Dysregulation of the Tryptophan Pathway Evidences Gender Differences in COPD



Abstract
:1. Introduction
2. Results
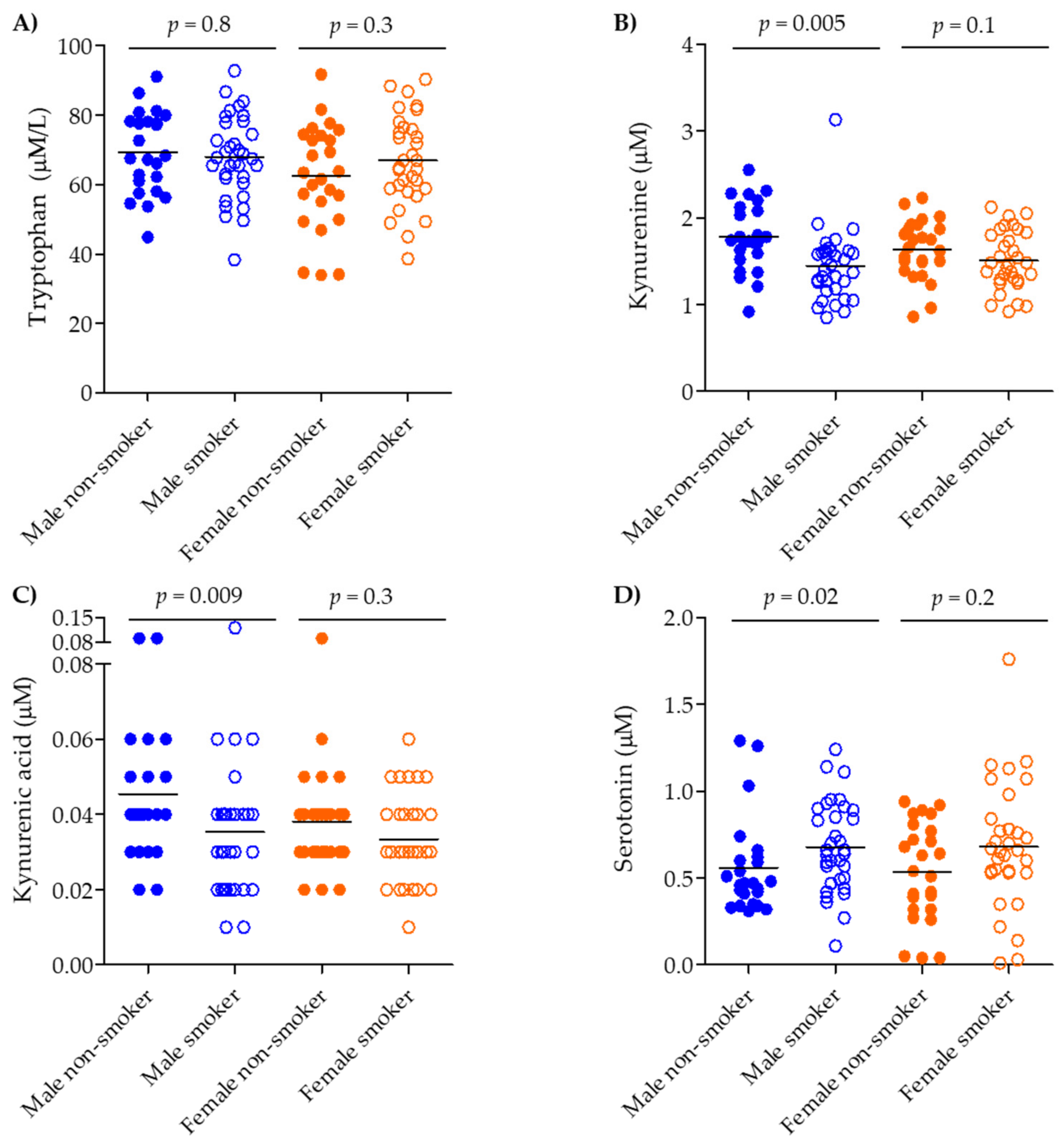
2.1. Serum Levels of the Tryptophan Pathway Metabolites
2.2. Serum IDO and TPH Activity
2.3. Correlation of Serum Tryptophan Metabolites and IDO with Clinical Data
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Design
4.2. Tryptophan Pathway Metabolites Measurement by LC-MS/MS
4.3. Cytokine and Chemokine Measurements
4.4. Measurement of IDO and TPH Activity
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Agusti, A.; Barnes, P.J. Update in chronic obstructive pulmonary disease 2011. Am. J. Respir. Crit. Care Med. 2012, 185, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Paré, P.D.; Hackett, T.-L. The Contribution of Small Airway Obstruction to the Pathogenesis of Chronic Obstructive Pulmonary Disease. Physiol. Rev. 2017, 97, 529–552. [Google Scholar] [CrossRef] [PubMed]
- Rabe, K.F.; Watz, H. Chronic Obstructive Pulmonary Disease. Lancet 2017, 389, 1931–1940. [Google Scholar] [CrossRef]
- Forsslund, H.; Mikko, M.; Karimi, R.; Grunewald, J.; Wheelock, Å.M.; Wahlström, J.; Sköld, C.M. Distribution of T-cell subsets in BAL fluid of patients with mild to moderate COPD depends on current smoking status and not airway obstruction. Chest 2014, 145, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Rojas, M.I.; Ramírez-Venegas, A.; Limón-Camacho, L.; Ochoa, L.; Hernández-Zenteno, R.; Sansores, R.H. Increase of Th17 cells in peripheral blood of patients with chronic obstructive pulmonary disease. Respir. Med. 2011, 105, 1648–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J. Inflammatory mechanisms in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2016, 138, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Wei, X.; Wang, J.; Cheng, Z.; Sun, W. Caveolin-1 Promotes the Imbalance of Th17/Treg in Patients with Chronic Obstructive Pulmonary Disease. Inflammation 2016, 39, 2008–2015. [Google Scholar] [CrossRef] [PubMed]
- Shaykhiev, R.; Sackrowitz, R.; Fukui, T.; Zuo, W.L.; Chao, I.W.; Strulovici-Barel, Y.; Downey, R.J.; Crystal, R.G. Smoking-induced CXCL14 expression in the human airway epithelium links chronic obstructive pulmonary disease to lung cancer. Am. J. Respir. Cell Mol. Biol. 2013, 49, 41–425. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Otten, W.; Merlot, E. Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids 2011, 41, 1195–1205. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Lau, W.K.W.; Chan-Yeung, M.M.W.; Yip, B.H.K.; Cheung, A.H.K.; Ip, M.S.M.; Mak, J.C.W. The role of circulating serotonin in the development of chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e31617. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.W.S.; Terentis, A.C.; King, N.J.C.; Thomas, S.R. Role of indoleamine 2,3-dioxygenase in health and disease. Clin. Sci. 2015, 129, 601–672. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.A.; Ottiger, M.; Vögeli, A.; Steuer, C.; Bernasconi, L.; Thomann, R.; Christ-Crain, M.; Henzen, C.; Hoess, C.; Zimmerli, W.; et al. Activation of the Serotonin Pathway is Associated with Poor Outcome in COPD Exacerbation: Results of a Long-Term Cohort Study. Lung 2017, 195, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed]
- Gál, E.M.; Sherman, A.D. l-Kynurenine Its synthesis and possible regulatory function in brain. Neurochem. Res. 1980, 5, 223–239. [Google Scholar] [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Herraiz, T.; Chaparro, C. Human monoamine oxidase is inhibited by tobacco smoke: β-carboline alkaloids act as potent and reversible inhibitors. Biochem. Biophys. Res. Commun. 2005, 326, 378–386. [Google Scholar] [CrossRef]
- Hayaishi, O. Utilization of Superoxide Anion by Indoleamine Oxygenase-Catalyzed Tryptophan and Indoleamine Oxidation. Adv. Exp. Med. Biol. 1996, 398, 285–289. [Google Scholar]
- Sage, L.K.; Fox, J.M.; Mellor, A.L.; Tompkins, S.M.; Tripp, R.A. Indoleamine 2,3-Dioxygenase (IDO) Activity During the Primary Immune Response to Influenza Infection Modifies the Memory T Cell Response to Influenza Challenge. Viral Immunol. 2014, 27, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Maneechotesuwan, K.; Wongkajornsilp, A.; Adcock, I.M.; Barnes, P.J. Simvastatin suppresses airway IL-17 and upregulates IL-10 in patients with stable COPD. Chest 2015, 148, 1164–1176. [Google Scholar] [CrossRef]
- Morecroft, I. Functional Interactions between 5-Hydroxytryptamine Receptors and the Serotonin Transporter in Pulmonary Arteries. J. Pharmacol. Exp. Ther. 2005, 313, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, A.A.; Davies, B.; Castagnoli, N. Isolation and characterization of a monoamine oxidase B selective inhibitor from tobacco smoke. Bioorg. Med. Chem. 2006, 14, 3392–3398. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Kolmert, J.; Yang, M.; Reinke, S.N.; Kamleh, M.A.; Snowden, S.; Heyder, T.; Levänen, B.; Erle, D.J.; Sköld, C.M.; et al. Metabolomics analysis identifies sex-associated metabotypes of oxidative stress and the autotaxin-lysoPA axis in COPD. Eur. Respir. J. 2017, 49. [Google Scholar] [CrossRef] [PubMed]
- Forsslund, H.; Yang, M.; Mikko, M.; Karimi, R.; Nyrén, S.; Engvall, B.; Grunewald, J.; Merikallio, H.; Kaarteenaho, R.; Wahlström, J.; et al. Gender differences in the t-cell profiles of the airways in COPD patients associated with clinical phenotypes. Int. J. COPD 2017, 12, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Balgoma, D.; Yang, M.; Sjödin, M.; Snowden, S.; Karimi, R.; Levänen, B.; Merikallio, H.; Kaarteenaho, R.; Palmberg, L.; Larsson, K.; et al. Linoleic acid-derived lipid mediators increase in a female-dominated subphenotype of COPD. Eur. Respir. J. 2016, 47, 1645–1656. [Google Scholar] [CrossRef] [Green Version]
- Ubhi, B.K.; Riley, J.H.; Shaw, P.A.; Lomas, D.A.; Tal-Singers, R.; MacNeef, W.; Griffin, J.L.; Connor, S.C. Metabolic profiling detects biomarkers of protein degradation in COPD patients. Eur. Respir. J. 2012, 40, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Maneechotesuwan, K.; Kasetsinsombat, K.; Wongkajornsilp, A.; Barnes, P.J. Decreased indoleamine 2,3-dioxygenase activity and IL-10/IL-17A ratio in patients with COPD. Thorax 2013, 68, 330–337. [Google Scholar] [CrossRef]
- Lewis, G.D.; Ngo, D.; Hemnes, A.R.; Farrell, L.; Domos, C.; Pappagianopoulos, P.P.; Dhakal, B.P.; Souza, A.; Shi, X.; Pugh, M.E.; et al. Metabolic Profiling of Right Ventricular-Pulmonary Vascular Function Reveals Circulating Biomarkers of Pulmonary Hypertension. J. Am. Coll. Cardiol. 2016, 67, 174–189. [Google Scholar] [CrossRef] [Green Version]
- Hervé, C.; Beyne, P.; Jamault, H.; Delacoux, E. Determination of tryptophan and its kynurenine pathway metabolites in human serum by high-performance liquid chromatography with simultaneous ultraviolet and fluorimetric detection. J. Chromatogr. B Biomed. Appl. 1996, 675, 157–161. [Google Scholar] [CrossRef]
- Meier, M.A.; Ottiger, M.; Vögeli, A.; Steuer, C.; Bernasconi, L.; Thomann, R.; Christ-Crain, M.; Henzen, C.; Hoess, C.; Zimmerli, W.; et al. Activation of the tryptophan/serotonin pathway is associated with severity and predicts outcomes in pneumonia: Results of a long-term cohort study. Clin. Chem. Lab. Med. 2017, 55, 1060–1069. [Google Scholar] [CrossRef]
- Suzuki, Y.; Suda, T.; Yokomura, K.; Suzuki, M.; Fujie, M.; Furuhashi, K.; Hahimoto, D.; Enomto, N.; Fujisawa, T.; Nakamura, Y.; et al. Serum activity of indoleamine 2,3-dioxygenase predicts prognosis of community-acquired pneumonia. J. Infect. 2011, 63, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Wendt, C.; Gulcev, M.; Reilly, C.; Griffin, T.; Broeckling, C.; Sandri, B.; Witthuhn, B.; Hodgson, S.; Woodruff, P. Tryptophan catabolism in acute exacerbations of chronic obstructive pulmonary disease. Int. J. Chron. Obstruct. Pulm. Dis. 2016, 11, 2435–2446. [Google Scholar] [CrossRef] [PubMed]
- Pertovaara, M.; Heliövaara, M.; Raitala, A.; Oja, S.S.; Knekt, P.; Hurme, M. The activity of the immunoregulatory enzyme indoleamine 2,3-dioxygenase is decreased in smokers. Clin. Exp. Immunol. 2006, 145, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C. Pathophysiology of airflow limitation in chronic obstructive pulmonary disease. Lancet 2004, 364, 709–721. [Google Scholar] [CrossRef]
- Saetta, M.; Mariani, M.; Panina-Bordignon, P.; Turato, G.; Buonsanti, C.; Baraldo, S.; Bellettato, C.M.; Papi, A.; Corbetta, L.; Zuin, R.; et al. Increased expression of the chemokine receptor CXCR3 and its ligand CXCL10 in peripheral airways of smokers with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2002, 165, 1404–1409. [Google Scholar] [CrossRef]
- Costa, C.; Rufino, R.; Traves, S.L.; Lapa, E.; Silva, J.R.; Barnes, P.J.; Donnelly, L.E. CXCR3 and CCR5 chemokines in induced sputum from patients with COPD. Chest 2008, 133, 26–33. [Google Scholar] [CrossRef]
- Taylor, M.W.; Feng, G.S. Relationship between interferon-gamma, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar] [CrossRef]
- Han, M.L.K.; Postma, D.; Mannino, D.M.; Giardino, N.D.; Buist, S.; Curtis, J.L.; Martinez, F.J. Gender and chronic obstructive pulmonary disease: Why it matters. Am. J. Respir. Crit. Care Med. 2007, 176, 1179–1184. [Google Scholar] [CrossRef]
- Baggiolini, M.; Dewald, B.; Moser, B. lnterleukin-8 and Related Chemotactic Cytokines—CXC and CC Chemokines. Adv. Immunol. 1993, 55, 97–179. [Google Scholar]
- Rollins, B.J.; Walz, A.; Baggiolini, M. Recombinant human MCP-1/JE induces chemotaxis, calcium flux, and the respiratory burst in human monocytes. Blood 1991, 78, 1112–1116. [Google Scholar] [Green Version]
- Tam, A.; Churg, A.; Wright, J.L.; Zhou, S.; Kirby, M.; Coxson, H.O.; Lam, S.; Man, S.F.P.; Sin, D.D. Sex differences in airway remodeling in a mouse model of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2016, 193, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.P.; Snyder, E.M.; Frantz, R.P.; Turner, S.T.; Johnson, B.D. Repeat length polymorphism of the serotonin transporter gene influences pulmonary artery pressure in heart failure. Am. Heart J. 2007, 153, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Eddahibi, S.; Chaouat, A.; Morrell, N.; Fadel, E.; Fuhrman, C.; Bugnet, A.S.; Dartevelle, P.; Housset, B.; Hamon, M.; Weitzenblum, E.; et al. Polymorphism of the Serotonin Transporter Gene and Pulmonary Hypertension in Chronic Obstructive Pulmonary Disease. Circulation 2003, 108, 1839–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Jiang, X.; Yuan, L.; Li, P.; Wang, J.; Wang, P.; Zhang, L.; Sun, B. Effect of Simvastatin on 5-HT and 5-HTT in a Rat Model of Pulmonary Artery Hypertension. Cell. Physiol. Biochem. 2015, 37, 1712–1724. [Google Scholar]
- Wang, Q.; Chen, D.; Nicholson, P.; Cheng, S.; Alen, M.; Mao, L.; Cheng, S. The Associations of Serum Serotonin with Bone Traits Are Age- and Gender-Specific. PLoS ONE 2014, 9, e109028. [Google Scholar] [CrossRef] [PubMed]
- Saetta, M.; Baraldo, S.; Corbino, L.; Turato, G.; Braccioni, F.; Rea, F.; Cavallesco, G.; Tropeano, G.; Mapp, C.E.; Maestrelli, P.; et al. CD8+ve cells in the lungs of smokers with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, 711–717. [Google Scholar] [CrossRef]
- Xia, H.J.; Zhang, G.H.; Wang, R.R.; Zheng, Y.T. The influence of age and sex on the cell counts of peripheral blood leukocyte subpopulations in Chinese rhesus macaques. Cell. Mol. Immunol. 2009, 6, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, F.; Verga, M.; Reggente, M.; Maria Casanova, F.; Santus, P.; Blasi, F.; Allegra, L.; Centanni, S. Anxiety and depression in COPD patients: The roles of gender and disease severity. Respir. Med. 2006, 100, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic Acid: The Janus-faced role of an immunomodulatory tryptophan metabolite and its link to pathological conditions. Front. Immunol. 2018, 8, 1957. [Google Scholar] [CrossRef]
- Van der Leek, A.P.; Yanishevsky, Y.; Kozyrskyj, A.L. The kynurenine pathway as a novel link between allergy and the gut microbiome. Front. Immunol. 2017, 8, 1374. [Google Scholar] [CrossRef]
- Tam, A.; Bates, J.H.T.; Churg, A.; Wright, J.L.; Man, S.F.P.; Sin, D.D. Sex-related differences in pulmonary function following 6 months of cigarette exposure: Implications for sexual dimorphism in mild COPD. PLoS ONE 2016, 11, e0164835. [Google Scholar] [CrossRef] [PubMed]
- Han, M.L.K.; Arteaga-Solis, E.; Blenis, J.; Bourjeily, G.; Clegg, D.J.; DeMeo, D.; Duffy, J.; Gaston, B.; Heller, N.M.; Hemnes, A.; et al. Female sex and gender in lung/sleep health and disease: Increased understanding of basic biological, pathophysiological, and behavioral mechanisms leading to better health for female patients with lung disease. Am. J. Respir. Crit. Care Med. 2018, 198, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous measurement of serum tryptophan and kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite (µM) | Never-Smokers | Smokers | COPD Smokers | COPD Ex-Smokers | p-value |
|---|---|---|---|---|---|
| Tryptophan | 67.2 (65.8, 77.6) | 71.5 (61.6, 79.4) | 63.1 (56.5, 68.9) | 67.6 (54.6, 77.7) | 0.2 |
| Kynurenine | 1.7 (1.5, 1.9) | 1.5 (1.1, 1.6) | 1.4 (1.3, 1.8) | 1.9 (1.5, 2.0) | 0.003 |
| Kynurenic acid | 0.04 (0.03, 0.04) | 0.03 (0.02, 0.04) | 0.04 (0.03, 0.04) | 0.05 (0.04, 0.06) | 0.009 |
| Serotonin | 0.5 (0.4, 0.75) | 0.6 (0.5, 0.8) | 0.7 (0.5, 0.9) | 0.4 (0.3, 0.7) | 0.03 |
| Cytokines and Chemokines | COPD Joint Gender | COPD Females | COPD Males | ||||||
|---|---|---|---|---|---|---|---|---|---|
| XY Pairs | r | p-value | XY Pairs | r | p-value | XY Pairs | r | p-value | |
| Blood cells | |||||||||
| CCL3 | 21 | 0.41 | 0.06 | 9 | 0.88 | 0.0016 | 12 | 0.01 | 1.0 |
| CCL4 | 18 | 0.56 | 0.02 | 8 | 0.74 | 0.04 | 10 | 0.39 | 0.3 |
| CXCL9 | 18 | 0.60 | 0.009 | 8 | 0.93 | 0.0009 | 10 | 0.26 | 0.5 |
| CXCL10 | 18 | 0.43 | 0.07 | 8 | 0.88 | 0.004 | 10 | 0.08 | 0.8 |
| CXCL12 | 17 | 0.43 | 0.09 | 9 | 0.73 | 0.02 | 8 | −0.14 | 0.7 |
| IL-1b | 20 | 0.46 | 0.04 | 9 | 0.62 | 0.08 | 11 | 0.45 | 0.2 |
| IL-12 | 19 | 0.40 | 0.09 | 9 | 0.67 | 0.05 | 10 | −0.08 | 0.8 |
| IL-13 | 19 | 0.46 | 0.04 | 8 | 0.81 | 0.01 | 11 | -* | 1.0 |
| BAL cells | |||||||||
| CXCR4 on CD8 | 20 | −0.37 | 0.1 | 8 | −0.76 | 0.03 | 12 | −0.03 | 0.9 |
| FOXP3+ of CD4+ | 19 | 0.57 | 0.01 | 8 | 0.65 | 0.08 | 11 | 0.39 | 0.2 |
| Parameters | Never-Smokers (n = 39) | Smokers (n = 40) | COPD Smokers (n = 27) | COPD Ex-Smokers (n = 10) |
|---|---|---|---|---|
| Gender distribution (number of males) | 19 | 20 | 14 | 5 |
| Age (years) | 60.0 (51.5, 64.0) | 52.5 (49.0, 57.8)* | 59.5 (55.8, 63.0)** | 63.5 (53.8, 65.0)** |
| BMI | 25.6 (23.3, 28.9) | 24.5 (22.1, 26.1) | 24.1 (21.1, 27.0) | 27.6 (22.7, 30.2) |
| Smoking (packyears) | N.A. | 33.5 (28.3, 40.0)* | 42.0 (35.8, 45.5)*,** | 32.5 (21.5, 38.5)* |
| Current smoking/day | N.A. | 17.0 (12.0, 20.0) | 20.0 (14.0, 20.0) | N.A. |
| GOLD Stage (1/2) | N.A. | N.A. | 12/14 | 5/5 |
| GOLD-2011 (A/B/C) | N.A. | N.A. | 19/7 | 6/3/1 |
| Blood leukocytes (×109/L) | 5.6 (4.8, 6.8) | 7.2 (6.6, 8.2)* | 7.7 (6.2, 9.2)* | 6.9 (6.5, 8.4)* |
| Blood platelets (×109/L) | 245.0 (216.0., 274.0) | 252.5 (231.5, 298.5) | 267.5 (237.3, 334.5)* | 217.7 (197.3, 303.0) |
| Antitrypsin (g/L) | 1.4 (1.3, 1.5) | 1.5 (1.4, 1.7) | 1.6 (1.4, 1.7)* | 1.4 (1.3, 1.7) |
| FEV1(%) | 116.0 (108.0, 121.0) | 107.0 (95.5, 116.0)* | 72.0 (63.5, 83.3)*,** | 69.5 (63.8, 75.5)*,** |
| FEV1/FVC (%) | 80.0 (76.0, 83.0) | 76.0 (72.3, 78.8)* | 60.0 (55.8, 65.0)*,** | 59.0 (56.0, 62.8)*,** |
| Emphysema (No/Yes) | N.A. | 22/18 | 6/20 | 2/8 |
| CB (No/Yes) | N.A. | 30/10 | 19/7 | 8/2 |
| Analyte | Linear Range |
|---|---|
| Kynurenine | 0.1–10.0 µM |
| Kynurenic acid | 0.01–0.5 µM |
| Serotonin | 0.0002–3.6 µM |
| Tryptophan | 5–5000 µM |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naz, S.; Bhat, M.; Ståhl, S.; Forsslund, H.; Sköld, C.M.; Wheelock, Å.M.; Wheelock, C.E. Dysregulation of the Tryptophan Pathway Evidences Gender Differences in COPD. Metabolites 2019, 9, 212. https://doi.org/10.3390/metabo9100212
Naz S, Bhat M, Ståhl S, Forsslund H, Sköld CM, Wheelock ÅM, Wheelock CE. Dysregulation of the Tryptophan Pathway Evidences Gender Differences in COPD. Metabolites. 2019; 9(10):212. https://doi.org/10.3390/metabo9100212
Chicago/Turabian StyleNaz, Shama, Maria Bhat, Sara Ståhl, Helena Forsslund, C. Magnus Sköld, Åsa M. Wheelock, and Craig E. Wheelock. 2019. "Dysregulation of the Tryptophan Pathway Evidences Gender Differences in COPD" Metabolites 9, no. 10: 212. https://doi.org/10.3390/metabo9100212
APA StyleNaz, S., Bhat, M., Ståhl, S., Forsslund, H., Sköld, C. M., Wheelock, Å. M., & Wheelock, C. E. (2019). Dysregulation of the Tryptophan Pathway Evidences Gender Differences in COPD. Metabolites, 9(10), 212. https://doi.org/10.3390/metabo9100212