Stable Isotope Tracer Analysis in Isolated Mitochondria from Mammalian Systems

Abstract
:1. Introduction
2. Results and Discussion
2.1. Method Rationale and Assessment of Mitochondrial Functions

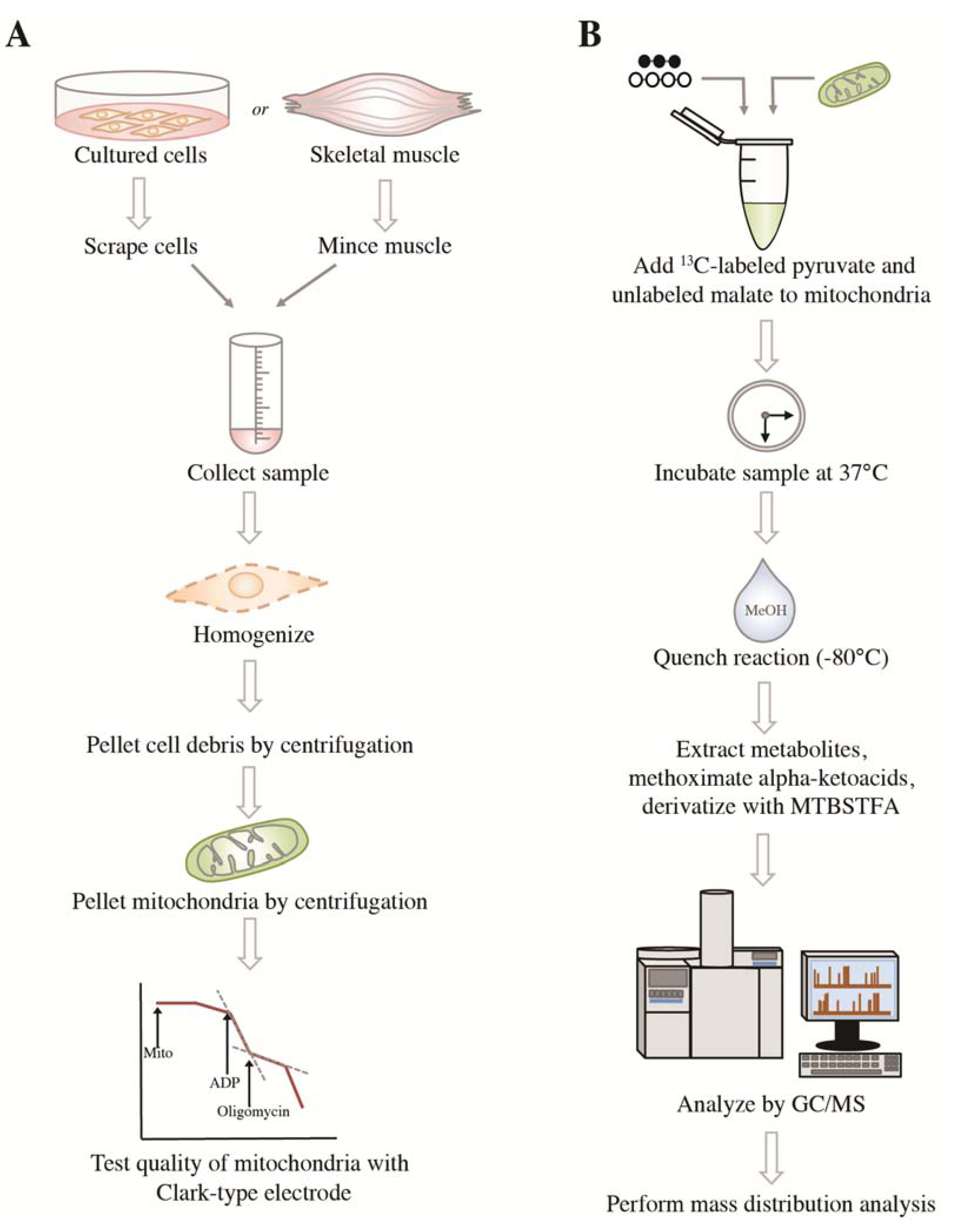{kind=link}
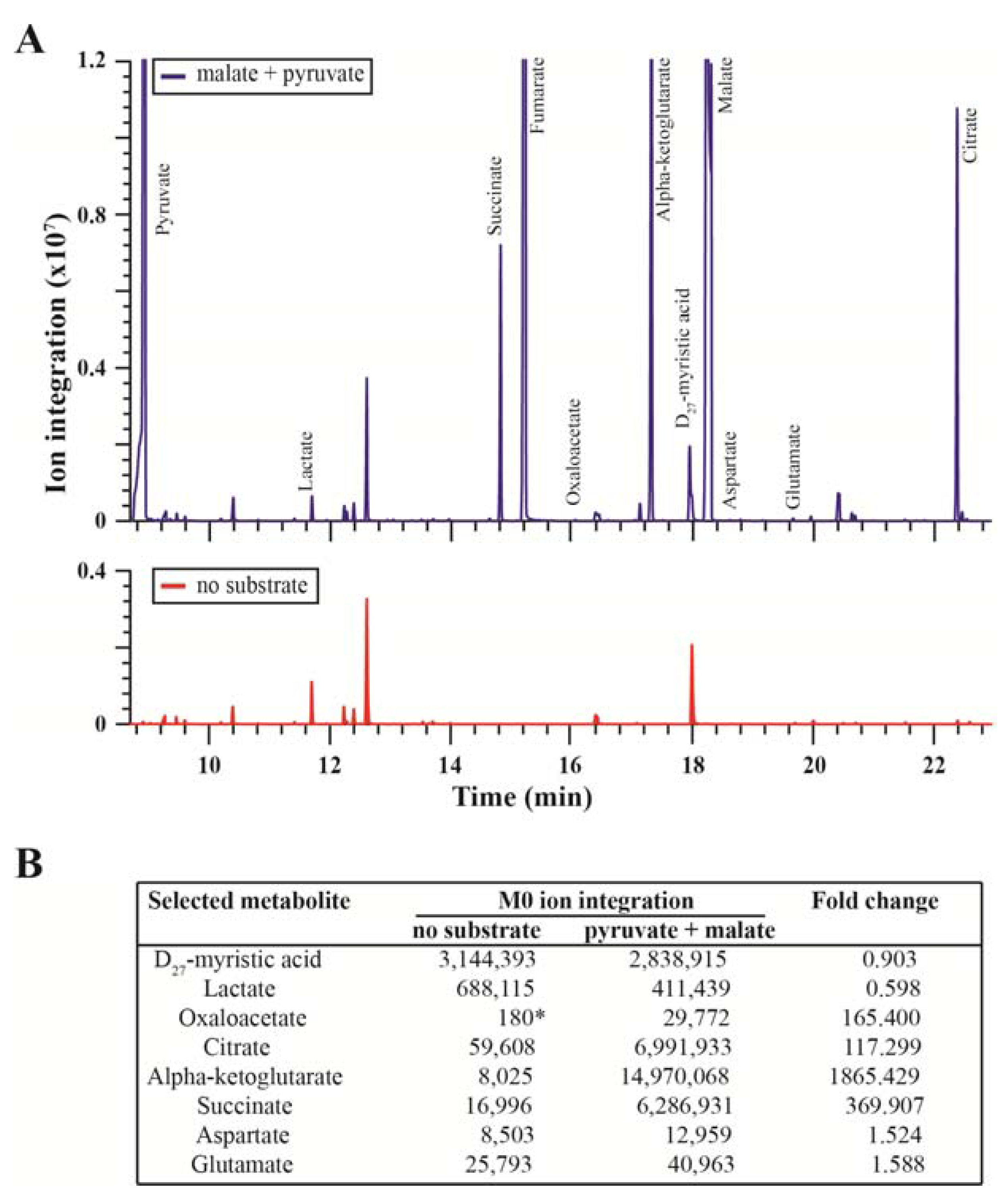{kind=link}
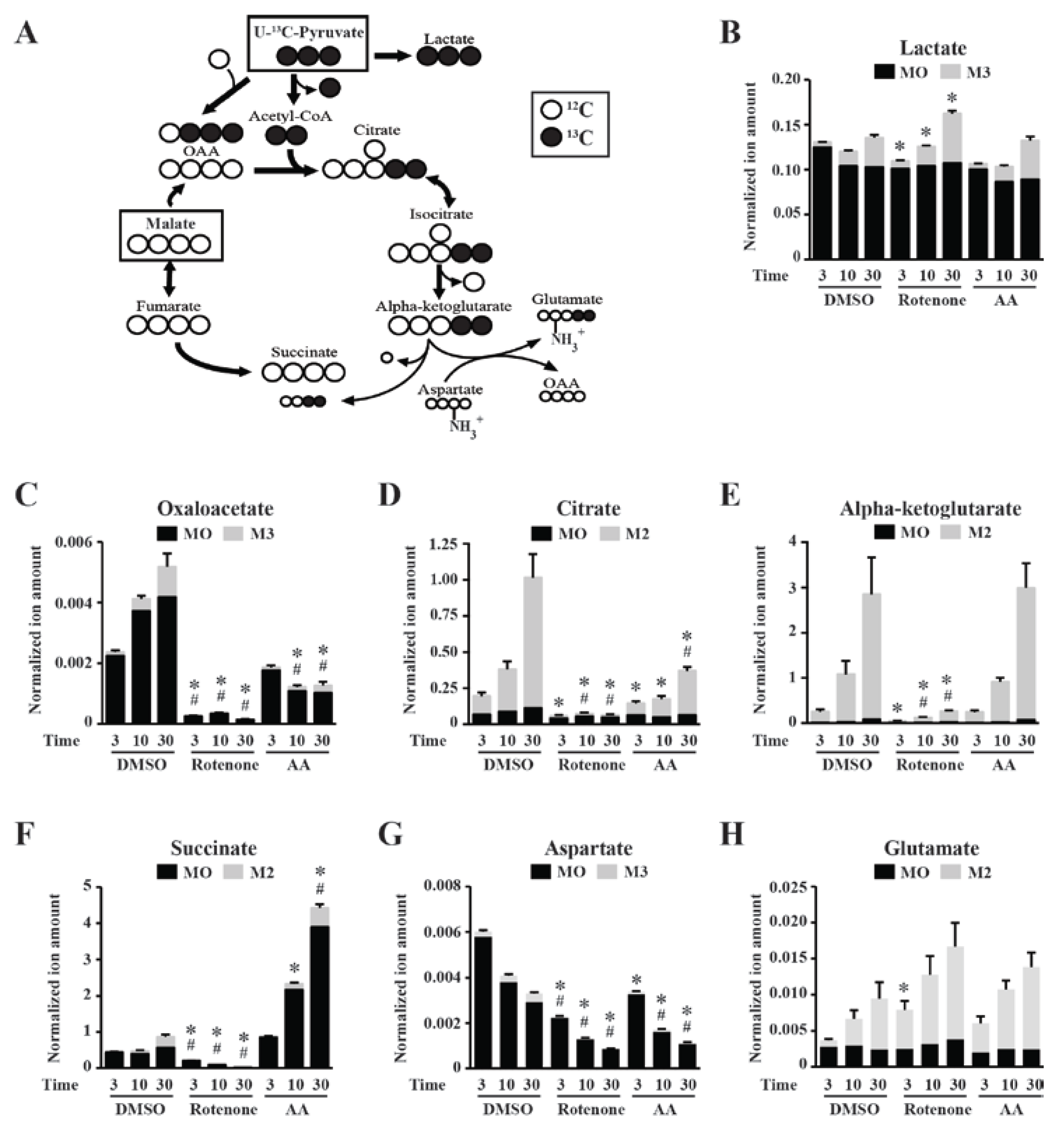{kind=link}
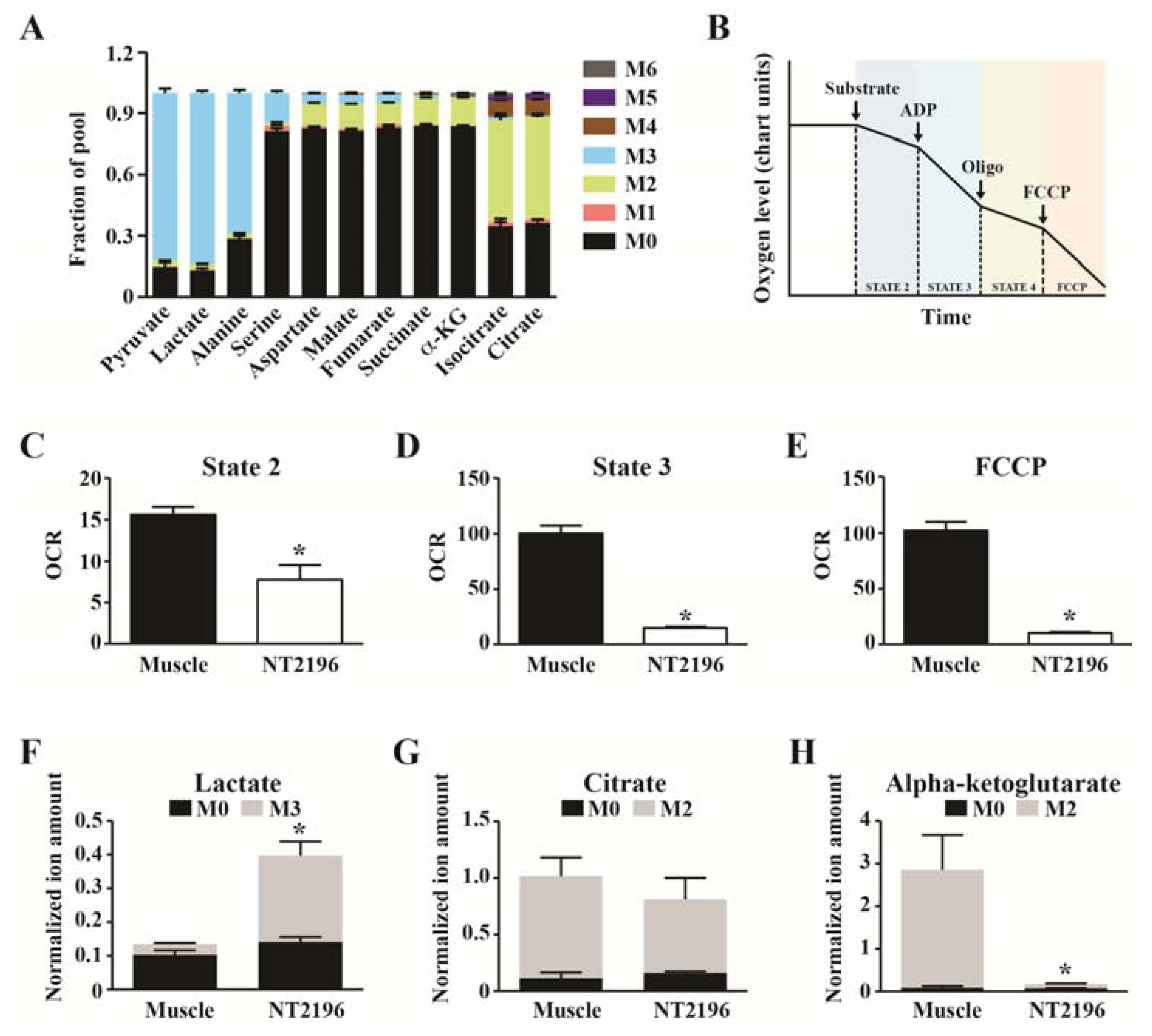{kind=link}
| Source | RCR | 13C-substrate(s) | Incubation time | Analyzed Metabolites | Platform | Reference |
|---|---|---|---|---|---|---|
| Rat brain | ³5 | U-13C-glutamate | 4 min | KG, Succ, Mal | NMR | [22] |
| Potato tubers | N/A | Multiple forms of 13C-glutamate | 0–11 h | KG, Gln, Mal, Asp | NMR | [23] |
| Arabidopsis cells | N/A | 1-13C-glucose | 16 h | Glycolytic, Cit, Succ, Fum, Mal and others | GC/MS | [24] |
| 1-13C-fructose 1,6-bisphosphate | ||||||
| Arabidopsis cells | N/A | 1-13C-glucose | 0–16 h | Glycolytic, Cit, Succ, Mal | NMR | [25] |
| 1-13C-fructose 1,6-bisphosphate | ||||||
| Rat brain | 3.0–4.5 | U-13C-glutamate | 5 min | Glu, Gln, Asp | LC/MS | [26] |
| U-13C-glutamine | ||||||
| U-13C-succinate |

2.2. Stable Isotope Tracer Analyses of Skeletal Muscle Mitochondria Treated with Electron Transport Chain (ETC) Inhibitors


2.3. Stable Isotope Tracer Analyses in Mitochondria Isolated from Cancer Cells

3. Experimental Section
3.1. Animals, Cells and Reagents
3.2. Isolation of Mitochondria from Mice
3.3. Isolation of Mitochondria from Cultured Cells
3.4. Respiration
3.5. Extraction of Metabolites from Cells and Isolated Mitochondria
3.6. GC/MS
3.7. Mass Isotopomer Distribution Analysis
| Metabolite | Molecular formula | m/z
quantifying ions (qualifyer) | Retention time
(min) |
|---|---|---|---|
| Pyruvate oxime | C6H12NO3Si | 174–177 (115) | 8.978 |
| Lactate | C11H25O3Si2 | 261–264 (233) | 11.742 |
| Alanine | C11H26NO2Si2 | 260–263 (232) | 12.312 |
| Succinate | C12H25O4Si2 | 289–293 (331) | 14.813 |
| Fumarate | C12H23O4Si2 | 287–291 (329) | 15.208 |
| Oxaloacetate oxime | C13H26NO5Si2 | 332–336 (374) | 16.068 |
| Serine | C17H40NO3Si3 | 390–393 (362) | 17.108 |
| Alpha-ketoglutarate oxime | C14H28NO5Si2 | 346–351 (258) | 17.319 |
| Malate | C18H39O5Si3 | 419–423 (287) | 18.275 |
| Aspartate | C18H40NO4Si3 | 418–422 (390) | 18.638 |
| Glutamate | C19H42NO4Si3 | 432–437 (330) | 19.685 |
| Citrate | C20H39O6Si3 | 459–465 (431,403) | 22.429 |
| Isocitrate | C20H39O6Si3 | 459–465 (431,403) | 22.518 |
| D27-myristic acid | C16H6D27O2Si | 312 (132) | 17.912 |
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gutierrez-Aguilar, M.; Baines, C.P. Physiological and pathological roles of mitochondrial slc25 carriers. Biochem. J. 2013, 454, 371–386. [Google Scholar] [CrossRef]
- Austin, S.; St-Pierre, J. Pgc1alpha and mitochondrial metabolism—Emerging concepts and relevance in ageing and neurodegenerative disorders. J. Cell Sci. 2012, 125, 4963–4971. [Google Scholar] [CrossRef]
- Vazquez, F.; Lim, J.H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; Clish, C.B.; Granter, S.R.; Widlund, H.R.; Spiegelman, B.M.; et al. Pgc1alpha expression defines a subset of human melanoma tumors with increased mitochondrial capacity and resistance to oxidative stress. Cancer Cell 2013, 23, 287–301. [Google Scholar] [CrossRef]
- Tennakoon, J.B.; Shi, Y.; Han, J.J.; Tsouko, E.; White, M.A.; Burns, A.R.; Zhang, A.; Xia, X.; Ilkayeva, O.R.; Xin, L.; et al. Androgens regulate prostate cancer cell growth via an ampk-pgc-1alpha-mediated metabolic switch. Oncogene 2013. [Google Scholar] [CrossRef]
- Fan, T.W.; Yuan, P.; Lane, A.N.; Higashi, R.M.; Wang, Y.; Hamidi, A.B.; Zhou, R.; Guitart, X.; Chen, G.; Manji, H.K.; et al. Stable isotope-resolved metabolomic analysis of lithium effects on glial-neuronal metabolism and interactions. Metabolomics 2010, 6, 165–179. [Google Scholar] [CrossRef]
- Fan, T.W.; Lane, A.N.; Higashi, R.M.; Yan, J. Stable isotope resolved metabolomics of lung cancer in a scid mouse model. Metabolomics 2011, 7, 257–269. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W.; Bousamra, M.; Higashi, R.M.; Yan, J.; Miller, D.M. Stable isotope-resolved metabolomics (sirm) in cancer research with clinical application to nonsmall cell lung cancer. OMICS. 2011, 15, 173–182. [Google Scholar] [CrossRef]
- Mullen, A.R.; Wheaton, W.W.; Jin, E.S.; Chen, P.H.; Sullivan, L.B.; Cheng, T.; Yang, Y.; Linehan, W.M.; Chandel, N.S.; DeBerardinis, R.J. Reductive carboxylation supports growth in tumour cells with defective mitochondria. Nature 2012, 481, 385–388. [Google Scholar]
- Frezza, C.; Zheng, L.; Folger, O.; Rajagopalan, K.N.; MacKenzie, E.D.; Jerby, L.; Micaroni, M.; Chaneton, B.; Adam, J.; Hedley, A.; et al. Haem oxygenase is synthetically lethal with the tumour suppressor fumarate hydratase. Nature 2011, 477, 225–228. [Google Scholar] [CrossRef]
- Pan, Z.; Raftery, D. Comparing and combining nmr spectroscopy and mass spectrometry in metabolomics. Anal. Bioanal. Chem. 2007, 387, 525–527. [Google Scholar] [CrossRef]
- Keshari, K.R.; Kurhanewicz, J.; Jeffries, R.E.; Wilson, D.M.; Dewar, B.J.; van Criekinge, M.; Zierhut, M.; Vigneron, D.B.; Macdonald, J.M. Hyperpolarized 13c spectroscopy and an NMR-compatible bioreactor system for the investigation of real-time cellular metabolism. Magn. Reson. Med. 2010, 63, 322–329. [Google Scholar] [CrossRef]
- Kurhanewicz, J.; Vigneron, D.B.; Brindle, K.; Chekmenev, E.Y.; Comment, A.; Cunningham, C.H.; Deberardinis, R.J.; Green, G.G.; Leach, M.O.; Rajan, S.S.; et al. Analysis of cancer metabolism by imaging hyperpolarized nuclei: Prospects for translation to clinical research. Neoplasia 2011, 13, 81–97. [Google Scholar]
- Merritt, M.E.; Harrison, C.; Storey, C.; Jeffrey, F.M.; Sherry, A.D.; Malloy, C.R. Hyperpolarized 13c allows a direct measure of flux through a single enzyme-catalyzed step by nmr. Proc. Natl. Acad. Sci. USA 2007, 104, 19773–19777. [Google Scholar] [CrossRef]
- Mamer, O.; Gravel, S.P.; Choiniere, L.; Chenard, V.; St-Pierre, J.; Avizonis, D. The complete targeted profile of the organic acid intermediates of the citric acid cycle using a single stable isotope dilution analysis, sodium borodeuteride reduction and selected ion monitoring GC/MS. Metabolomics 2013, 9, 1019–1030. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via uplc-ms. Nat.Protoc. 2013, 8, 17–32. [Google Scholar]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PloS One 2013, 8, e73076. [Google Scholar] [CrossRef]
- Kujala, U.M.; Makinen, V.P.; Heinonen, I.; Soininen, P.; Kangas, A.J.; Leskinen, T.H.; Rahkila, P.; Wurtz, P.; Kovanen, V.; Cheng, S.; et al. Long-term leisure-time physical activity and serum metabolome. Circulation 2013, 127, 340–348. [Google Scholar] [CrossRef]
- Dallmann, R.; Viola, A.U.; Tarokh, L.; Cajochen, C.; Brown, S.A. The human circadian metabolome. Proc. Natl. Acad. Sci. USA 2012, 109, 2625–2629. [Google Scholar] [CrossRef]
- Guo, A.C.; Jewison, T.; Wilson, M.; Liu, Y.; Knox, C.; Djoumbou, Y.; Lo, P.; Mandal, R.; Krishnamurthy, R.; Wishart, D.S. Ecmdb: The E. coli metabolome database. Nucleic Acids Res. 2013, 41, D625–D630. [Google Scholar] [CrossRef]
- Jewison, T.; Knox, C.; Neveu, V.; Djoumbou, Y.; Guo, A.C.; Lee, J.; Liu, P.; Mandal, R.; Krishnamurthy, R.; Sinelnikov, I.; et al. Ymdb: The yeast metabolome database. Nucleic Acids Res. 2012, 40, D815–D820. [Google Scholar] [CrossRef]
- Reinke, S.N.; Hu, X.; Sykes, B.D.; Lemire, B.D. Caenorhabditis elegans diet significantly affects metabolic profile, mitochondrial DNA levels, lifespan and brood size. Mol. Genet. Metab. 2010, 100, 274–282. [Google Scholar] [CrossRef]
- Bakken, I.J.; Sonnewald, U.; Clark, J.B.; Bates, T.E. [U-13C]glutamate metabolism in rat brain mitochondria reveals malic enzyme activity. Neuroreport 1997, 8, 1567–1570. [Google Scholar] [CrossRef]
- Aubert, S.; Bligny, R.; Douce, R.; Gout, E.; Ratcliffe, R.G.; Roberts, J.K. Contribution of glutamate dehydrogenase to mitochondrial glutamate metabolism studied by 13C and 31P nuclear magnetic resonance. J. Exp. Bot. 2001, 52, 37–45. [Google Scholar] [CrossRef]
- Giege, P.; Heazlewood, J.L.; Roessner-Tunali, U.; Millar, A.H.; Fernie, A.R.; Leaver, C.J.; Sweetlove, L.J. Enzymes of glycolysis are functionally associated with the mitochondrion in arabidopsis cells. Plant Cell 2003, 15, 2140–2151. [Google Scholar] [CrossRef]
- Graham, J.W.; Williams, T.C.; Morgan, M.; Fernie, A.R.; Ratcliffe, R.G.; Sweetlove, L.J. Glycolytic enzymes associate dynamically with mitochondria in response to respiratory demand and support substrate channeling. Plant Cell 2007, 19, 3723–3738. [Google Scholar] [CrossRef]
- Bak, L.K.; Zieminska, E.; Waagepetersen, H.S.; Schousboe, A.; Albrecht, J. Metabolism of [U-13C]glutamine and [U-13C]glutamate in isolated rat brain mitochondria suggests functional phosphate-activated glutaminase activity in matrix. Neurochem. Res. 2008, 33, 273–278. [Google Scholar]
- Gregersen, N.; Hansen, J.; Palmfeldt, J. Mitochondrial proteomics—A tool for the study of metabolic disorders. J. Inherit. Metab. Dis. 2012, 35, 715–726. [Google Scholar] [CrossRef]
- Angelini, R.; Vitale, R.; Patil, V.A.; Cocco, T.; Ludwig, B.; Greenberg, M.L.; Corcelli, A. Lipidomics of intact mitochondria by maldi-tof/ms. J. Lipid Res. 2012, 53, 1417–1425. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Fahy, E.; Guan, Z.; Kelly, S.; Li, X.; McDonald, J.G.; Milne, S.; Myers, D.; Park, H.; Ryan, A.; et al. Subcellular organelle lipidomics in tlr-4-activated macrophages. J. Lipid Res. 2010, 51, 2785–2797. [Google Scholar] [CrossRef]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef]
- Kombu, R.S.; Brunengraber, H.; Puchowicz, M.A. Analysis of the citric acid cycle intermediates using gas chromatography-mass spectrometry. Methods Mol. Bio. 2011, 708, 147–157. [Google Scholar] [CrossRef]
- St-Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002, 277, 44784–44790. [Google Scholar] [CrossRef]
- Chance, B.; Williams, G.R. Respiratory enzymes in oxidative phosphorylation. I. Kinetics of oxygen utilization. J. Biol. Chem. 1955, 217, 383–393. [Google Scholar]
- Chance, B.; Williams, G.R.; Holmes, W.F.; Higgins, J. Respiratory enzymes in oxidative phosphorylation. V. A mechanism for oxidative phosphorylation. J. Biol. Chem. 1955, 217, 439–451. [Google Scholar]
- Fendt, S.M.; Bell, E.L.; Keibler, M.A.; Olenchock, B.A.; Mayers, J.R.; Wasylenko, T.M.; Vokes, N.I.; Guarente, L.; Vander Heiden, M.G.; Stephanopoulos, G. Reductive glutamine metabolism is a function of the alpha-ketoglutarate to citrate ratio in cells. Nat. Commun. 2013, 4, 2236. [Google Scholar]
- Xu, Q.W.; Vu, H.; Liu, L.P.; Wang, T.C.; Schaefer, W.H. Metabolic profiles show specific mitochondrial toxicities in vitro in myotube cells. J.Biomol.Nmr. 2011, 49, 207–219. [Google Scholar] [CrossRef]
- Metallo, C.M.; Gameiro, P.A.; Bell, E.L.; Mattaini, K.R.; Yang, J.; Hiller, K.; Jewell, C.M.; Johnson, Z.R.; Irvine, D.J.; Guarente, L.; et al. Reductive glutamine metabolism by idh1 mediates lipogenesis under hypoxia. Nature 2012, 481, 380–384. [Google Scholar]
- Minet, A.D.; Gaster, M. Pyruvate carboxylase is expressed in human skeletal muscle. Biochem. Bioph. Res. Co. 2010, 402, 196–197. [Google Scholar] [CrossRef]
- Elustondo, P.A.; White, A.E.; Hughes, M.E.; Brebner, K.; Pavlov, E.; Kane, D.A. Physical and functional association of lactate dehydrogenase (ldh) with skeletal muscle mitochondria. J. Biol. Chem. 2013, 288, 25309–25317. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of ldh-a expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef]
- Klimcakova, E.; Chenard, V.; McGuirk, S.; Germain, D.; Avizonis, D.; Muller, W.J.; St-Pierre, J. Pgc-1alpha promotes the growth of erbb2/neu-induced mammary tumors by regulating nutrient supply. Cancer Res. 2012, 72, 1538–1546. [Google Scholar] [CrossRef]
- Ursini-Siegel, J.; Rajput, A.B.; Lu, H.; Sanguin-Gendreau, V.; Zuo, D.; Papavasiliou, V.; Lavoie, C.; Turpin, J.; Cianflone, K.; Huntsman, D.G.; et al. Elevated expression of decr1 impairs erbb2/neu-induced mammary tumor development. Mol. Cell. Biol. 2007, 27, 6361–6371. [Google Scholar] [CrossRef]
- McGuirk, S.; Gravel, S.P.; Deblois, G.; Papadopoli, D.J.; Faubert, B.; Wegner, A.; Hiller, K.; Avizonis, D.; Akavia, U.D.; Jones, R.G.; et al. Pgc-1alpha supports glutamine metabolism in breast cancer. Cancer Metab. 2013, 1, 22. [Google Scholar] [CrossRef]
- Danial, N.N.; Gramm, C.F.; Scorrano, L.; Zhang, C.Y.; Krauss, S.; Ranger, A.M.; Datta, S.R.; Greenberg, M.E.; Licklider, L.J.; Lowell, B.B.; et al. Bad and glucokinase reside in a mitochondrial complex that integrates glycolysis and apoptosis. Nature 2003, 424, 952–956. [Google Scholar] [CrossRef]
- Austin, S.; Klimcakova, E.; St-Pierre, J. Impact of pgc-1alpha on the topology and rate of superoxide production by the mitochondrial electron transport chain. Free Radical Bio. Med. 2011, 51, 2243–2248. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef]
- Morita, M.; Gravel, S.P.; Chenard, V.; Sikstrom, K.; Zheng, L.; Alain, T.; Gandin, V.; Avizonis, D.; Arguello, M.; Zakaria, C.; et al. Mtorc1 controls mitochondrial activity and biogenesis through 4e-bp-dependent translational regulation. Cell Metab. 2013, 18, 698–711. [Google Scholar] [CrossRef]
- Nanchen, A.; Fuhrer, T.; Sauer, U. Determination of metabolic flux ratios from 13c-experiments and gas chromatography-mass spectrometry data: Protocol and principles. Methods Mol. Bio. 2007, 358, 177–197. [Google Scholar] [CrossRef]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Cao, Q.; Yu, J.; Laxman, B.; Mehra, R.; Lonigro, R.J.; Li, Y.; et al. Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 2009, 457, 910–914. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gravel, S.-P.; Andrzejewski, S.; Avizonis, D.; St-Pierre, J. Stable Isotope Tracer Analysis in Isolated Mitochondria from Mammalian Systems. Metabolites 2014, 4, 166-183. https://doi.org/10.3390/metabo4020166
Gravel S-P, Andrzejewski S, Avizonis D, St-Pierre J. Stable Isotope Tracer Analysis in Isolated Mitochondria from Mammalian Systems. Metabolites. 2014; 4(2):166-183. https://doi.org/10.3390/metabo4020166
Chicago/Turabian StyleGravel, Simon-Pierre, Sylvia Andrzejewski, Daina Avizonis, and Julie St-Pierre. 2014. "Stable Isotope Tracer Analysis in Isolated Mitochondria from Mammalian Systems" Metabolites 4, no. 2: 166-183. https://doi.org/10.3390/metabo4020166
APA StyleGravel, S. -P., Andrzejewski, S., Avizonis, D., & St-Pierre, J. (2014). Stable Isotope Tracer Analysis in Isolated Mitochondria from Mammalian Systems. Metabolites, 4(2), 166-183. https://doi.org/10.3390/metabo4020166