Xanthan Gum Removal for 1H-NMR Analysis of the Intracellular Metabolome of the Bacteria Xanthomonas axonopodis pv. citri 306

Abstract
:
1. Introduction
2. Results and Discussion
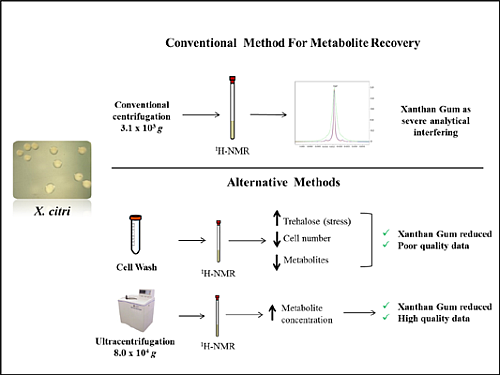
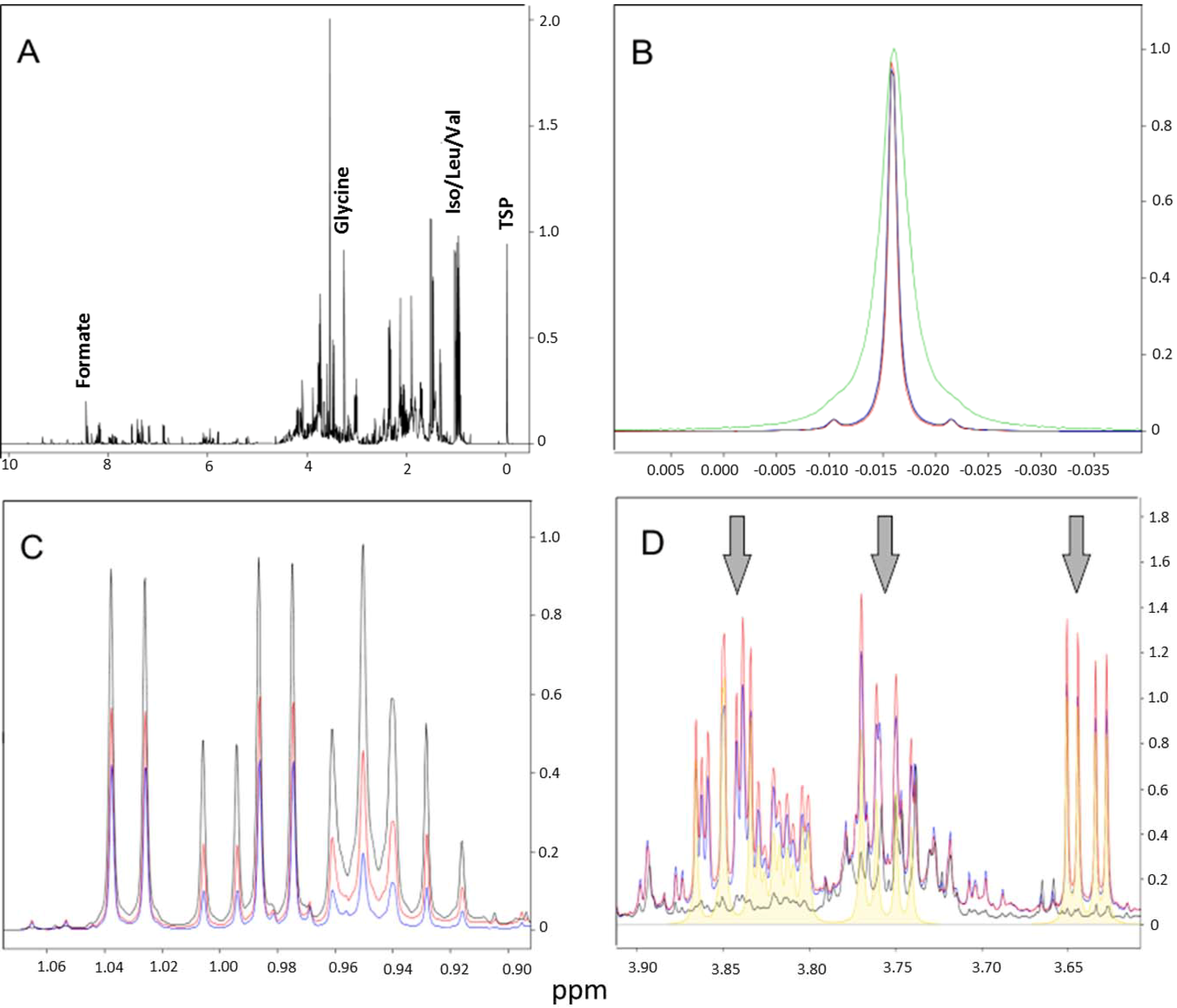
2.1. Selection of the Best Intervention Procedure for Xanthan Gum Removal

2.1.1. Cell-Washing Protocols Lead to a Marked Decrease in Overall Intracellular Metabolites’ Retrieval, but One in Particular Followed the Opposite Trend


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Ultracentrifugation | 3 washing cycles | 5 washing cycles |
|---|---|---|---|
| 2-Aminoadipate | 1,659.9 | 1,638.5 | 1,608.5 |
| 2-Methylglutarate | 21.6 | 57.7 | 44.3 |
| 3-Hydroxybutyrate | 186.2 | 992.4 | 515.5 |
| 3-Hydroxyisovalerate | 12.3 | 9.0 | 5.4 |
| 4-Aminohippurate | 48.1 | 20.9 | 24.7 |
| AMP | 24.2 | 36.9 | 29.6 |
| ATP | 42.7 | 54.9 | 56.9 |
| Acetate | 1,044.2 | 941.1 | 842.0 |
| Adenine | 39.2 | 35.6 | 33.1 |
| Adenosine | 129.1 | 305.2 | 282.9 |
| Alanine | 2,817.5 | 3,519.6 | 1,996.2 |
| Asparagine | 171.6 | 260.3 | 153.1 |
| Aspartate | 805.9 | 1,175.2 | 997.6 |
| Beta-Alanine | 497.8 | 358.2 | 313.6 |
| Betaine | 741.9 | 885.4 | 789,0 |
| Cytidine | 199.1 | 247.0 | 183.6 |
| Dimethylamine | 15.4 | 21.0 | 22.4 |
| Dimethylformamide | 2.2 | 6.9 | N.Q. |
| dTTP | 105.6 | 64.7 | 63.0 |
| Formate | 1,118.5 | 1,201.3 | 1,387.3 |
| Fumarate | 70.0 | 11.3 | 17.6 |
| GTP | 51.7 | 96.2 | 119.9 |
| Glucose | 902.2 | 4,351.7 | 3,225.6 |
| Glutamate | 9,596.4 | 9,021.5 | 6,871.9 |
| Glutamine | 2,134.1 | 1,336.5 | 1,277.5 |
| Glycerol | 283.6 | 408.9 | N.Q. |
| Glycine | 4,289.0 | 2,848.7 | 2,511.9 |
| Guanosine | 128.2 | 97.8 | 66.4 |
| Hypoxanthine | 718.7 | 339.8 | 287.4 |
| Inosine | 200.0 | 108.7 | 92.2 |
| Isoleucine | 1,851.1 | 841.9 | 365.1 |
| Isovalerate | 61.6 | 37.0 | 11.0 |
| Lactate | 159.8 | 154.5 | 125.4 |
| Leucine | 3,041.4 | 1,492.0 | 564.5 |
| Lysine | 2,699.0 | 1386.3 | 1,019.9 |
| Methionine | 878.7 | 539.0 | 282.3 |
| Methylhistidine | 68.8 | 11.1 | 0 |
| NAD+ | 498.2 | 476.7 | 484.3 |
| NADP+ | 60.3 | 60.7 | 66.4 |
| Niacinamide | 84.2 | 43.1 | 28.0 |
| Phenylalanine | 1,218.1 | 376.4 | 76.4 |
| Proline | 1,453.1 | 1,693.5 | 1,139.6 |
| Pyroglutamate | 612.1 | 451.2 | 154.5 |
| Succinate | 89.0 | 897.2 | 864.3 |
| Threonine | 1,204.5 | 1,286.8 | 663.5 |
| Thymidine | 29.2 | 23.9 | 19.5 |
| Trehalose | 306.6 | 9,496.0 | 11,356.6 |
| Trimethylamine | 13.0 | 12.0 | 14.0 |
| Tryptophan | 297.6 | 149.1 | 77.9 |
| Tyrosine | 782.2 | 496.1 | 353.6 |
| UDP-Acetylglucosamine | 80.2 | 63.7 | 50.6 |
| UDP-galactose | 66.2 | 54.8 | 36.8 |
| UDP-glucose | 114.0 | 75.7 | 69.3 |
| UDP-glucuronate | 25.8 | 21.0 | 20.0 |
| Uracil | 673.0 | 203.1 | 233.3 |
| Uridine | 224.5 | 147.0 | 108.3 |
| Urocanate | 0 | 4.0 | 0 |
| Valine | 3,673.5 | 2,361.9 | 1,628.9 |
| Xanthosine | 4.9 | 6.5 | 7.8 |
| SCORE | 35 | 17 | 7 |
2.1.2. Decrease in Overall Metabolites’ Concentration in Cell-Washing Protocols is Due to Cell Loss
| Dilution | Unwashed | 3 washing cycles | 5 washing cycles |
|---|---|---|---|
| 1:103 | ID | ID | ID |
| 1:104 | ID | 800/816 | 656/680 |
| 1:105 | ID | 146/190 | 126/156 |
| 1:106 | 664/672 | 17/24 | 30/46 |
| 1:107 | 480/696 | 0/1 | 6/6 |
2.2. Lower Amounts of Cells for Metabolomic Studies by 1H-NMR
| Pellet volumes | |||
|---|---|---|---|
| Metabolites | 500 μL | 250 μL | 125 μL |
| 2-Aminoadipate | 653.9 ± 12.6 | 305.9 ± 33.2 | 148.6 ± 13.1 |
| 2-Methylglutarate | 55.9 ± 2.5 | 21.9 ± 7.5 | 7.0 ± 0.2 |
| 3-Hydroxybutyrate | 235.5 ± 22.7 | 115.2 ± 12.9 | 52.4 ± 4.4 |
| 3-Hydroxyisovalerate | 116.8 ± 18.2 | 73.3 ± 5.9 | 30.4 ± 5.0 |
| 4-Aminohippurate | 84.6 ± 4.9 | 30.2 ± 9.1 | 11.9 ± 1.2 |
| AMP | 1.9 ± 0.4 | 7.1 ± 4.3 | 12.7 ± 4.0 |
| ATP | 75.3 ± 3.5 | 32.1 ± 5.7 | 17.5 ± 3.3 |
| Acetate | 914.3 ± 173.7 | 470.7 ± 116.8 | 213.8 ± 26.7 |
| Adenine | 124.7 ± 8.5 | 58.0 ± 13.3 | 29.4 ± 4.4 |
| Adenosine | 268.6 ± 49.5 | 121.8 ± 20.8 | 49.6 ± 5.1 |
| Alanine | 4,475.6 ± 226.7 | 1,999.3 ± 383.6 | 1,042.9 ± 89.6 |
| Asparagine | 188.2 ± 7 | 90.3 ± 22.6 | 51.2 ± 8.1 |
| Aspartate | 1,579.5 ± 145.9 | 738.3 ± 100.8 | 368.0 ± 34.6 |
| Beta-Alanine | 288.3 ± 30.4 | 166.3 ± 16.1 | 79.3 ± 4.6 |
| Betaine | 842.9 ± 31.0 | 377.6 ± 52.3 | 188.7 ± 12.4 |
| Cytidine | 196.8 ± 19.7 | 54.4 ± 18.3 | 25.8 ± 2.1 |
| Dimethylamine | 3.2 ± 0.8 | 2.3 ± 0.2 | 1.0 ± 0.1 |
| Dimethylformamide | 26.1 ± 4.5 | 11.5 ± 2.2 | 4.4 ± 1.4 |
| dTTP | 145.5 ± 2.5 | 77.3 ± 8.6 | 41.0 ± 3.8 |
| Formate | 1,517.6 ± 250.1 | 801.4 ± 130.0 | 478.1 ± 40.3 |
| Fumarate | 9.4 ± 1.8 | 4.2 ± 0.7 | 2.2 ± 0.2 |
| GTP | 53.6 ± 2.9 | 21.9 ± 7.9 | 12.4 ± 2.9 |
| Glucose | 740.3 ± 51.6 | 279.8 ± 63.2 | 149.0 ± 18.4 |
| Glutamate | 12,186.7 ± 401.5 | 5,571.3 ± 595.8 | 2,725.4 ± 244.0 |
| Glutamine | 2,820.6 ± 15.9 | 1,411.0 ± 218.3 | 694.5 ± 58.8 |
| Glycerol | 781.2 ± 41.5 | 332.6 ± 34.2 | 210.9 ± 46.5 |
| Glycine | 5,078.8 ± 401.2 | 2,324.3 ± 371.9 | 1,162.5 ± 97.6 |
| Guanosine | 132.7 ± 14 | 55.2 ± 16.1 | 26.6 ± 2.4 |
| Hypoxanthine | 877.9 ± 18.8 | 376.6 ± 69.3 | 167.1 ± 15.4 |
| Inosine | 161.6 ± 35.2 | 33.3 ± 8.4 | 19.6 ± 3.2 |
| Isoleucine | 2,668.0 ± 105.8 | 1,259.1 ± 182.3 | 654.6 ± 63.3 |
| Isovalerate | 255.3 ± 27.2 | 121.5 ± 13.6 | 45.2 ± 3.0 |
| Lactate | 279 ± 38.7 | 123.3 ± 18.6 | 66.7 ± 12.1 |
| Leucine | 4,479.2 ± 107.7 | 2,059.9 ± 343.0 | 1,084.8 ± 104.2 |
| Lysine | 3,142.5 ± 250.2 | 1,630.0 ± 138.5 | 898.5 ± 43.6 |
| Methionine | 1,243.4 ± 56.3 | 614.5 ± 80.6 | 335.8 ± 21.4 |
| Methylhistidine | 127.2 ± 8.9 | 50.0 ± 7.0 | 23.6 ± 1.5 |
| NAD+ | 499.1 ± 33.1 | 234.8 ± 25.1 | 110.4 ± 10.3 |
| NADP+ | 40.6 ± 3.2 | 15.7 ± 5.8 | 9.5 ± 0.4 |
| Niacinamide | 170.5 ± 16.4 | 78.9 ± 7.5 | 33.8 ± 4.2 |
| Nicotinate | 73.6 ± 6.2 | 32 ± 3.8 | 14.8 ± 1.0 |
| Nicotinic acid adenine dinucleotide | 18.3 ± 2.5 | 8.7 ± 1.3 | 4.9 ± 1.6 |
| Phenylalanine | 2,213.1 ± 39.9 | 1,062.5 ± 123.9 | 544.5 ± 33.0 |
| Proline | 2,132.1 ± 145 | 843.2 ± 192.5 | 453.3 ± 57.4 |
| Pyroglutamate | 1,360.7 ± 109.1 | 629.0 ± 88.0 | 285.6 ± 28.2 |
| Succinate | 423.9 ± 36 | 147.4 ± 37.2 | 71.0 ± 9.8 |
| Threonine | 2,494.9 ± 149.1 | 1,120.0 ± 150.2 | 564.9 ± 44.7 |
| Thymidine | 35.4 ± 1.4 | 9.2 ± 4.3 | 3.9 ± 0.2 |
| Trehalose | 313.8 ± 39.4 | 182.3 ± 27.1 | 73.1 ± 5.5 |
| Trimethylamine | 9.9 ± 0.9 | 7.6 ± 0.9 | 6.4 ± 0.9 |
| Tryptophan | 432.3 ± 30.4 | 244.2 ± 44.4 | 140.0 ± 14.8 |
| Tyrosine | 1,266.3 ± 48.1 | 626.3 ± 80.2 | 311.1 ± 26.8 |
| UDP-Acetylglucosamine | 66.7 ± 4.2 | 28.6 ± 4.2 | 18.1 ± 0.6 |
| UDP-galactose | 68.3 ± 2 | 32.1 ± 6.3 | 16.9 ± 1.1 |
| UDP-glucose | 147.8 ± 7 | 78.8 ± 10.8 | 43.1 ± 3.1 |
| UDP-glucuronate | 22.0 ± 0 | 9.4 ± 1.0 | 5.8 ± 1.3 |
| Uracil | 1,013.9 ± 101.6 | 438.5 ± 57.4 | 199.4 ± 14.0 |
| Uridine | 198.4 ± 15 | 61.7 ± 17.0 | 22.6 ± 0.5 |
| Urocanate | 16.0 ± 2.1 | 9.1 ± 1.6 | 3.5 ± 0.4 |
| Valine | 4,583.7 ± 167.1 | 2,167.1 ± 296.2 | 1,106.0 ± 84.9 |
| Xanthosine | 19.5 ± 0.5 | 8.5 ± 1.2 | 4.2 ± 0.2 |

3. Experimental Section
3.1. Bacterial Strains and Cell Culture Growth Conditions
3.2. Metabolic Extraction Procedures
3.3. Colony Formation Units
3.4. Sample Preparation for NMR Analysis
3.5. NMR Data Acquisition and Metabolite Identification
4. Conclusions
Supplementary Files
Acknowledgments
Abbreviations
| D2O | deuterium oxide |
| FID | free induction decay |
| 1H-NMR | proton nuclear magnetic resonance |
| HR-MAS-NMR | high resolution solid-state NMR |
| LB | Luria Bertani Broth medium |
| MAS | magic angle spinning |
| M/C | methanol/chloroform |
| OD | optical density |
| RF | radio frequency |
| TBS | tris-buffered saline |
| TSP | 3-(Trimethylsilyl)propanoic acid |
| X. citri | Xanthomonas axonopodis pv. citri 306 |
Author Contributions
Conflicts of Interest
References
- Ryan, R.P.; Vorholter, F.J.; Potnis, N.; Jones, J.B.; van Sluys, M.A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of xanthomonas: Understanding bacterium-plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Graham, J.H.; Gottwald, T.R.; Cubero, J.; Achor, D.S. Xanthomonas axonopodis pv. Citri: Factors affecting successful eradication of citrus canker. Mol. Plant Pathol. 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Yan, Q.; Wang, N. High-throughput screening and analysis of genes of xanthomonas citri subsp. Citri involved in citrus canker symptom development. Mol. Plant-Microbe Interact. 2012, 25, 69–84. [Google Scholar] [CrossRef]
- Savvides, A.L.; Katsifas, E.A.; Hatzinikolaou, D.G.; Karagouni, A.D. Xanthan production by xanthomonas campestris using whey permeate medium. World J. Microbiol. Biotechnol. 2012, 28, 2759–2764. [Google Scholar] [CrossRef]
- Koek, M.M.; Muilwijk, B.; van der Werf, M.J.; Hankemeier, T. Microbial metabolomics with gas chromatography/mass spectrometry. Anal. Chem. 2006, 78, 1272–1281. [Google Scholar] [CrossRef]
- Liebeke, M.; Dorries, K.; Meyer, H.; Lalk, M. Metabolome analysis of gram-positive bacteria such as staphylococcus aureus by gc-ms and lc-ms. Methods Mol. Biol. 2012, 815, 377–398. [Google Scholar] [CrossRef]
- Martinez, P.; Galvez, S.; Ohtsuka, N.; Budinich, M.; Cortes, M.P.; Serpell, C.; Nakahigashi, K.; Hirayama, A.; Tomita, M.; Soga, T.; et al. Metabolomic study of chilean biomining bacteria acidithiobacillus ferrooxidans strain wenelen and acidithiobacillus thiooxidans strain licanantay. Metabolomics 2013, 9, 247–257. [Google Scholar] [CrossRef]
- Tang, J. Microbial metabolomics. Curr. Genomics 2011, 12, 391–403. [Google Scholar] [CrossRef]
- Tremaroli, V.; Workentine, M.L.; Weljie, A.M.; Vogel, H.J.; Ceri, H.; Viti, C.; Tatti, E.; Zhang, P.; Hynes, A.P.; Turner, R.J.; et al. Metabolomic investigation of the bacterial response to a metal challenge. Appl. Environ. Microbiol. 2009, 75, 719–728. [Google Scholar] [CrossRef]
- Van der Werf, M.J.; Overkamp, K.M.; Muilwijk, B.; Coulier, L.; Hankemeier, T. Microbial metabolomics: Toward a platform with full metabolome coverage. Anal. Biochem. 2007, 370, 17–25. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Wu, C.Y.; Chen, T.; Chen, X.; Zhao, X.M. Integrating metabolomics into a systems biology framework to exploit metabolic complexity: Strategies and applications in microorganisms. Appl. Microbiol. Biotechnol. 2006, 70, 151–161. [Google Scholar] [CrossRef]
- Maharjan, R.P.; Ferenci, T. Global metabolite analysis: The influence of extraction methodology on metabolome profiles of escherichia coli. Anal. Biochem. 2003, 313, 145–154. [Google Scholar] [CrossRef]
- Bolten, C.J.; Kiefer, P.; Letisse, F.; Portais, J.C.; Wittmann, C. Sampling for metabolome analysis of microorganisms. Anal. Chem. 2007, 79, 3843–3849. [Google Scholar]
- Sana, T.R.; Fischer, S.; Wohlgemuth, G.; Katrekar, A.; Jung, K.H.; Ronald, P.C.; Fiehn, O. Metabolomic and transcriptomic analysis of the rice response to the bacterial blight pathogen Xanthomonas oryzae pv. Oryzae. Metabolomics 2010, 6, 451–465. [Google Scholar] [CrossRef]
- Le Belle, J.E.; Harris, N.G.; Williams, S.R.; Bhakoo, K.K. A comparison of cell and tissue extraction techniques using high-resolution 1h-nmr spectroscopy. NMR Biomed. 2002, 15, 37–44. [Google Scholar] [CrossRef]
- Weljie, A.M.; Newton, J.; Mercier, P.; Carlson, E.; Slupsky, C.M. Targeted profiling: quantitative analysis of 1h nmr metabolomics data. Anal. Chem. 2006, 78, 4430–4442. [Google Scholar] [CrossRef]
- Miller, J.H. A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia Coli and Related Bacteria; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1992. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pegos, V.R.; Canevarolo, R.R.; Sampaio, A.P.; Balan, A.; Zeri, A.C.M. Xanthan Gum Removal for 1H-NMR Analysis of the Intracellular Metabolome of the Bacteria Xanthomonas axonopodis pv. citri 306. Metabolites 2014, 4, 218-231. https://doi.org/10.3390/metabo4020218
Pegos VR, Canevarolo RR, Sampaio AP, Balan A, Zeri ACM. Xanthan Gum Removal for 1H-NMR Analysis of the Intracellular Metabolome of the Bacteria Xanthomonas axonopodis pv. citri 306. Metabolites. 2014; 4(2):218-231. https://doi.org/10.3390/metabo4020218
Chicago/Turabian StylePegos, Vanessa R., Rafael R. Canevarolo, Aline P. Sampaio, Andrea Balan, and Ana C. M. Zeri. 2014. "Xanthan Gum Removal for 1H-NMR Analysis of the Intracellular Metabolome of the Bacteria Xanthomonas axonopodis pv. citri 306" Metabolites 4, no. 2: 218-231. https://doi.org/10.3390/metabo4020218
APA StylePegos, V. R., Canevarolo, R. R., Sampaio, A. P., Balan, A., & Zeri, A. C. M. (2014). Xanthan Gum Removal for 1H-NMR Analysis of the Intracellular Metabolome of the Bacteria Xanthomonas axonopodis pv. citri 306. Metabolites, 4(2), 218-231. https://doi.org/10.3390/metabo4020218