Feed Restriction Reveals Distinct Serum Metabolome Profiles in Chickens Divergent in Feed Efficiency Traits

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Design and Determination of Feed Efficiency
2.3. Blood Leukocyte Counts and Clinical Biochemistry
2.4. Serum Metabolomics
2.5. Statistical Analysis
3. Results
3.1. Restrictive Feeding Improves Residual Feed Intake
3.2. Serum Metabolome
3.3. Serum Biochemistry and White Blood Cell Counts
3.4. Supervised Data Integration: Serum Predictor Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bottje, W.G.; Carstens, G.E. Association of mitochondrial function and feed efficiency in poultry and livestock species. J. Anim. Sci. 2009, 87, E48–E63. [Google Scholar] [CrossRef] [PubMed]
- Siegerstetter, S.-C.; Schmitz-Esser, S.; Magowan, E.; Wetzels, S.U.; Zebeli, Q.; Lawlor, P.G.; O’Connell, N.E.; Metzler-Zebeli, B.U. Intestinal microbiota profiles associated with low and high residual feed intake in chickens across two geographical locations. PLoS ONE 2017, 12, e0187766. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Magowan, E.; Hollmann, M.; Ball, M.E.E.; Molnár, A.; Witter, K.; Ertl, R.; Hawken, R.J.; Lawlor, P.G.; O’Connell, N.E.; et al. Differences in intestinal size, structure, and function contributing to feed efficiency in broiler chickens reared at geographically distant locations. Poult. Sci. 2018, 97, 5781–591. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Magowan, E.; Hollmann, M.; Ball, M.E.E.; Molnár, A.; Lawlor, P.G.; Hawken, R.J.; O’Connell, N.E.; Zebeli, Q. Assessing serum metabolite profiles as predictors for feed efficiency in broiler chickens reared at geographically distant locations. Br. Poult. Sci. 2017, 58, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Reyer, H.; Metzler-Zebeli, B.U.; Trakooljul, N.; Oster, M.; Muráni, E.; Ponsuksili, S.; Hadlich, F.; Wimmers, K. Transcriptional shifts account for divergent resource allocation in feed efficient broiler chickens. Sci. Rep. 2018, 8, 12903. [Google Scholar] [CrossRef] [PubMed]
- Van der Klein, S.A.S.; Silva, F.A.; Kwakkel, R.P.; Zuidhof, M.J. The effect of quantitative feed restriction on allometric growth in broilers. Poult. Sci. 2016, 96, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Ernest, B.; Gooding, J.R.; Das, S.; Saxton, A.M.; Simon, J.; Dupont, J.; Métayer-Coustard, S.; Campagna, S.R.; Voy, B.H. Transcriptomic and metabolomic profiling of chicken adipose tissue in response to insulin neutralization and fasting. BMC Genomics 2012, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Siegerstetter, S.-C.; Petri, R.M.; Magowan, E.; Lawlor, P.G.; Zebeli, Q.; O’Connell, N.E.; Metzler-Zebeli, B.U. Feed restriction modulates the fecal microbiota composition, nutrient retention and feed efficiency in chickens divergent in residual feed intake. Front. Microbiol. 2018, 9, 2698. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Molnár, A.; Hollmann, M.; Magowan, E.; Hawken, R.J.; Lawlor, P.G.; Zebeli, Q. Comparison of growth performance and excreta composition in broiler chickens when ranked according to various feed efficiency metrics. J. Anim. Sci. 2016, 94, 2890–2899. [Google Scholar] [CrossRef] [PubMed]
- Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Dis. 1983, 27, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Eberspächer, E.; Grüll, D.; Kowalczyk, L.; Molnar, T.; Zebeli, Q. Enzymatically modified starch ameliorates postprandial serum triglycerides and lipid metabolome in growing pigs. PLoS ONE 2015, 10, e0130553. [Google Scholar] [CrossRef] [PubMed]
- Lê Cao, K.A.; Costello, M.E.; Lakis, V.A.; Bartolo, F.; Chua, X.Y.; Brazeilles, R.; Rondeau, P. MixMC: A multivariate statistical framework to gain insight into microbial communities. PLoS ONE 2016, 11, e0160169. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Saneyasu, T.; Tsuchii, N.; Nakano, Y.; Kitashiro, A.; Tsuchihashi, T.; Shindo, H.; Honda, K.; Kamisoyama, H. Effects of short-term fasting on the Akt-mediated pathway involved in protein metabolism in chicken skeletal muscle. Domest. Anim. Endocrinol. 2017, 61, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.R.; Li, H.; Emmerson, D.A.; Webb, K.E., Jr.; Wong, E.A. Dietary protein quality and feed restriction influence abundance of nutrient transporter mRNA in the small intestine of broiler chicks. J. Nutr. 2008, 138, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Yaman, M.A.; Kita, K.; Okumura, J. Different responses of protein synthesis to refeeding in various muscles of fasted chicks. Br. Poult. Sci. 2000, 41, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fragkos, K.C.; Forbes, A. Citrulline as a marker of intestinal function and absorption in clinical settings: A systematic review and meta-analysis. United Eur. Gastroenterol. J. 2018, 6, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Siegerstetter, S.-C.; Magowan, E.; Lawlor, P.G.; Petri, R.M.; O’Connell, N.E.; Zebeli, Q. Feed restriction modifies intestinal microbiota-host mucosal networking in chickens divergent in residual feed intake. mSystems 2019. [Google Scholar] [CrossRef] [PubMed]
- Knarreborg, A.; Engberg, R.M.; Jensen, S.K.; Jensen, B.B. Quantitative determination of bile salt hydrolase activity in bacteria isolated from the small intestine of chickens. Appl. Environ. Microbiol. 2002, 68, 6425–6428. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.W.; Lassiter, K.; Piekarski-Welsher, A.; Dridi, S.; Reverter-Gomez, A.; Hudson, N.J.; Bottje, W.G. Proteomics of breast muscle tissue associated with the phenotypic expression of feed efficiency within a pedigree male broiler line: I. Highlight on mitochondria. PLoS ONE 2016, 11, e0155679. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched chain amino acids: Beyond nutrition metabolism. Int. J. Mol. Sci. 2018, 19, E954. [Google Scholar] [CrossRef] [PubMed]
- Bottje, W.; Kong, B.W. Cell Biology Symposium: Feed efficiency: Mitochondrial function to global gene expression. J. Anim. Sci. 2013, 91, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P.; Garrett, M.R.; Perry, G.; Smith, M.A. Carnosine: A versatile antioxidant and antiglycating agent. Sci. Aging Knowl. Environ. 2005, 18, pe12. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Granda, A.; Damms-Machado, A.; Basrai, M.; Bischoff, S.C. Changes in plasma acylcarnitine and lysophosphatidylcholine levels following a high-fructose diet: A targeted metabolomics study in healthy women. Nutrients 2018, 10, E1254. [Google Scholar] [CrossRef] [PubMed]
- Bottje, W.G.; Lassiter, K.; Dridi, S.; Hudson, N.; Kong, B.W. Enhanced expression of proteins involved in energy production and transfer in breast muscle of pedigree male broilers exhibiting high feed efficiency. Poult. Sci. 2017, 96, 2454–2458. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.; Sassolas, A.; Guichardant, M.; Cugnet-Anceau, C.; Moret, M.; Moulin, P.; Lagarde, M.; Calzada, C. LDL from obese patients with the metabolic syndrome show increased lipid peroxidation and activate platelets. Diabetologia 2011, 54, 2931–2940. [Google Scholar] [CrossRef] [PubMed]
- Murgia, A.; Hinz, D.; Liggi, S.; Denes, J.; Hall, Z.; West, J.; Santoru, M.L.; Piras, C.; Manis, C.; Usai, P.; et al. Italian cohort of patients affected by inflammatory bowel disease is characterised by variation in glycerophospholipid, free fatty acids and amino acid levels. Metabolomics 2018, 14, 140. [Google Scholar] [CrossRef]
- Floegel, A.; Stefan, N.; Yu, Z.; Mühlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Häring, H.U.; Hrabe de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2012, 62, 639–648. [Google Scholar] [CrossRef] [PubMed]



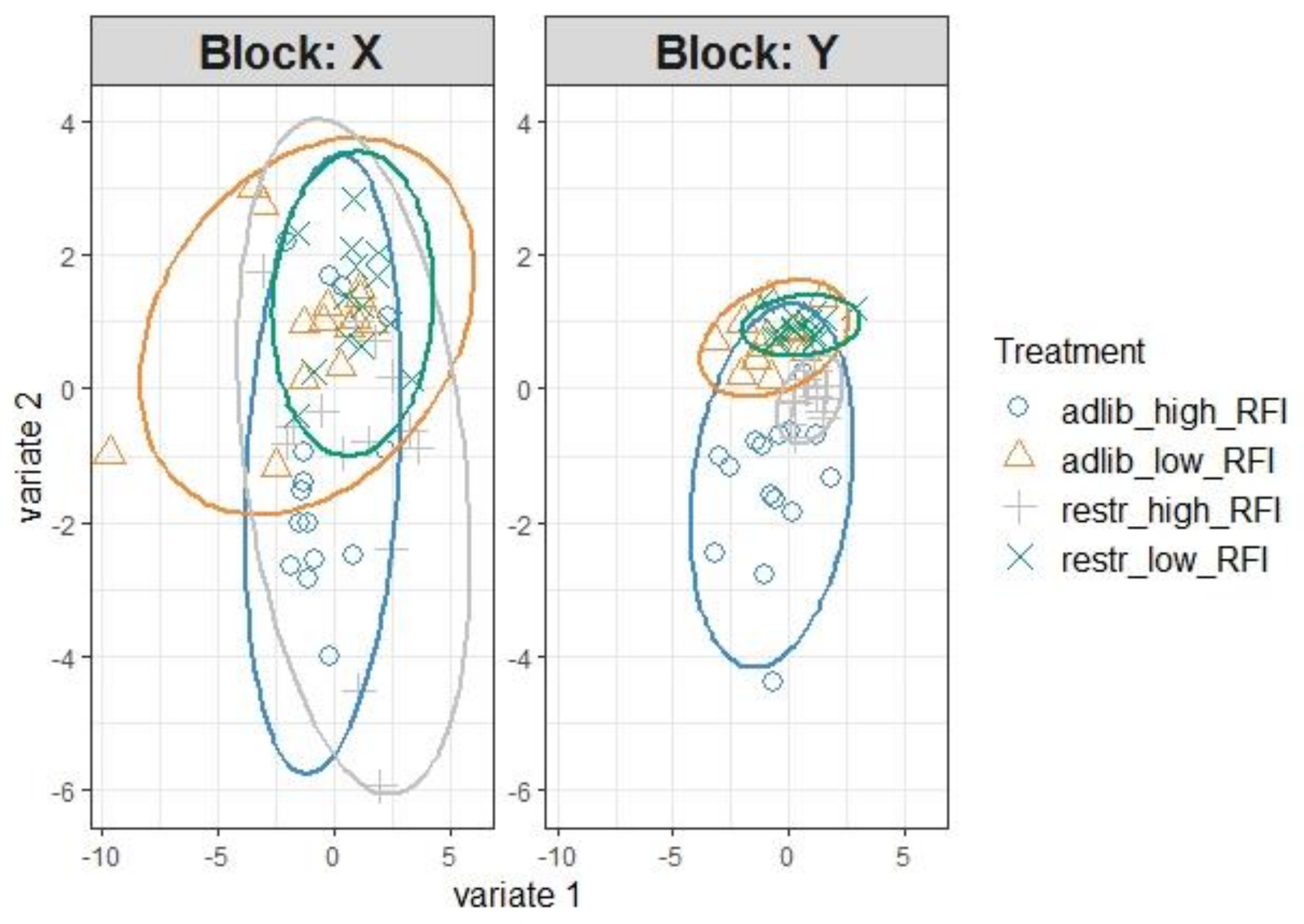{kind=link}
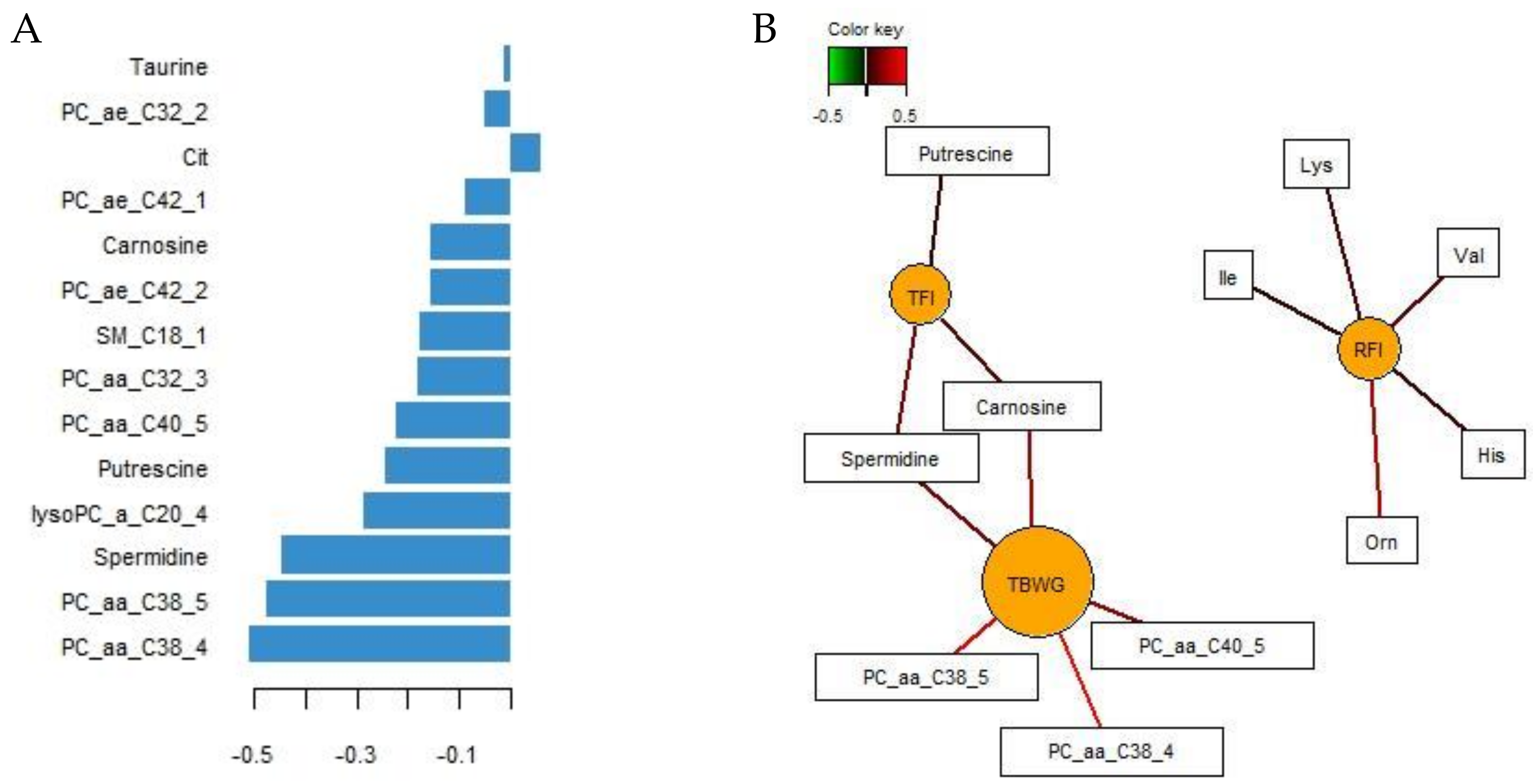{kind=link}
| Ad Libitum Feeding | Restrictive Feeding | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | Low RFI | High RFI | Low RFI | High RFI | SEM | FL | RFI | FL × RFI |
| Amino Acids | ||||||||
| Asparagine | 108.7 | 86.5 | 133.2 | 153.6 | 15.27 | 0.004 | 0.952 | 0.169 |
| Citrulline | 8.1 | 8.7 | 12.0 | 13.4 | 1.03 | <0.001 | 0.345 | 0.710 |
| Glycine | 555.4 ab | 527.6 ab | 481.8 b | 585.9 a | 31.87 | 0.810 | 0.233 | 0.042 |
| Histidine | 91.6 | 111.5 | 80.6 | 127.2 | 6.99 | 0.733 | <0.001 | 0.062 |
| Isoleucine | 124.9 | 145.2 | 120.0 | 158.3 | 7.19 | 0.570 | <0.001 | 0.215 |
| Leucine | 292.8 | 316.2 | 287.9 | 344.7 | 13.27 | 0.379 | 0.004 | 0.214 |
| Lysine | 365.8 | 427.6 | 337.6 | 429.7 | 27.57 | 0.638 | 0.007 | 0.586 |
| Ornithine | 41.0 | 56.0 | 38.2 | 65.9 | 5.15 | 0.495 | <0.001 | 0.228 |
| Proline | 446.2 | 474.5 | 466.5 | 573.8 | 20.88 | 0.006 | 0.002 | 0.064 |
| Serine | 669.1 | 714.5 | 701.3 | 848.0 | 33.56 | 0.017 | 0.006 | 0.137 |
| Taurine | 124.3 b | 169.8 a | 121.4 b | 105.1 b | 13.28 | 0.014 | 0.275 | 0.024 |
| Threonine | 464.8 | 536.3 | 483.0 | 566.4 | 28.67 | 0.403 | 0.009 | 0.835 |
| Tryptophan | 85.6 | 87.2 | 83.8 | 94.8 | 3.29 | 0.378 | 0.059 | 0.159 |
| Tyrosine | 265.8 | 219.1 | 257.8 | 241.5 | 13.86 | 0.606 | 0.027 | 0.277 |
| Valine | 199.5 | 232.4 | 188.2 | 257.1 | 11.49 | 0.566 | <0.001 | 0.123 |
| Biogenic Amines | ||||||||
| Carnosine | 21.6 | 16.6 | 15.8 | 12.5 | 1.74 | 0.006 | 0.023 | 0.622 |
| Methionine sulphoxide | 11.8 | 12.5 | 11.6 | 13.2 | 0.59 | 0.726 | 0.054 | 0.433 |
| Sarcosine | 18.3 | 21.6 | 17.7 | 22.2 | 1.20 | 0.987 | 0.002 | 0.633 |
| Symmetric dimethylarginine | 0.88 a | 0.80 ab | 0.78 b | 0.84 ab | 0.03 | 0.348 | 0.662 | 0.040 |
| Spermidine | 0.36 | 0.31 | 0.25 | 0.26 | 0.04 | 0.067 | 0.600 | 0.436 |
| 4-Hydroxyproline | 155.0 | 130.0 | 157.7 | 172.0 | 10.50 | 0.038 | 0.615 | 0.067 |
| Ad Libitum Feeding | Restrictive Feeding | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite 1 | Low RFI | High RFI | Low RFI | High RFI | SEM | FL | RFI | FL × RFI |
| Hexadecanoylcarnitine | 0.022 | 0.024 | 0.019 | 0.022 | 0.001 | 0.094 | 0.028 | 0.803 |
| LysoPC a C16:1 | 1.00 | 1.19 | 1.26 | 1.54 | 0.07 | <0.001 | 0.002 | 0.506 |
| LysoPC a C17:0 | 0.22 | 0.21 | 0.20 | 0.19 | 0.01 | 0.028 | 0.461 | 0.930 |
| LysoPC a C18:0 | 23.6 | 27.6 | 25.5 | 28.1 | 1.25 | 0.362 | 0.012 | 0.558 |
| LysoPC a C18:1 | 7.65 | 9.10 | 9.01 | 11.05 | 0.47 | <0.001 | <0.001 | 0.534 |
| LysoPC a C18:2 | 13.1 | 14.0 | 13.9 | 15.3 | 0.69 | 0.141 | 0.105 | 0.705 |
| LysoPC a C20:3 | 1.16 | 1.48 | 1.35 | 1.74 | 0.10 | 0.025 | <0.001 | 0.756 |
| LysoPC a C20:4 | 4.90 | 4.88 | 4.21 | 4.12 | 0.32 | 0.029 | 0.862 | 0.923 |
| LysoPC a C26:0 | 0.13 | 0.10 | 0.08 | 0.07 | 0.02 | 0.013 | 0.182 | 0.326 |
| LysoPC a C26:1 | 0.098 | 0.062 | 0.050 | 0.045 | 0.012 | 0.010 | 0.087 | 0.202 |
| LysoPC a C28:0 | 0.19 | 0.16 | 0.14 | 0.12 | 0.02 | 0.026 | 0.172 | 0.636 |
| LysoPC a C28:1 | 0.18 | 0.13 | 0.10 | 0.08 | 0.02 | 0.005 | 0.115 | 0.334 |
| Sphingomyelin C24:0 | 12.0 | 13.6 | 13.2 | 15.0 | 0.733 | 0.090 | 0.022 | 0.866 |
| Ad Libitum Feeding | Restrictive Feeding | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite 1 | Low RFI | High RFI | Low RFI | High RFI | SEM | FL | RFI | FL × RFI |
| PC aa C24:0 | 0.14 | 0.10 | 0.07 | 0.06 | 0.017 | 0.003 | 0.147 | 0.371 |
| PC aa C30:2 | 0.079 | 0.050 | 0.039 | 0.029 | 0.011 | 0.006 | 0.076 | 0.371 |
| PC aa C32:0 | 26.7 | 26.3 | 27.7 | 28.0 | 1.452 | <0.001 | 0.967 | 0.828 |
| PC aa C32:1 | 8.6 | 10.2 | 12.5 | 16.9 | 1.160 | <0.001 | 0.012 | 0.224 |
| PC aa C32:3 | 0.78 | 0.70 | 0.67 | 0.67 | 0.035 | 0.046 | 0.204 | 0.274 |
| PC aa C34:1 | 182.8 | 203.8 | 225.9 | 268.1 | 12.236 | <0.001 | 0.013 | 0.390 |
| PC aa C36:0 | 2.6 | 3.0 | 2.9 | 3.6 | 0.197 | 0.036 | 0.013 | 0.487 |
| PC aa C36:1 | 109.4 | 134.9 | 135.1 | 170.1 | 9.242 | 0.002 | 0.002 | 0.611 |
| PC aa C36:3 | 104.3 | 111.8 | 114.4 | 129.0 | 5.596 | 0.018 | 0.053 | 0.525 |
| PC aa C36:5 | 10.3 | 11.5 | 10.6 | 11.9 | 0.574 | 0.580 | 0.037 | 0.991 |
| PC aa C38:3 | 77.4 | 91.7 | 83.1 | 98.3 | 5.038 | 0.227 | 0.005 | 0.939 |
| PC aa C38:4 | 268.3 | 258.6 | 233.8 | 229.9 | 10.030 | 0.003 | 0.505 | 0.773 |
| PC aa C38:5 | 59.3 | 58.6 | 52.2 | 54.0 | 2.260 | 0.012 | 0.808 | 0.568 |
| PC aa C40:5 | 23.2 | 21.1 | 19.8 | 17.6 | 1.052 | 0.002 | 0.048 | 0.937 |
| PC aa C42:4 | 0.75 | 0.65 | 0.69 | 0.61 | 0.032 | 0.125 | 0.007 | 0.803 |
| PC aa C42:5 | 0.68 | 0.61 | 0.64 | 0.57 | 0.031 | 0.149 | 0.022 | 0.970 |
| PC aa C42:6 | 0.75 | 0.67 | 0.71 | 0.65 | 0.033 | 0.388 | 0.045 | 0.730 |
| PC ae C30:0 | 0.17 | 0.16 | 0.15 | 0.15 | 0.007 | 0.009 | 0.235 | 0.260 |
| PC ae C30:1 | 0.30 | 0.20 | 0.17 | 0.14 | 0.038 | 0.013 | 0.095 | 0.433 |
| PC ae C32:2 | 0.32 | 0.29 | 0.26 | 0.25 | 0.012 | 0.001 | 0.073 | 0.459 |
| PC ae C36:4 | 17.2 | 16.0 | 14.4 | 13.5 | 1.307 | 0.050 | 0.411 | 0.901 |
| PC ae C38:0 | 1.58 | 1.71 | 1.47 | 1.70 | 0.080 | 0.468 | 0.030 | 0.503 |
| PC ae C40:1 | 1.25 | 1.40 | 1.20 | 1.45 | 0.067 | 0.968 | 0.004 | 0.432 |
| PC ae C40:4 | 2.97 | 2.75 | 2.61 | 2.38 | 0.166 | 0.034 | 0.177 | 0.959 |
| PC ae C42:3 | 0.32 | 0.39 | 0.34 | 0.44 | 0.024 | 0.131 | 0.001 | 0.578 |
| PC ae C44:4 | 0.11 | 0.09 | 0.09 | 0.09 | 0.006 | 0.011 | 0.163 | 0.174 |
| Ad Libitum Feeding | Restrictive Feeding | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | Low RFI | High RFI | Low RFI | High RFI | SEM | FL | RFI | FL × RFI |
| Uric acid (mg/dL) | 1.20 | 1.50 | 1.11 | 1.11 | 0.10 | 0.024 | 0.284 | 0.289 |
| Cholesterol (mg/dL) | 138 | 151 | 146 | 162 | 4.9 | 0.060 | 0.004 | 0.750 |
| Lymphocytes (%) | 84.5 | 83.1 | 87.1 | 88.4 | 1.85 | 0.038 | 0.966 | 0.476 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metzler-Zebeli, B.U.; Siegerstetter, S.-C.; Magowan, E.; Lawlor, P.G.; O’Connell, N.E.; Zebeli, Q. Feed Restriction Reveals Distinct Serum Metabolome Profiles in Chickens Divergent in Feed Efficiency Traits. Metabolites 2019, 9, 38. https://doi.org/10.3390/metabo9020038
Metzler-Zebeli BU, Siegerstetter S-C, Magowan E, Lawlor PG, O’Connell NE, Zebeli Q. Feed Restriction Reveals Distinct Serum Metabolome Profiles in Chickens Divergent in Feed Efficiency Traits. Metabolites. 2019; 9(2):38. https://doi.org/10.3390/metabo9020038
Chicago/Turabian StyleMetzler-Zebeli, Barbara U., Sina-Catherine Siegerstetter, Elizabeth Magowan, Peadar G. Lawlor, Niamh E. O’Connell, and Qendrim Zebeli. 2019. "Feed Restriction Reveals Distinct Serum Metabolome Profiles in Chickens Divergent in Feed Efficiency Traits" Metabolites 9, no. 2: 38. https://doi.org/10.3390/metabo9020038
APA StyleMetzler-Zebeli, B. U., Siegerstetter, S. -C., Magowan, E., Lawlor, P. G., O’Connell, N. E., & Zebeli, Q. (2019). Feed Restriction Reveals Distinct Serum Metabolome Profiles in Chickens Divergent in Feed Efficiency Traits. Metabolites, 9(2), 38. https://doi.org/10.3390/metabo9020038