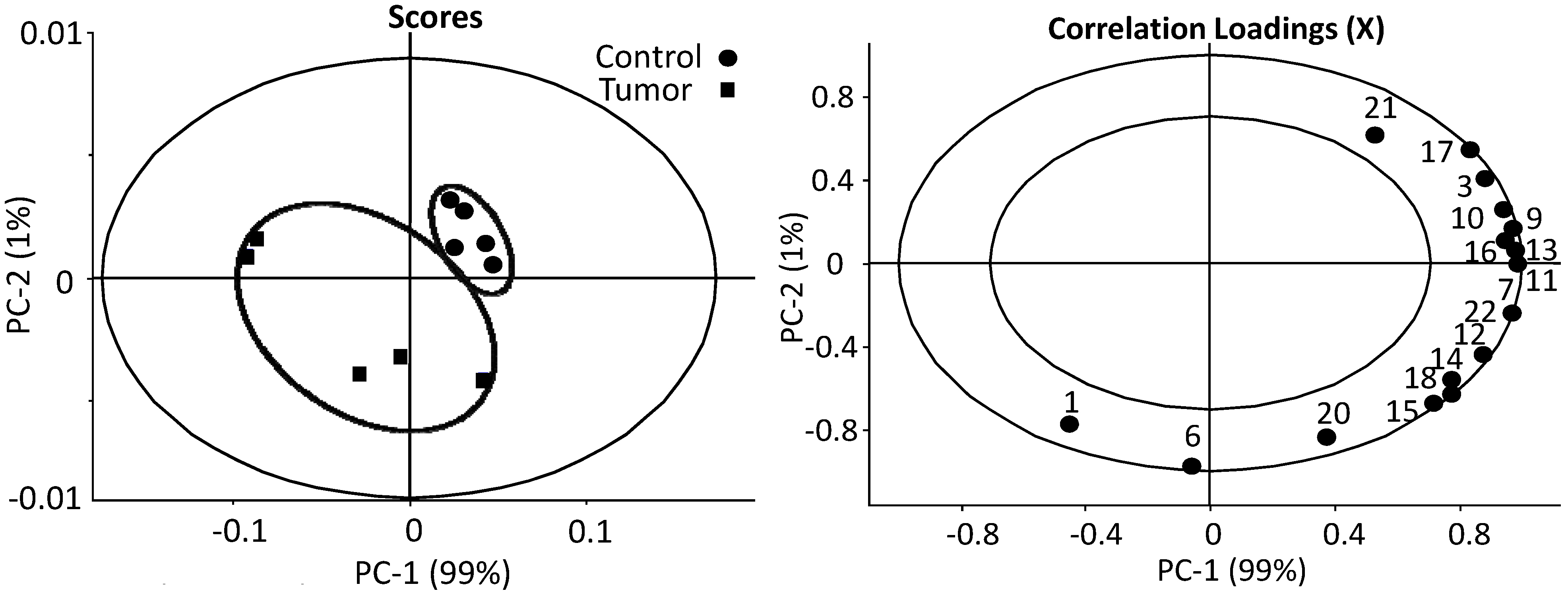
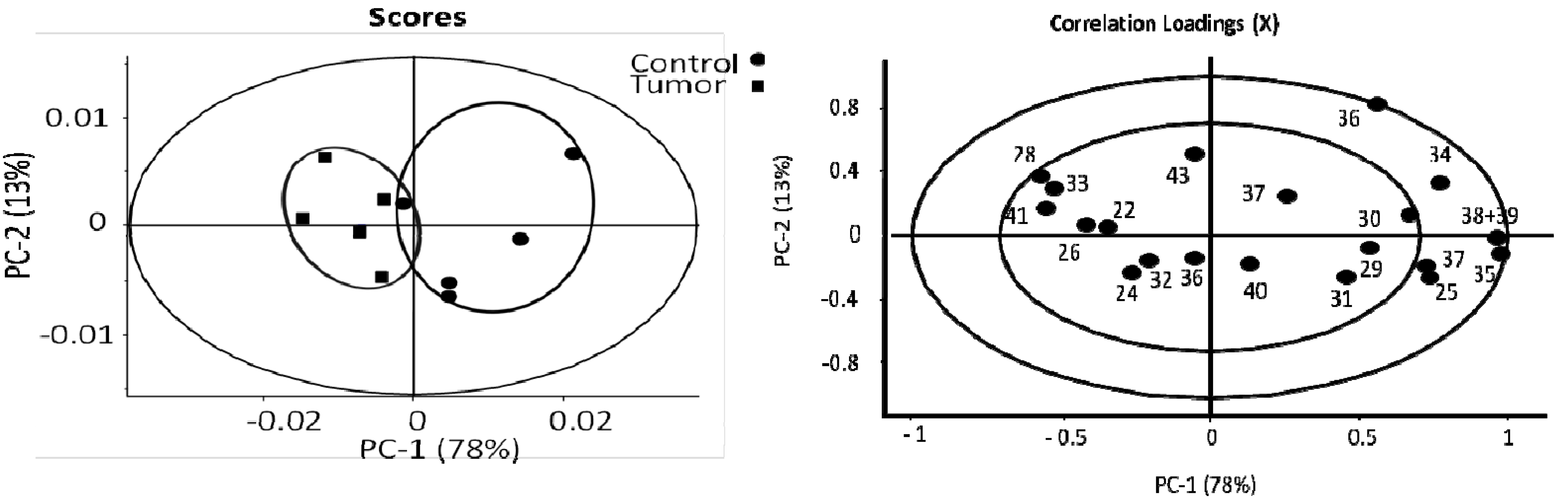
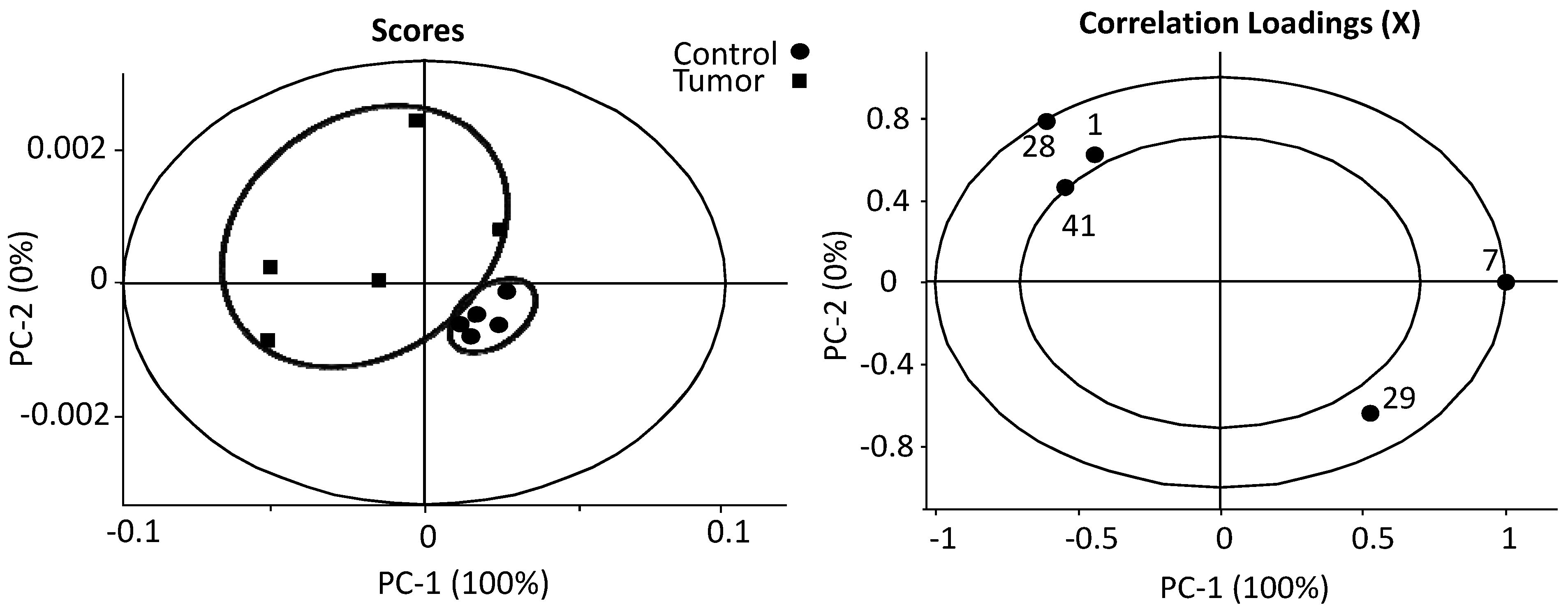
3.1. The Relative Intensities of Metabolites in NMR Spectroscopy


3.2. The Estimates of Absolute Peak Intensities of Metabolites and Statistical Analysis




4. Discussion


5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Beckonert, O.; Coen, M.; Keun, H.C.; Wang, Y.L.; Ebbels, T.M.D.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. High-resolution magic-angle-spinning NMR spectroscopy for metabolic profiling of intact tissues. Nat. Protoc. 2010, 5, 1019–1032. [Google Scholar] [CrossRef]
- Holmes, E.; Nicholson, J.K.; Nicholls, A.W.; Lindon, J.C.; Connor, S.C.; Polley, S.; Connelly, J. The identification of novel biomarkers of renal toxicity using automatic data reduction techniques and PCA of proton NMR spectra of urine. Chemom. Intell. Lab. Syst. 1998, 44, 245–255. [Google Scholar] [CrossRef]
- Watson, M. Drugs in clinical development for melanoma†: Summary and table. Pharm. Med. 2012, 26, 171–183. [Google Scholar] [CrossRef]
- Jerant, A.F.; Johnson, J.T.; Sheridan, C.D.; Caffrey, T.J. Early detection and treatment of skin cancer. Am. Fam. Physician 2000, 62, 357–368. [Google Scholar]
- Morvan, D.; Demidem, A.; Papon, J.; de Latour, M.; Madelmont, J.C. Melanoma tumors acquire a new phospholipid metabolism phenotype under cystemustine as revealed by high-resolution magic angle spinning proton nuclear magnetic resonance spectroscopy of intact tumor sampled. Cancer Res. 2002, 62, 1890–1897. [Google Scholar]
- Morvan, D.; Demidem, A.; Guenin, S.; Madelmont, J.C. Methionine-dependence phenotype of tumors: Metabolite profiling in a melanoma model using L-[methyl-13C]-methionine and high-resolution magic angle spinning 1H-13C nuclear magnetic resonance spectroscopy. Magn. Reson. Med. 2006, 55, 984–996. [Google Scholar] [CrossRef]
- Li, W.; Slominski, R.; Slominski, A.T. High-resolution magic angle spinning nuclear magnetic resonance analysis of metabolic changes in melanoma cells after induction of melanogenesis. Anal. Biochem. 2009, 386, 282–284. [Google Scholar] [CrossRef]
- Morvan, D.; Demidem, A.; Papon, J.; Madelmont, J.C. Quantitative HRMAS proton total correlation spectroscopy applied to cultured melanoma cells treated by chloroethyl nitrosourea: Demonstration of phospholipid metabolism alterations. Magn. Reson. Med. 2003, 49, 241–248. [Google Scholar] [CrossRef]
- Fages, A.; Morvan, D.; Schwartz, L.; Steyaert, J.M.; Stepien, G.; Demidem, A. Disturbance of metabolic pathways of glucose consumption by CENU treatment in B16 melanoma tumors: A NMR spectroscopy-based [1,2-13C]glucose fluxomics. Bull. du Cancer 2010, 97, S39–S40. [Google Scholar]
- Guitera, P.; Bourgeat, P.; Stretch, J.R.; Scolyer, R.A.; Ourselin, S.; Lean, C.; Thompson, J.F.; Bourne, R. Diagnostic value of 8.5 T magnetic resonance spectroscopy of benign and malignant skin lesion biopsies. Melanoma Res. 2010, 20, 311–317. [Google Scholar] [CrossRef]
- Triba, M.N.; Starzec, A.; Bouchemal, N.; Guenin, E.; Perret, G.Y.; Le Moyec, L. Metabolomic profiling with NMR discriminates between biphosphonate and doxorubicin effects on B16 melanoma cells. NMR Biomed. 2010, 23, 1009–1016. [Google Scholar] [CrossRef]
- Bourne, R.M.; Stanwell, P.; Stretch, J.R.; Scolyer, R.A.; Thompson, J.F.; Mountford, C.E.; Lean, C.L. In vivo and ex vivo proton MR spectroscopy of primary and secondary melanoma. Eur. J. Radiol. 2005, 53, 506–513. [Google Scholar] [CrossRef]
- Stretch, J.R.; Somorjai, R.; Bourne, R.; Hsiao, E.; Scolyer, R.A.; Dolenko, B.; Thompson, J.F.; Mountford, C.E.; Lean, C.L. Melanoma metastases in regional lymph nodes are accurately detected by proton magnetic resonance spectroscopy of fine-needle aspirate biopsy samples. Ann. Surg. Oncol. 2005, 12, 943–949. [Google Scholar] [CrossRef]
- Maire, C.; Kluza, J.; Jendoubi, M.; Corazoa, P.; Guerreshi, P.; Touil, Y.; Polokowska, R.; Formstecher, P.; Mortier, L.; Marchetti, P. HIF-1 alpha is an important orchestrator of bioenergetic metabolism in melanoma. J. Investig. Dermatol. 2011, 131, 2152. [Google Scholar]
- Comito, G.; Calvani, M.; Giannoni, E.; Bianchini, F.; Calorini, L.; Torre, E.; Migliore, C.; Giordano, S.; Chiarugi, P. HIF-1 alpha stabilization by mitochondrial ROS promotes Met-dependent invasive growth and vasculogenic mimicry in melanoma cells. Free Radic. Biol. Med. 2011, 51, 893–904. [Google Scholar] [CrossRef]
- Trisciuoglio, D.; Gabellini, C.; Desideri, M.; Ziparo, E.; del Bufalo, D. Bcl-2 regulates HIF-1a protein stabilization in hypoxic melanoma cells via the molecular chaperone HSP90 beta. EJC Suppl. 2010, 8, 153–153. [Google Scholar]
- Zaidi, M.R.; Day, C.P.; Merlino, G. From UVs to metastases: Modeling melanoma initiation and progression in the mouse. J. Investig. Dermatol. 2008, 128, 2381–2391. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, Y.; Kim, M.; Kim, D.S.; Lee, S.C.; Chi, S.W.; Lee, D.H.; Park, S.G.; Park, B.C.; Bae, K.H.; et al. Comparative proteomic analysis of mouse melanoma cell line B16, a metastatic descendant B16F10, and B16 overexpressing the metastasis-associated tyrosine phosphatase PRL-3. Oncol. Res. 2009, 17, 601–612. [Google Scholar] [CrossRef]
- Yoshimura, K.; Meckel, K.F.; Laird, L.S.; Chia, C.Y.; Park, J.J.; Olino, K.L.; Tsunedomi, R.; Harada, T.; Iizuka, N.; Hazama, S.; et al. Integrin alpha 2 mediates selective metastasis to the liver. Cancer Res. 2009, 69, 7320–7328. [Google Scholar] [CrossRef]
- Overwijk, W.W.; Restifo, N.P. B16 as a Mouse Model for Human Melanoma. In Current Protocols in Immunology; John Wiley & Sons, Inc.: Hoboken, NJ., USA, 2001. [Google Scholar]
- Valle, E.F.; Zalka, A.D.; Groszek, L.; Stackpole, C.W. Patterning of B16-melanoma metastasis and colonization generally relates to tumor-cell growth-stimulating or growth-inhibiting effects of organs and tissues. Clin. Exp. Metastasis 1992, 10, 419–429. [Google Scholar] [CrossRef]
- Stackpole, C.W.; Valle, E.F.; Alterman, A.L. B16 melanoma metastasis to an artificial organ implant. Cancer Res. 1991, 51, 2444–2450. [Google Scholar]
- Larue, L.; Beermann, F. Cutaneous melanoma in genetically modified animals. Pigment Cell Res. 2007, 20, 485–497. [Google Scholar] [CrossRef]
- Gheorgheosu, D.; Dehelean, C.; Cristea, M.; Muntean, D. Development of the B16 murine melanoma model. Ann. RSCB 2011, 2, 148–152. [Google Scholar]
- Roomi, M.W.; Kalinovsky, T.; Roomi, N.W.; Ivanov, V.; Rath, M.; Niedzwiecki, A. Suppression of growth and hepatic metastasis of murine B16FO melanoma cells by a novel nutrient mixture. Oncol. Rep. 2008, 20, 809–817. [Google Scholar]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.G.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef]
- Washburn, K.W. A modification of the folch method of lipid extraction for poultry. Poult. Sci. 1989, 68, 1425–1427. [Google Scholar] [CrossRef]
- Iverson, S.J.; Lang, S.L.C.; Cooper, M.H. Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef]
- Akoh, C.C.; Min, D.B. Food Lipids: Chemistry, Nutrition, and Biotechnology, 3rd ed.; Food Science and Technology: New York, NY., USA, 2008. [Google Scholar]
- Govindaraju, V.; Young, K.; Maudsley, A.A. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed. 2000, 13, 129–153. [Google Scholar] [CrossRef]
- Martin, F.P.J.; Dumas, M.E.; Wang, Y.L.; Legido-Quigley, C.; Yap, I.K.S.; Tang, H.R.; Zirah, S.; Murphy, G.M.; Cloarec, O.; Lindon, J.C.; et al. A top-down systems biology view of microbiome-mammalian metabolic interactions in a mouse model. Mol. Syst. Biol. 2007, 3, 1–16. [Google Scholar]
- Martin, F.P.J.; Wang, Y.L.; Sprenger, N.; Holmes, E.; Lindon, J.C.; Kochhar, S.; Nicholson, J.K. Effects of probiotic Lactobacillus Paracasei treatment on the host gut tissue metabolic profiles probed via magic-angle-spinning NMR spectroscopy. J. Proteome Res. 2007, 6, 1471–1481. [Google Scholar] [CrossRef]
- Forina, M.; Lanteri, S.; Oliveros, M.C.C.; Millan, C.P. Selection of useful predictors in multivariate calibration. Anal. Bioanal. Chem. 2004, 380, 397–418. [Google Scholar] [CrossRef]
- Huang, B.Y.; Kwock, L.; Castillo, M.; Smith, J.K. Association of choline levels and tumor perfusion in brain metastases assessed with proton MR spectroscopy and dynamic susceptibility contrast-enhanced perfusion weighted MRI. Technol. Cancer Res. Treat. 2010, 9, 327–337. [Google Scholar]
- Aboagye, E.O. Phosphatidylcholine metabolic transformation and progression signature as a pharmacodynamic biomarker. Oncotarget 2010, 1, 163–166. [Google Scholar]
- Schallreuter, K.U.; Hasse, S.; Rokos, H.; Chavan, B.; Shalbaf, M.; Spencer, J.D.; Wood, J.M. Cholesterol regulates melanogenesis in human epidermal melanocytes and melanoma cells. Exp. Dermatol. 2009, 18, 680–688. [Google Scholar] [CrossRef]
- Reynier, P.; Briane, D.; Cao, A.; Lievre, N.; Naejus, R.; Bissieres, P.; Salzmann, J.L.; Taillandier, E. In vitro and in vivo transfection of melanoma cells B16-F10 mediated by cholesterol-based cationic liposomes. J. Drug Target. 2002, 10, 557–566. [Google Scholar] [CrossRef]
- Jani, J.P.; Specht, S.; Stemmler, N.; Blanock, K.; Singh, S.V.; Gupta, V.; Katoh, A. Metastasis of B16f10 mouse melanoma inhibited by lovastatin, an inhibitor of cholesterol-biosynthesis. Invasion Metastasis 1993, 13, 314–324. [Google Scholar]
- McKnight, T.R.; Smith, K.J.; Chu, P.W.; Chiu, K.S.; Cloyd, C.P.; Chang, S.M.; Phillips, J.J.; Berger, M.S. Choline metabolism, proliferation, and angiogenesis in nonenhancing grades 2 and 3 astrocytoma. J. Magn. Reson. Imaging 2011, 33, 808–816. [Google Scholar] [CrossRef]
- Stewart, J.D.; Marchan, R.; Lesjak, M.S.; Lambert, J.; Hergenroeder, R.; Ellis, J.K.; Lau, C.H.; Keun, H.C.; Schmitz, G.; Schiller, J.; et al. Choline-releasing glycerophosphodiesterase EDI3 drives tumor cell migration and metastasis. Proc. Natl. Acad. Sci. USA 2012, 109, 8155–8160. [Google Scholar] [CrossRef]
- Moestue, S.A.; Borgan, E.; Huuse, E.M.; Lindholm, E.M.; Sitter, B.; Borresen-Dale, A.L.; Engebraaten, O.; Maelandsmo, G.M.; Gribbestad, I.S. Distinct choline metabolic profiles are associated with differences in gene expression for basal-like and luminal-like breast cancer xenograft models. BMC Cancer 2010, 10, 433. [Google Scholar] [CrossRef] [Green Version]
- Rocha, C.M.; Barros, A.S.; Gil, A.M.; Goodfellow, B.J.; Humpfer, E.; Spraul, M.; Carreira, I.M.; Melo, J.B.; Bernardo, J.; Gomes, A.; et al. Metabolic profiling of human lung cancer tissue by H-1 High Resolution Magic Angle Spinning (HRMAS) NMR spectroscopy. J. Proteome Res. 2010, 9, 319–332. [Google Scholar] [CrossRef]
- Patwardhan, G.A.; Liu, Y.Y. Sphingolipids and expression regulation of genes in cancer. Prog. Lipid Res. 2011, 50, 104–114. [Google Scholar] [CrossRef]
- Tajima, N.; Itokazu, Y.; Korpi, E.R.; Somerharju, P.; Kakela, R. Activity of BKCa channel is modulated by membrane cholesterol content and association with Na+/K+-ATPase in human melanoma IGR39 cells. J. Biol. Chem. 2011, 286, 5624–5638. [Google Scholar] [CrossRef]
- Benton, C.R.; Holloway, G.P.; Campbell, S.E.; Yoshida, Y.; Tandon, N.N.; Glatz, J.F.C.; Luiken, J.J.J.F.P.; Spriet, L.L.; Bonen, A. Rosiglitazone increases fatty acid oxidation and fatty acid translocase (FAT/CD36) but not carnitine palmitoyltransferase I in rat muscle mitochondria. J. Physiol. Lond. 2008, 586, 1755–1766. [Google Scholar] [CrossRef]
- Holloway, G.P.; Bezaire, V.; Heigenhauser, G.J.F.; Tandon, N.N.; Glatz, J.F.C.; Luiken, J.J.F.P.; Bonen, A.; Spriet, L.L. Mitochondrial long chain fatty acid oxidation, fatty acid translocase/CD36 content and carnitine palmitoyltransferase I activity in human skeletal muscle during aerobic exercise. J. Physiol. Lond. 2006, 571, 201–210. [Google Scholar] [CrossRef]
- Jakobs, B.S.; Wanders, R.J.A. Impaired peroxisomal fatty acid oxidation in human skin fibroblasts with a mitochondrial acylcarnitine carnitine translocase deficiency. J. Inherit. Metab. Dis. 1996, 19, 185–187. [Google Scholar] [CrossRef]
- Giordano, A.; Calvani, M.; Petillo, O.; Grippo, P.; Tuccillo, F.; Melone, M.A.B.; Bonelli, P.; Calarco, A.; Peluso, G. tBid induces alterations of mitochondrial fatty acid oxidation flux by malonyl-CoA-independent inhibition of carnitine palmitoyltransferase-1. Cell Death Differ. 2005, 12, 603–613. [Google Scholar] [CrossRef]
- Thupari, J.N.; Pinn, M.L.; Kuhajda, F.P. Fatty acid synthase inhibition in human breast cancer cells leads to malonyl-CoA-induced inhibition of fatty acid oxidation and cytotoxicity. Biochem. Biophys. Res. Commun. 2001, 285, 217–223. [Google Scholar] [CrossRef]
- Kroemer, G.; Pouyssegur, J. Tumor cell metabolism: Cancer’s Achilles’ heel. Cancer Cell 2008, 13, 472–482. [Google Scholar] [CrossRef]
- Visarius, T.M.; Bahler, H.; Kupfer, A.; Cerny, T.; Lauterburg, B.H. Thiodiglycolic acid is excreted by humans receiving ifosfamide and inhibits mitochondrial function in rats. Drug Metab. Dispos. 1998, 26, 193–196. [Google Scholar]
- Yokota, H.; Guo, J.F.; Matoba, M.; Higashi, K.; Tonami, H.; Nagao, Y. Lactate, choline, and creatine levels measured by vitro H-1-MRS as prognostic parameters in patients with non-small-cell lung cancer. J. Magn. Reson. Imaging 2007, 25, 992–999. [Google Scholar] [CrossRef]
- Howe, F.A.; Barton, S.J.; Cudlip, S.A.; Stubbs, M.; Saunders, D.E.; Murphy, M.; Wilkins, P.; Opstad, K.S.; Doyle, V.L.; McLean, M.A.; et al. Metabolic profiles of human brain tumors using quantitative in vivo H-1 magnetic resonance spectroscopy. Magn. Reson. Med. 2003, 49, 223–232. [Google Scholar] [CrossRef]
- Marchan, R.; Lesjak, M.S.; Stewart, J.D.; Winter, R.; Seeliger, J.; Hengstler, J.G. Choline-releasing glycerophosphodiesterase EDI3 links the tumor metabolome to signaling network activities. Cell Cycle 2012, 11, 4499–4506. [Google Scholar] [CrossRef]
- Moestue, S.A.; Giskeodegard, G.F.; Cao, M.D.; Bathen, T.F.; Gribbestad, I.S. Glycerophosphocholine (GPC) is a poorly understood biomarker in breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, E2506–E2506. [Google Scholar]
- Miller, B.L. A review of chemical issues in 1H-NMR spectroscopy—N-acetyl-L-aspartate, creatine and choline. NMR Biomed. 1991, 4, 47–52. [Google Scholar] [CrossRef]
- Yu, J.; Kim, A.K. Effect of taurine on antioxidant enzyme system in B16F10 melanoma cells. Adv. Exp. Med. Biol. 2009, 643, 491–499. [Google Scholar] [CrossRef]
- Morvan, D.; Demidem, A. Metabolomics by proton nuclear magnetic resonance spectroscopy of the response to chloroethylnitrosourea reveals drug efficacy and tumor adaptive metabolic pathways. Cancer Res. 2007, 67, 2150–2159. [Google Scholar] [CrossRef]
- Pollock, P.M.; Cohen-Solal, K.; Sood, R.; Namkoong, J.; Martino, J.J.; Koganti, A.; Zhu, H.; Robbins, C.; Makalowska, I.; Shin, S.S.; et al. Melanoma mouse model implicates metabotropic glutamate signaling in melanocytic neoplasia. Nat. Genet. 2003, 34, 108–112. [Google Scholar] [CrossRef]
- Le, M.N.; Chan, J.L.K.; Rosenberg, S.A.; Nabatian, A.S.; Merrigan, K.T.; Cohen-Solal, K.A.; Goydos, J.S. The glutamate release inhibitor Riluzole decreases migration, invasion, and proliferation of melanoma cells. J. Investig. Dermatol. 2010, 130, 2240–2249. [Google Scholar] [CrossRef]
- Rustin, P. Mitochondria, from cell death to proliferation. Nat. Genet. 2002, 30, 352–353. [Google Scholar] [CrossRef]
- Selak, M.A.; Armour, S.M.; MacKenzie, E.D.; Boulahbel, H.; Watson, D.G.; Mansfield, K.D.; Pan, Y.; Simon, M.C.; Thompson, C.B.; Gottlieb, E. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell 2005, 7, 77–85. [Google Scholar] [CrossRef]
- Tiziani, S.; Lodi, A.; Khanim, F.L.; Viant, M.R.; Bunce, C.M.; Gunther, U.L. Metabolomic profiling of drug responses in acute myeloid leukaemia cell lines. PLoS One 2009, 4, e4251. [Google Scholar]
- Burrows, N.; Babur, M.; Resch, J.; Williams, K.J.; Brabant, G. Hypoxia-inducible factor in thyroid carcinoma. J. Thyroid Res. 2011, 2011. Article ID 762905. [Google Scholar]
- Jones, N.; Kim, P.J.; Agani, F. HIF-1 deficiency promotes hypoxia-induced cell death in melanoma cells. FASEB J. 2004, 18, A726–A727. [Google Scholar]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Feng, J.; Isern, N.G.; Burton, S.D.; Hu, J.Z. Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics. Metabolites 2013, 3, 1011-1035. https://doi.org/10.3390/metabo3041011
Feng J, Isern NG, Burton SD, Hu JZ. Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics. Metabolites. 2013; 3(4):1011-1035. https://doi.org/10.3390/metabo3041011
Chicago/Turabian StyleFeng, Ju, Nancy G. Isern, Sarah D. Burton, and Jian Zhi Hu. 2013. "Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics" Metabolites 3, no. 4: 1011-1035. https://doi.org/10.3390/metabo3041011
APA StyleFeng, J., Isern, N. G., Burton, S. D., & Hu, J. Z. (2013). Studies of Secondary Melanoma on C57BL/6J Mouse Liver Using 1H NMR Metabolomics. Metabolites, 3(4), 1011-1035. https://doi.org/10.3390/metabo3041011