Investigation of Daphnia magna Sub-Lethal Exposure to Organophosphate Esters in the Presence of Dissolved Organic Matter Using 1H NMR-Based Metabolomics


Abstract
:1. Introduction
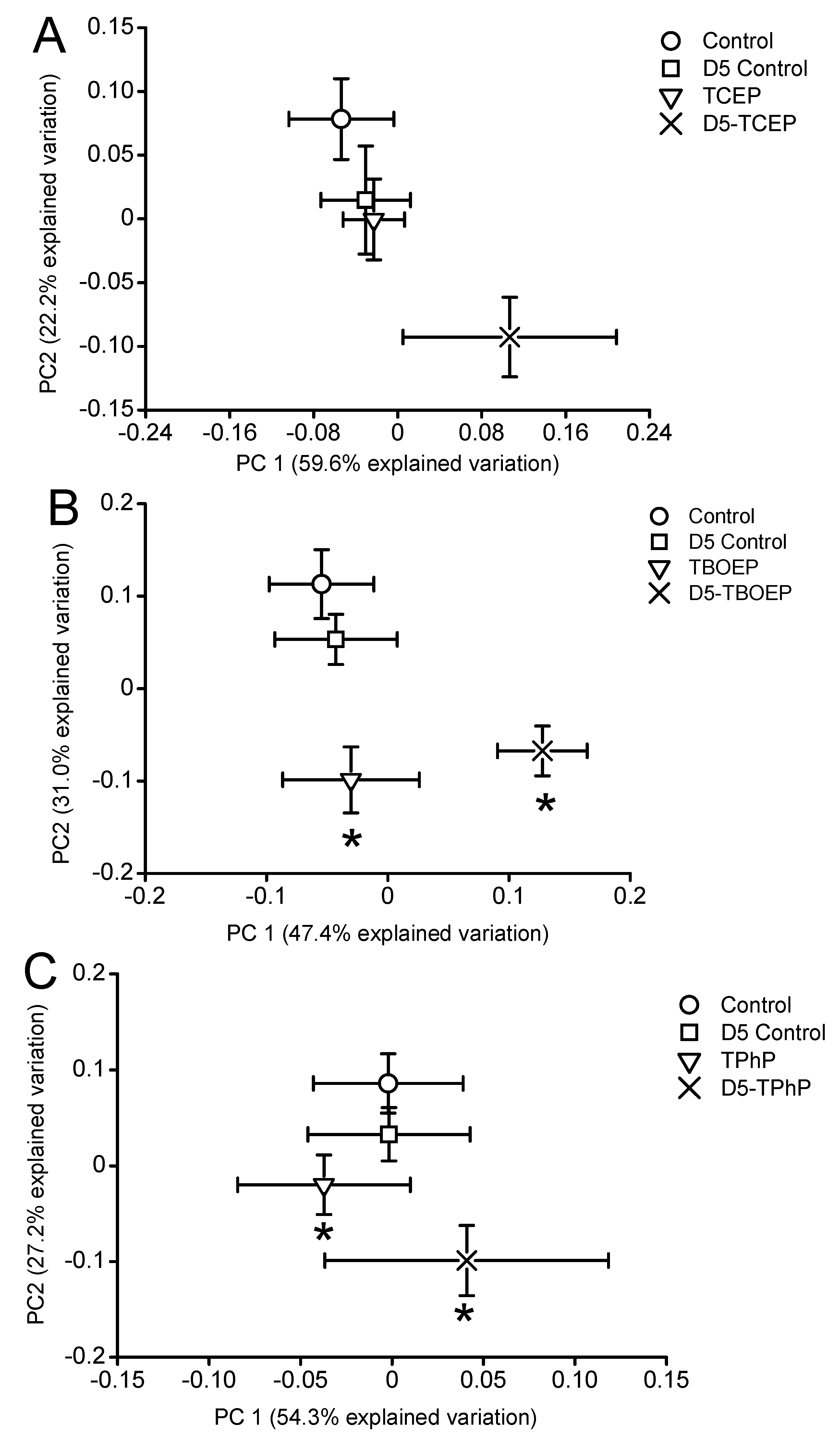
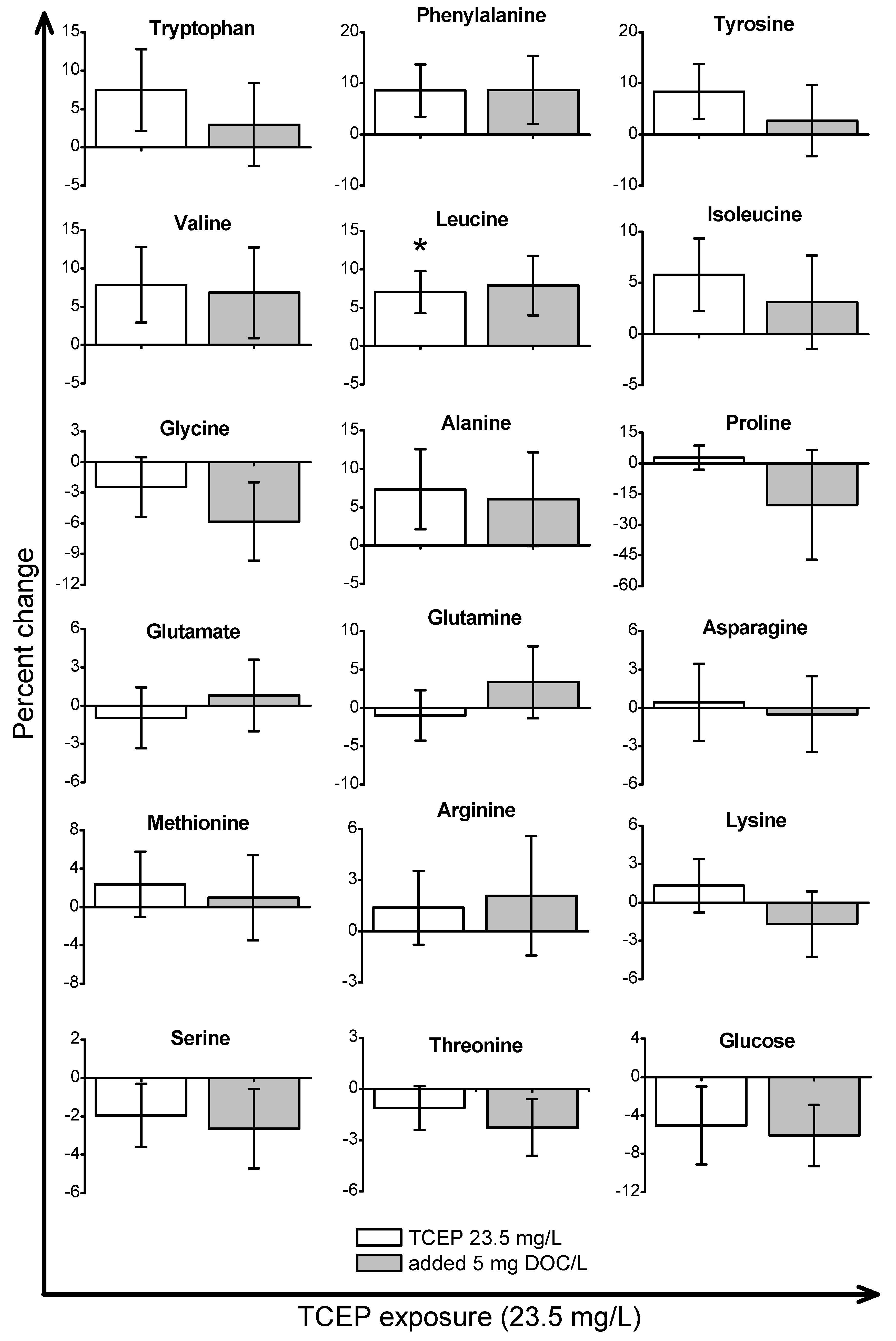
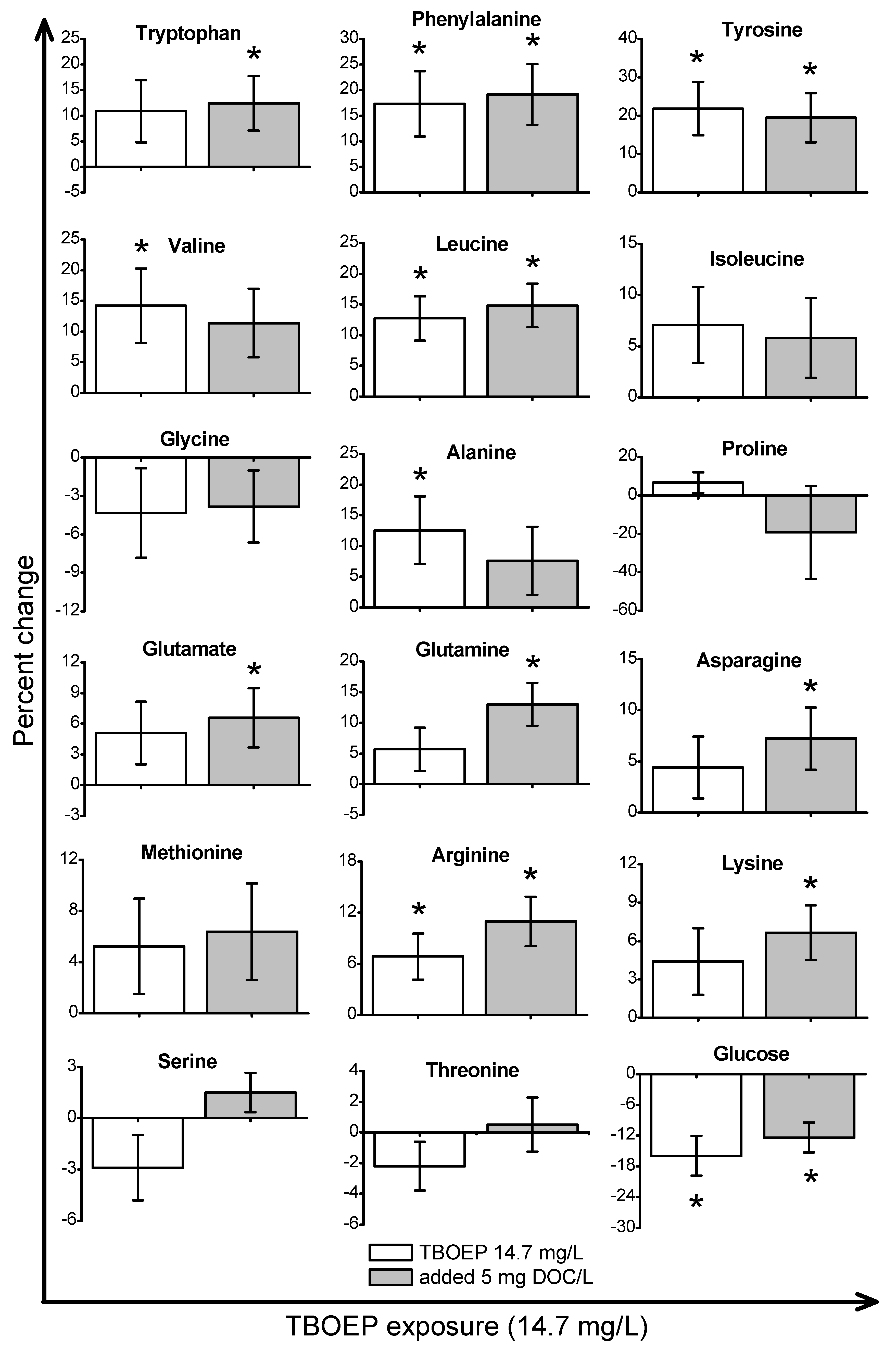
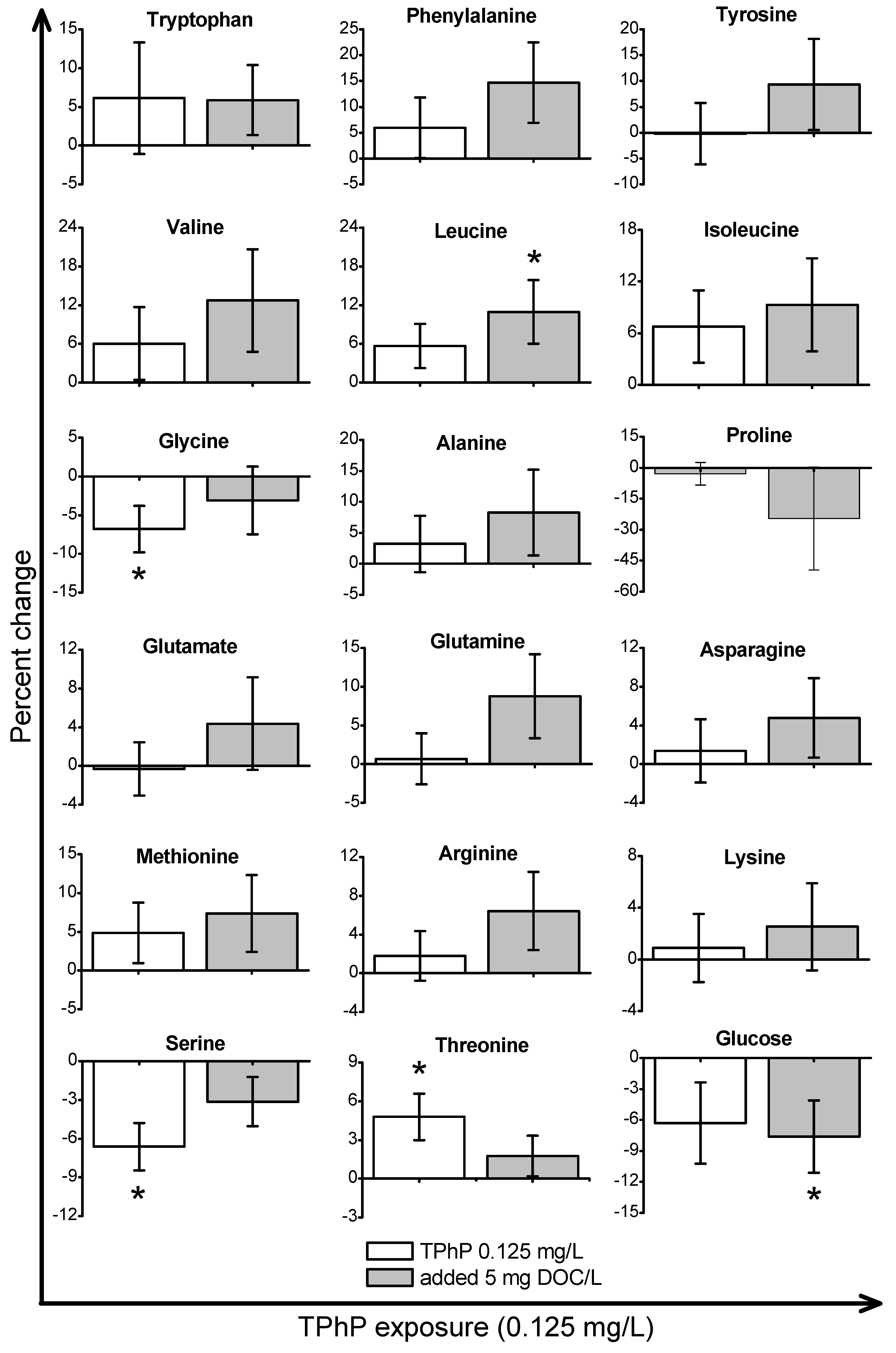
2. Results and Discussion
2.1. Metabolic Response in the Presence of DOM
2.2. Metabolic Response to OPE Exposures in the Absence of DOM
2.3. Metabolic Response to OPE Exposures in the Presence of DOM
3. Materials and Methods
3.1. Daphnia Culturing
3.2. Exposure to OPEs with DOM
3.3. Metabolite Extraction
3.4. 1H NMR Acquisition
3.5. Data and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Kim, S.; Park, J.; Kim, H.S.; Hur, J. Effects of dissolved organic matter (DOM) sources and nature of solid extraction sorbent on recoverable DOM composition: Implication into potential lability of different compound groups. Anal. Bioanal. Chem. 2016, 408, 4809–4819. [Google Scholar] [CrossRef] [PubMed]
- Sobek, S.; Tranvik, L.J.; Prairie, Y.T.; Kortelainen, P.; Cole, J.J. Patterns and regulation of dissolved organic carbon: An analysis of 7500 widely distributed lakes. Limnol. Oceanogr. 2007, 52, 1208–1219. [Google Scholar] [CrossRef]
- Jaffé, R.; McKnight, D.; Maie, N.; Cory, R.; McDowell, W.; Campbell, J. Spatial and temporal variations in DOM composition in ecosystems: The importance of long-term monitoring of optical properties. J. Geophys. Res. Biogeosci. 2008, 113, G04032. [Google Scholar] [CrossRef]
- Saebelfeld, M.; Minguez, L.; Griebel, J.; Gessner, M.O.; Wolinska, J. Humic dissolved organic carbon drives oxidative stress and severe fitness impairments in Daphnia. Aquat. Toxicol. 2017, 182, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.E.; Ouerghemmi, N.; Herrmann, S.; Bouchnak, R.; Timofeyev, M.A.; Menzel, R. Stress by poor food quality and exposure to humic substances: Daphnia magna responds with oxidative stress, lifespan extension, but reduced offspring numbers. Hydrobiologia 2010, 652, 223–236. [Google Scholar] [CrossRef]
- Chen, S.; Ke, R.; Zha, J.; Wang, Z.; Khan, S.U. Influence of humic acid on bioavailability and toxicity of benzo [k] fluoranthene to Japanese medaka. Environ. Sci. Technol. 2008, 42, 9431–9436. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, C.; Cheng, P.; He, X.; Zhu, Y.; Zhang, Y. Influences of humic acid on the bioavailability of phenanthrene and alkyl phenanthrenes to early life stages of marine medaka (Oryzias melastigma). Environ. Pollut. 2016, 210, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Liu, J.; Yin, Y.; Shen, M. Evaluating the sorption of organophosphate esters to different sourced humic acids and its effects on the toxicity to Daphnia magna. Environ. Toxicol. Chem. 2013, 32, 2755–2761. [Google Scholar] [CrossRef] [PubMed]
- Poerschmann, J.; Kopinke, F. Sorption of very hydrophobic organic compounds (VHOCs) on dissolved humic organic matter (DOM). 2. Measurement of sorption and application of a Flory−Huggins concept to interpret the data. Environ. Sci. Technol. 2001, 35, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Haftka, J.J.; Scherpenisse, P.; Jonker, M.T.; Hermens, J.L. Using polyacrylate-coated SPME fibers to quantify sorption of polar and ionic organic contaminants to dissolved organic carbon. Environ. Sci. Technol. 2013, 47, 4455–4462. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Xia, X.; Bi, S.; Jiang, X.; Wang, H.; Zhai, Y.; Wen, W. Quantifying Bioavailability of Pyrene Associated with Dissolved Organic Matter of Various Molecular Weights to Daphnia magna. Environ. Sci. Technol. 2017, 52, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Dai, Z.; Rabearisoa, A.H.; Zhao, P.; Jiang, X. Comparing humic substance and protein compound effects on the bioaccumulation of perfluoroalkyl substances by Daphnia magna in water. Chemosphere 2015, 119, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Venier, M.; Dove, A.; Romanak, K.; Backus, S.; Hites, R. Flame retardants and legacy chemicals in Great Lakes’ water. Environ. Sci. Technol. 2014, 48, 9563–9572. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, U.E.; Möller, A.; Xie, Z.; Ebinghaus, R.; Einax, J.W. Occurrence and fate of organophosphorus flame retardants and plasticizers in coastal and marine surface waters. Water Res. 2012, 46, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Marklund, A.; Andersson, B.; Haglund, P. Organophosphorus flame retardants and plasticizers in Swedish sewage treatment plants. Environ. Sci. Technol. 2005, 39, 7423–7429. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, M.; Douville, M.; Houde, M. Chronic toxicity evaluation of the flame retardant tris (2-butoxyethyl) phosphate (TBOEP) using Daphnia magna transcriptomic response. Chemosphere 2015, 132, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Su, G.; Zou, M.; Yu, L.; Letcher, R.J.; Yu, H.; Giesy, J.P.; Zhou, B.; Liu, C. Effects of tris (1, 3-dichloro-2-propyl) phosphate on growth, reproduction, and gene transcription of Daphnia magna at environmentally relevant concentrations. Environ. Sci. Technol. 2015, 49, 12975–12983. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, H.; Dang, Y.; Liu, C. Effects of triphenyl phosphate on growth, reproduction and transcription of genes of Daphnia magna. Aquat. Toxicol. 2017, 195, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yuan, S.; Su, G.; Li, M.; Wang, Q.; Zhu, G.; Letcher, R.J.; Li, Y.; Han, Z.; Liu, C. Whole-Life-Stage Characterization in the Basic Biology of Daphnia magna and Effects of TDCIPP on Growth, Reproduction, Survival, and Transcription of Genes. Environ. Sci. Technol. 2017, 51, 13967–13975. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, L.D.; Loguinov, A.V.; Teng, Q.; Antczak, P.; Dailey, K.P.; Nowinski, D.T.; Kornbluh, J.; Lin, X.X.; Lachenauer, E.; Arai, A. Gene transcription, metabolite and lipid profiling in eco-indicator Daphnia magna indicate diverse mechanisms of toxicity by legacy and emerging flame-retardants. Environ. Sci. Technol. 2015, 49, 7400–7410. [Google Scholar] [CrossRef] [PubMed]
- Brandao, F.; Cappello, T.; Raimundo, J.; Santos, M.A.; Maisano, M.; Mauceri, A.; Pacheco, M.; Pereira, P. Unravelling the mechanisms of mercury hepatotoxicity in wild fish (Liza aurata) through a triad approach: Bioaccumulation, metabolomic profiles and oxidative stress. Metallomics 2015, 7, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.; Staff, F.J.; Widdows, J.; Compton, R.M.; Falciani, F.; Viant, M.R. Discovery of metabolic signatures for predicting whole organism toxicology. Toxicol. Sci. 2010, 115, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Giannetto, A.; Parrino, V.; Maisano, M.; Oliva, S.; De Marco, G.; Guerriero, G.; Mauceri, A.; Fasulo, S. Baseline levels of metabolites in different tissues of mussel Mytilus galloprovincialis (Bivalvia: Mytilidae). Comp. Biochem. Physiol. D 2018, 26, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Keun, H.C.; Ebbels, T.M.; Antti, H.; Bollard, M.E.; Beckonert, O.; Schlotterbeck, G.; Senn, H.; Niederhauser, U.; Holmes, E.; Lindon, J.C. Analytical reproducibility in 1H NMR-based metabonomic urinalysis. Chem. Res. Toxicol. 2002, 15, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Maisano, M.; Mauceri, A.; Fasulo, S. 1H NMR-based metabolomics investigation on the effects of petrochemical contamination in posterior adductor muscles of caged mussel Mytilus galloprovincialis. Ecotoxicol. Environ. Saf. 2017, 142, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Fasulo, S.; Iacono, F.; Cappello, T.; Corsaro, C.; Maisano, M.; D’Agata, A.; Giannetto, A.; De Domenico, E.; Parrino, V.; Paro, G.L. Metabolomic investigation of Mytilus galloprovincialis (Lamarck 1819) caged in aquatic environments. Ecotoxicol. Environ. Saf. 2012, 84, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, I.; Demiri, B.; Xu, S.; Constantin, A.; Yan, N.D.; Cristescu, M.E. An integrated multi-disciplinary approach for studying multiple stressors in freshwater ecosystems: Daphnia as a model organism. Integr. Comp. Biol. 2011, 51, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, Y.; Fukunaga, I.; Itoh, H. Distribution of organophosphoric acid triesters between water and sediment at a sea-based solid waste disposal site. J. Mater. Cycle Waste Manag. 1999, 1, 53–61. [Google Scholar]
- European Union. European Union Risk Assessment Report: Tris(2-Chloroethyl) Phosphate (TCEP); EU: Brussels, Belgium, 2009; pp. 1–213. [Google Scholar]
- Douville, M.; Jean, K.; Houde, M. Multitrophic aquatic toxicity of emerging brominated and phosphorous flame retardants. Fresen. Environ. Bull. 2016, 25, 3265–3271. [Google Scholar]
- Meems, N.; Steinberg, C.; Wiegand, C. Direct and interacting toxicological effects on the waterflea (Daphnia magna) by natural organic matter, synthetic humic substances and cypermethrin. Sci. Total Environ. 2004, 319, 123–136. [Google Scholar] [CrossRef]
- Taylor, N.S.; Kirwan, J.A.; Yan, N.D.; Viant, M.R.; Gunn, J.M.; McGeer, J.C. Metabolomics confirms that dissolved organic carbon mitigates copper toxicity. Environ. Toxicol. Chem. 2016, 35, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Roznere, I.; Watters, G.T.; Wolfe, B.A.; Daly, M. Nontargeted metabolomics reveals biochemical pathways altered in response to captivity and food limitation in the freshwater mussel Amblema plicata. Comp. Biochem. Physiol. D 2014, 12, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Shinji, J.; Okutsu, T.; Jayasankar, V.; Jasmani, S.; Wilder, M.N. Metabolism of amino acids during hyposmotic adaptation in the whiteleg shrimp, Litopenaeus vannamei. Amino Acids 2012, 43, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Yoshikawa, N.; Sarower, M.G.; Okada, S. Physiological function and metabolism of free D-alanine in aquatic animals. Biol. Pharm. Bull. 2005, 28, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Cristale, J.; Vázquez, A.G.; Barata, C.; Lacorte, S. Priority and emerging flame retardants in rivers: Occurrence in water and sediment, Daphnia magna toxicity and risk assessment. Environ. Int. 2013, 59, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Guo, J.; Wang, Y.; Li, Z.; Liang, K.; Corcoran, M.B.; Hosseini, S.; Bonina, S.M.; Rockne, K.J.; Sturchio, N.C. Organophosphate Esters in Sediment of the Great Lakes. Environ. Sci. Technol. 2017, 51, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Ripszam, M.; Paczkowska, J.; Figueira, J.; Veenaas, C.; Haglund, P. Dissolved organic carbon quality and sorption of organic pollutants in the Baltic Sea in light of future climate change. Environ. Sci. Technol. 2015, 49, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Chiou, C.T.; Malcolm, R.L.; Brinton, T.I.; Kile, D.E. Water solubility enhancement of some organic pollutants and pesticides by dissolved humic and fulvic acids. Environ. Sci. Technol. 1986, 20, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Environment Canada. Biological Test Method: Reference Method for Determining Acute Lethality of Effluents to Daphnia magna; Environmental Technology Centre: Ottawa, ON, Canada, 2000.
- Ritchie, J.D.; Perdue, E.M. Proton-binding study of standard and reference fulvic acids, humic acids, and natural organic matter. Geochim. Cosmochim. Acta 2003, 67, 85–96. [Google Scholar] [CrossRef]
- Agatz, A.; Cole, T.A.; Preuss, T.G.; Zimmer, E.; Brown, C.D. Feeding inhibition explains effects of imidacloprid on the growth, maturation, reproduction, and survival of Daphnia magna. Environ. Sci. Technol. 2013, 47, 2909–2917. [Google Scholar] [CrossRef] [PubMed]
- Nagato, E.G.; Lankadurai, B.P.; Soong, R.; Simpson, A.J.; Simpson, M.J. Development of an NMR microprobe procedure for high-throughput environmental metabolomics of Daphnia magna. Magn. Reson. Chem. 2015, 53, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.J.; Brown, S.A. Purge NMR: Effective and easy solvent suppression. J. Magn. Reson. 2005, 175, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. 1H NMR-based metabolomics of Daphnia magna responses after sub-lethal exposure to triclosan, carbamazepine and ibuprofen. Comp. Biochem. Physiol. D 2016, 19, 199–210. [Google Scholar]
- Nagato, E.G.; Simpson, A.J.; Simpson, M.J. Metabolomics reveals energetic impairments in Daphnia magna exposed to diazinon, malathion and bisphenol-A. Aquat. Toxicol. 2016, 170, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, M.N.; Nagato, E.G.; Lankadurai, B.P.; Simpson, A.J.; Simpson, M.J. Analysis of Sub-Lethal Toxicity of Perfluorooctane Sulfonate (PFOS) to Daphnia magna Using 1H Nuclear Magnetic Resonance-Based Metabolomics. Metabolites 2017, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Lewis, I.A.; Hegeman, A.D.; Anderson, M.E.; Li, J.; Schulte, C.F.; Westler, W.M.; Eghbalnia, H.R.; Sussman, M.R.; Markley, J.L. Metabolite identification via the madison metabolomics consortium database. Nat. Biotechnol. 2008, 26, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; Simpson, A.J.; Simpson, M.J. Metabolomic responses to sublethal contaminant exposure in neonate and adult Daphnia magna. Environ. Toxicol. Chem. 2017, 36, 938–946. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound and Chemical Structure | Aqueous Solubility (mg/L) | Log KOW | 48-h Daphnia magna LC50 (mg/L) | Estimated Log KDOC |
|---|---|---|---|---|
 Tris(2-chloroethyl) phosphate (TCEP) | 7000 a | 1.43 a | 24-h EC50 = 235 mg/L b | 1.81 c |
 Tris(2-butoxyethyl) phosphate (TBOEP) | 1100 a | 3.65 a | 147 mg/L d | 3.18 c |
 Triphenyl phosphate (TPhP) | 1.90 a | 4.76 a | 1.25 mg/L e | 3.87 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovacevic, V.; Simpson, A.J.; Simpson, M.J. Investigation of Daphnia magna Sub-Lethal Exposure to Organophosphate Esters in the Presence of Dissolved Organic Matter Using 1H NMR-Based Metabolomics. Metabolites 2018, 8, 34. https://doi.org/10.3390/metabo8020034
Kovacevic V, Simpson AJ, Simpson MJ. Investigation of Daphnia magna Sub-Lethal Exposure to Organophosphate Esters in the Presence of Dissolved Organic Matter Using 1H NMR-Based Metabolomics. Metabolites. 2018; 8(2):34. https://doi.org/10.3390/metabo8020034
Chicago/Turabian StyleKovacevic, Vera, André J. Simpson, and Myrna J. Simpson. 2018. "Investigation of Daphnia magna Sub-Lethal Exposure to Organophosphate Esters in the Presence of Dissolved Organic Matter Using 1H NMR-Based Metabolomics" Metabolites 8, no. 2: 34. https://doi.org/10.3390/metabo8020034
APA StyleKovacevic, V., Simpson, A. J., & Simpson, M. J. (2018). Investigation of Daphnia magna Sub-Lethal Exposure to Organophosphate Esters in the Presence of Dissolved Organic Matter Using 1H NMR-Based Metabolomics. Metabolites, 8(2), 34. https://doi.org/10.3390/metabo8020034