Ozone and Wounding Stresses Differently Alter the Temporal Variation in Formylated Phloroglucinols in Eucalyptus globulus Leaves

 ,
, 

Abstract
:1. Introduction
2. Results
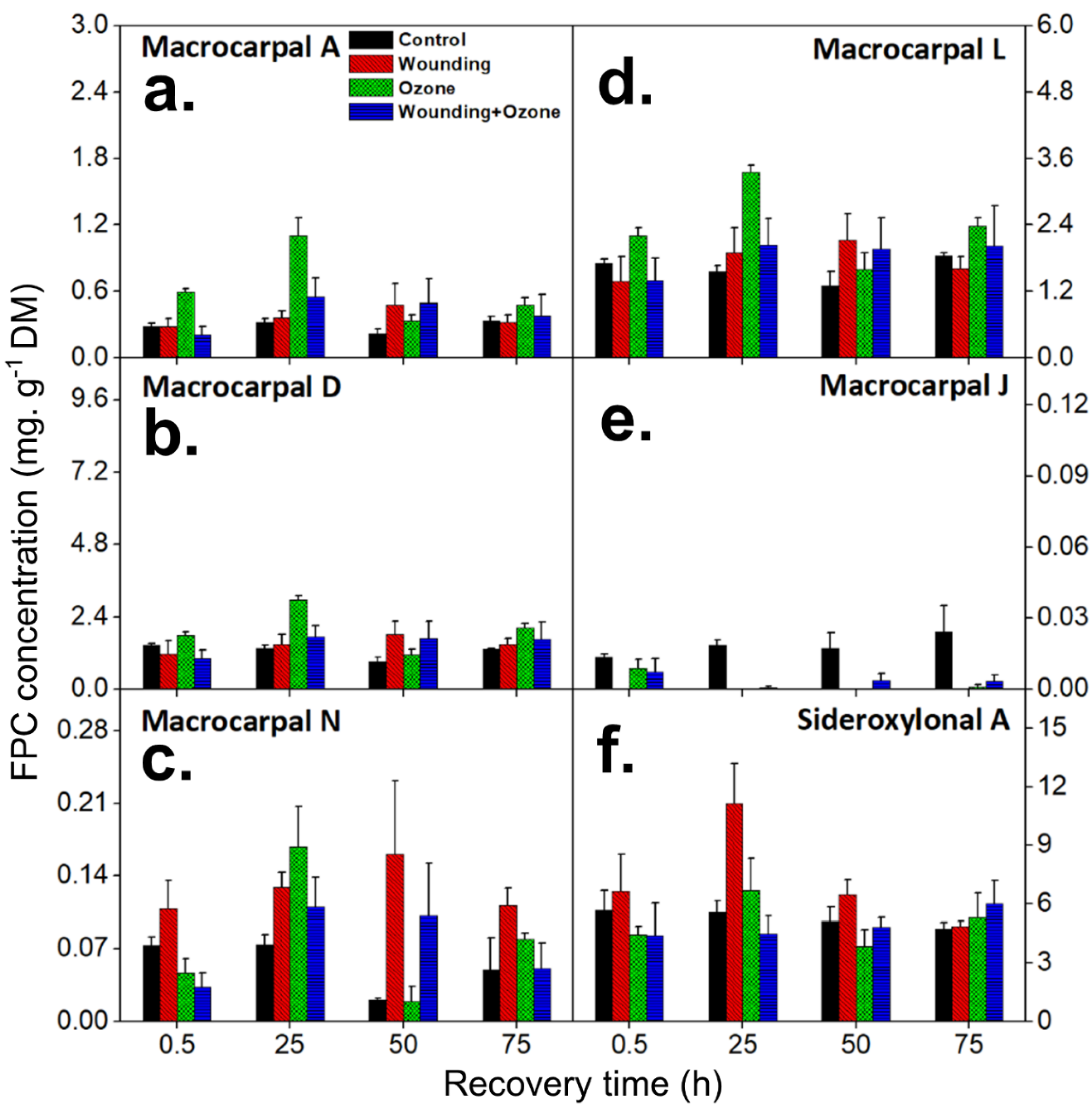
2.1. Wounding and O3 Treatments Alter Individual FPC Concentrations in E. globulus Leaves
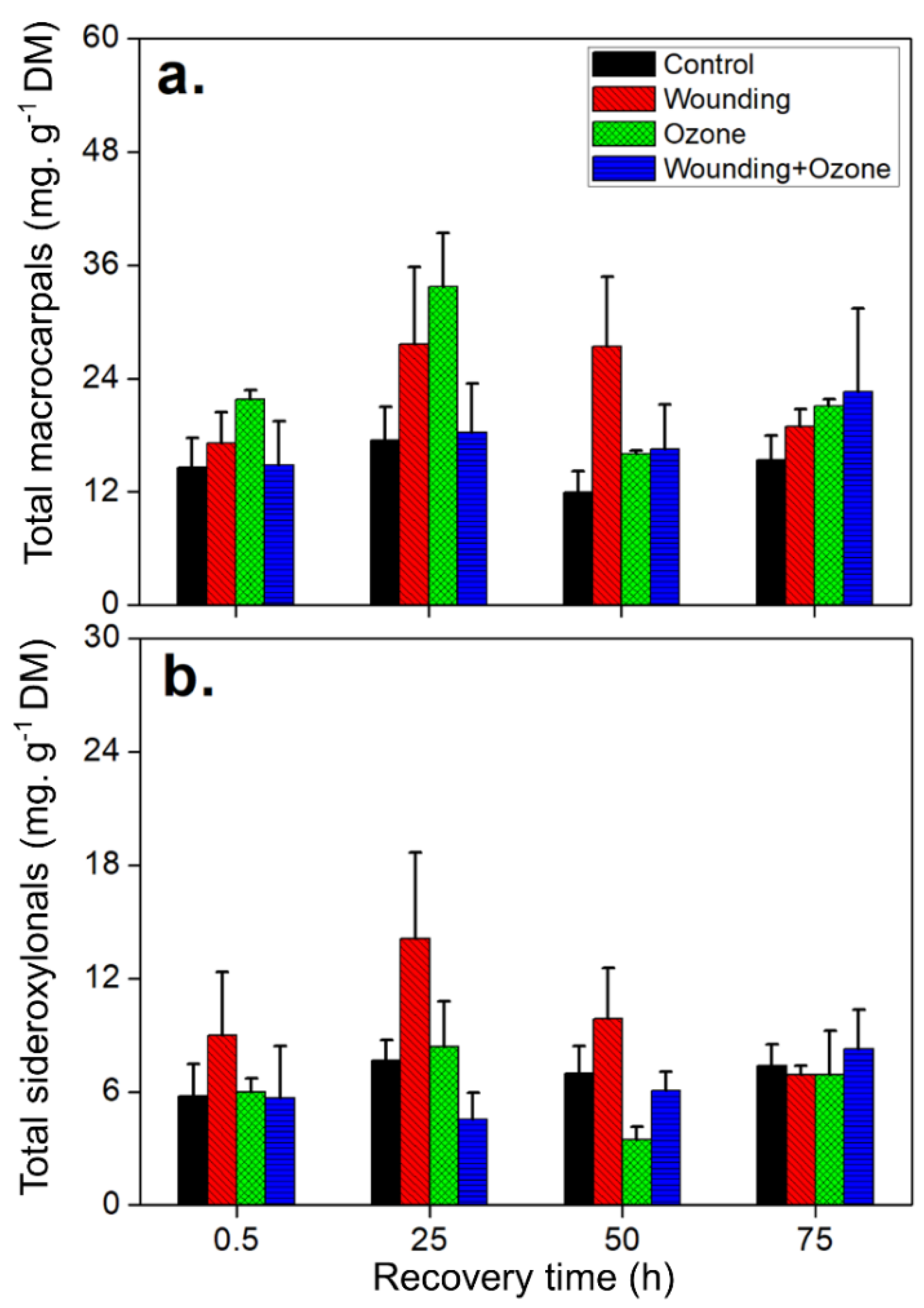
2.2. Wounding and O3 Treatments Affect Total FPC and Phenolic Content in E. globulus Leaves
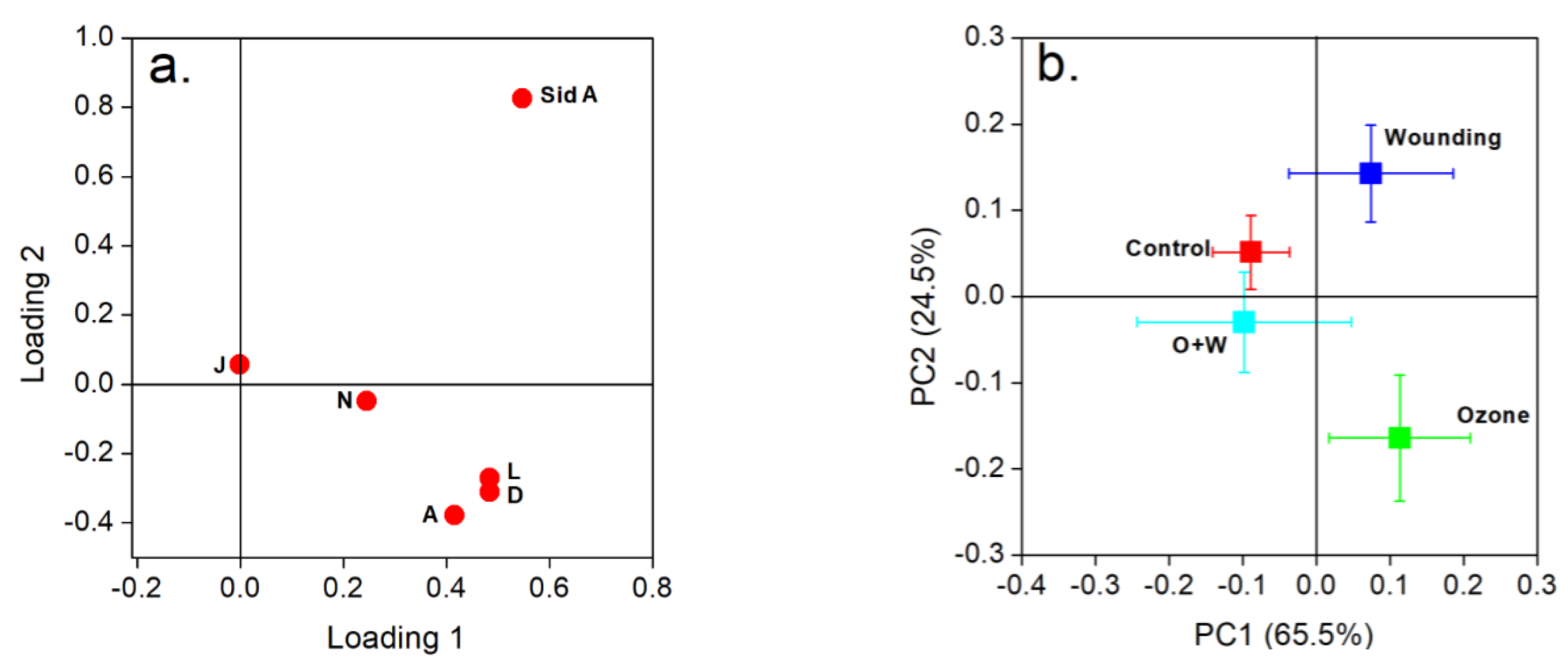
2.3. Wounding and O3 Treatments Alter the FPC Composition in E. globulus Leaves
3. Discussion
3.1. FPCs Responses to O3 Stress
3.2. FPCs Responses to Wounding Stress
3.3. FPCs Responses to Combination of O3 and Wounding
3.4. Correlations of FPCs with Other Specialized Metabolites
4. Material and Methods
4.1. Plant Materials
4.2. Wounding and O3 Treatments
4.3. FPCs Extraction and UHPLC-DAD-ESI-Q-TOF-MS/MS Analysis
4.4. Total Phenolic Contents Extraction and Determination
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sillman, S. The relation between ozone, NOx and hydrocarbons in urban and polluted rural environments. Atmos. Environ. 1999, 33, 1821–1845. [Google Scholar] [CrossRef]
- Carter, W.P. Development of ozone reactivity scales for volatile organic compounds. Air Waste 1994, 44, 881–899. [Google Scholar] [CrossRef]
- Young, P.; Archibald, A.; Bowman, K.; Lamarque, J.-F.; Naik, V.; Stevenson, D.; Tilmes, S.; Voulgarakis, A.; Wild, O.; Bergmann, D. Pre-industrial to end 21st century projections of tropospheric ozone from the Atmospheric Chemistry and Climate Model Intercomparison Project (ACCMIP). Atmos. Chem. Phys. 2013, 13, 2063–2090. [Google Scholar] [CrossRef] [Green Version]
- Wohlgemuth, H.; Mittelstrass, K.; Kschieschan, S.; Bender, J.; Weigel, H.J.; Overmyer, K.; Kangasjärvi, J.; Sandermann, H.; Langebartels, C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant Cell Environ. 2002, 25, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Langebartels, C.; Wohlgemuth, H.; Kschieschan, S.; Grün, S.; Sandermann, H. Oxidative burst and cell death in ozone-exposed plants. Plant Physiol. Biochem. 2002, 40, 567–575. [Google Scholar] [CrossRef]
- Kangasjärvi, J.; Jaspers, P.; Kollist, H. Signalling and cell death in ozone-exposed plants. Plant Cell Environ. 2005, 28, 1021–1036. [Google Scholar] [CrossRef] [Green Version]
- Kanagendran, A.; Pazouki, L.; Niinemets, Ü. Differential regulation of volatile emission from Eucalyptus globulus leaves upon single and combined ozone and wounding treatments through recovery and relationships with ozone uptake. Environ. Exp. Bot. 2018, 145, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Shadkami, F.; Helleur, R.; Cox, R. Profiling secondary metabolites of needles of ozone-fumigated white pine (Pinus strobus) clones by thermally assisted hydrolysis/methylation GC/MS. J. Chem. Ecol. 2007, 33, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Huang, W.; Chen, W.; Dong, T.; Liu, C.; Chen, Z.; Xu, S.; Ruan, Y. Changes of main secondary metabolites in leaves of Ginkgo biloba in response to ozone fumigation. J. Environ. Sci. 2009, 21, 199–203. [Google Scholar] [CrossRef]
- Mikkelsen, B.L.; Olsen, C.E.; Lyngkjær, M.F. Accumulation of secondary metabolites in healthy and diseased barley, grown under future climate levels of CO2, ozone and temperature. Phytochemistry 2015, 118, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M.; Pellegrini, E.; D’Angiolillo, F.; Petersen, M.; Nali, C.; Pistelli, L.; Lorenzini, G. Ozone-elicited secondary metabolites in shoot cultures of Melissa officinalis L. Plant Cell Tissue Organ Cult. 2015, 120, 617–629. [Google Scholar] [CrossRef]
- Kanagendran, A.; Pazouki, L.; Bichele, R.; Külheim, C.; Niinemets, Ü. Temporal regulation of terpene synthase gene expression in Eucalyptus globulus leaves upon ozone and wounding stresses: Relationships with stomatal ozone uptake and emission responses. Environ. Exp. Bot. 2018, 155, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Fares, S.; Park, J.-H.; Ormeno, E.; Gentner, D.R.; McKay, M.; Loreto, F.; Karlik, J.; Goldstein, A.H. Ozone uptake by citrus trees exposed to a range of ozone concentrations. Atmos. Environ. 2010, 44, 3404–3412. [Google Scholar] [CrossRef]
- Fares, S.; Oksanen, E.; Lännenpää, M.; Julkunen-Tiitto, R.; Loreto, F. Volatile emissions and phenolic compound concentrations along a vertical profile of Populus nigra leaves exposed to realistic ozone concentrations. Photosynth. Res. 2010, 104, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Saviranta, N.M.; Julkunen-Tiitto, R.; Oksanen, E.; Karjalainen, R.O. Leaf phenolic compounds in red clover (Trifolium pratense L.) induced by exposure to moderately elevated ozone. Environ. Pollut. 2010, 158, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Loponen, J.; Pihlaja, K.; Oksanen, E. Effects of long-term open-field ozone exposure on leaf phenolics of European silver birch (Betula pendula Roth). J. Chem. Ecol. 2001, 27, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Kännaste, A.; Remmel, T.; Niinemets, Ü. Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses. Environ. Exp. Bot. 2014, 100, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Mild versus severe stress and BVOCs: Thresholds, priming and consequences. Trends Plant Sci. 2010, 15, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Roy, J. Response of Plants to Multiple Stresses; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Kothari, A.R.; Burnett, N.P. Herbivores alter plant–wind interactions by acting as a point mass on leaves and by removing leaf tissue. Ecol. Evol. 2017, 7, 6884–6893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León, J.; Rojo, E.; Sánchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.; Villarroel, C.; Ataide, L.; Dermauw, W.; Glas, J. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eschler, B.; Pass, D.; Willis, R.; Foley, W. Distribution of foliar formylated phloroglucinol derivatives amongst Eucalyptus species. Biochem. Syst. Ecol. 2000, 28, 813–824. [Google Scholar] [CrossRef]
- Foley, W.J.; Lassak, E.V. The Potential of Bioactive Constituents of Eucalyptus Foliage as Non-Wood Products from Plantations; Rural Industries Research and Development Corporation: Canberra, Australia, 2004; Publication no. 04/154. [Google Scholar]
- Moore, B.D.; Wallis, I.R.; Palá-Paúl, J.; Brophy, J.J.; Willis, R.H.; Foley, W.J. Antiherbivore chemistry of Eucalyptus--cues and deterrents for marsupial folivores. J. Chem. Ecol. 2004, 30, 1743–1769. [Google Scholar] [CrossRef] [PubMed]
- Wallis, I.; Watson, M.; Foley, W. Secondary metabolites in Eucalyptus melliodora: Field distribution and laboratory feeding choices by a generalist herbivore, the common brushtail possum. Aust. J. Zool. 2002, 50, 507–519. [Google Scholar] [CrossRef]
- McKiernan, A.B.; Hovenden, M.J.; Brodribb, T.J.; Potts, B.M.; Davies, N.W.; O’Reilly-Wapstra, J.M. Effect of limited water availability on foliar plant secondary metabolites of two Eucalyptus species. Environ. Exp. Bot. 2014, 105, 55–64. [Google Scholar] [CrossRef]
- McKiernan, A.B.; Potts, B.M.; Brodribb, T.J.; Hovenden, M.J.; Davies, N.W.; McAdam, S.A.; Ross, J.J.; Rodemann, T.; O’Reilly-Wapstra, J.M. Responses to mild water deficit and rewatering differ among secondary metabolites but are similar among provenances within Eucalyptus species. Tree Physiol. 2015, 36, 133–147. [Google Scholar] [PubMed]
- Niinemets, Ü. Uncovering the hidden facets of drought stress: Secondary metabolites make the difference. Tree Physiol. 2015, 36, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Sidana, J.; Rohilla, R.K.; Roy, N.; Barrow, R.A.; Foley, W.J.; Singh, I.P. Antibacterial sideroxylonals and loxophlebal A from Eucalyptus loxophleba foliage. Fitoterapia 2010, 81, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Alliot, J.; Gravel, E.; Larquetoux, L.; Nicolas, M.; Doris, E. Semisynthesis of macrocarpal C and analogues by selective dehydration of macrocarpal A or B. J. Nat. Prod. 2013, 76, 2346–2349. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Emura, M.; Kan, Y.; Yamada, H.; Ogawa, K.; Hamanaka, N. Macrocarpals: HIV-RTase inhibitors of Eucalyptus globulus. Tetrahedron Lett. 1992, 33, 2983–2986. [Google Scholar] [CrossRef]
- Qin, X.-J.; Yan, H.; Ni, W.; Yu, M.-Y.; Khan, A.; Liu, H.; Zhang, H.-X.; He, L.; Hao, X.-J.; Di, Y.-T. Cytotoxic meroterpenoids with rare skeletons from Psidium guajava cultivated in temperate zone. Sci. Rep. 2016, 6, 32748. [Google Scholar] [CrossRef] [PubMed]
- Eyles, A.; Davies, N.W.; Mohammed, C. Wound wood formation in Eucalyptus globulus and Eucalyptus nitens: Anatomy and chemistry. Can. J. For. Res. 2003, 33, 2331–2339. [Google Scholar] [CrossRef]
- Kim, J.-P.; Lee, I.-K.; Yun, B.-S.; Chung, S.-H.; Shim, G.-S.; Koshino, H.; Yoo, I.-D. Ellagic acid rhamnosides from the stem bark of Eucalyptus globulus. Phytochemistry 2001, 57, 587–591. [Google Scholar] [CrossRef]
- Jensen, L.M.; Wallis, I.R.; Foley, W.J. The relative concentrations of nutrients and toxins dictate feeding by a vertebrate browser, the greater glider Petauroides volans. PLoS ONE 2015, 10, e0121584. [Google Scholar] [CrossRef] [PubMed]
- Lawler, I.R.; Foley, W.J.; Eschler, B.M. Foliar concentration of a single toxin creates habitat patchiness for a marsupial folivore. Ecology 2000, 81, 1327–1338. [Google Scholar] [CrossRef]
- Moore, B.D.; Foley, W.J.; Wallis, I.R.; Cowling, A.; Handasyde, K.A. Eucalyptus foliar chemistry explains selective feeding by koalas. Biol. Lett. 2005, 1, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Sidana, J.; Bharate, S.B.; Foley, W.J. Phloroglucinol compounds of natural origin: Synthetic aspects. Nat. Prod. Rep. 2010, 27, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, L.; Filomeno, C.; Teixeira, R. Chemical variability and biological activities of Eucalyptus spp. essential oils. Molecules 2016, 21, 1671. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Delfine, S. Emission of isoprene from salt-stressed Eucalyptus globulus leaves. Plant Physiol. 2000, 123, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Chenavas, S.; Fiorini-Puybaret, C.; Joulia, P.; Larrouquet, C.; Waton, H.; Martinez, A.; Casabianca, H.; Fabre, B. New formylated phloroglucinol compounds from Eucalyptus globulus foliage. Phytochem. Lett. 2015, 11, 69–73. [Google Scholar] [CrossRef]
- Osawa, K.; Yasuda, H.; Morita, H.; Takeya, K.; Itokawa, H. Macrocarpals H, I, and J from the leaves of Eucalyptus globulus. J. Nat. Prod. 1996, 59, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Leipner, J.R.; Oxborough, K.; Baker, N.R. Primary sites of ozone-induced perturbations of photosynthesis in leaves: Identification and characterization in Phaseolus vulgaris using high resolution chlorophyll fluorescence imaging. J. Exp. Bot. 2001, 52, 1689–1696. [Google Scholar] [PubMed]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Monk, R.; Murray, F. The relative tolerance of some Eucalyptus species to ozone exposure. Water Air Soil Pollut. 1995, 85, 1405–1411. [Google Scholar] [CrossRef]
- Loreto, F.; Pinelli, P.; Manes, F.; Kollist, H. Impact of ozone on monoterpene emissions and evidence for an isoprene-like antioxidant action of monoterpenes emitted by Quercus ilex leaves. Tree Physiol. 2004, 24, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Yoshikawa, K.; Kawahara, Y.; Arihara, S.; Hashimoto, T. Aromatic compounds and their antioxidant activity of Acer saccharum. J. Nat. Med. 2011, 65, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Pass, D.M.; Foley, W.J.; Bowden, B. Vertebrate herbivory on Eucalyptus—Identification of specific feeding deterrents for common ringtail possums (Pseudocheirus peregrinus) by bioassay-guided fractionation of Eucalyptus ovata foliage. J. Chem. Ecol. 1998, 24, 1513–1527. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusia, J.; Asensio, D.; Munne-Bosch, S. Linking isoprene with plant thermotolerance, antioxidants and monoterpene emissions. Plant Cell Environ. 2005, 28, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Severino, J.F.; Stich, K.; Soja, G. Ozone stress and antioxidant substances in Trifolium repens and Centaurea jacea leaves. Environ. Pollut. 2007, 146, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, S.; Piccioni, C.; Reale, L.; Ederli, L.; Della Torre, G.; Ferranti, F. Ozone-induced cell death in tobacco cultivar Bel W3 plants. The role of programmed cell death in lesion formation. Plant Physiol. 2003, 133, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- Sgarbi, E.; Fornasiero, R.B.; Lins, A.P.; Bonatti, P.M. Phenol metabolism is differentially affected by ozone in two cell lines from grape (Vitis vinifera L.) leaf. Plant Sci. 2003, 165, 951–957. [Google Scholar] [CrossRef]
- Sharma, Y.K.; Davis, K.R. Ozone-induced expression of stress-related genes in Arabidopsis thaliana. Plant Physiol. 1994, 105, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Barta, C.; Brilli, F.; Nogues, I. On the induction of volatile organic compound emissions by plants as consequence of wounding or fluctuations of light and temperature. Plant Cell Environ. 2006, 29, 1820–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.M.; Wallis, I.R.; Marsh, K.J.; Moore, B.D.; Wiggins, N.L.; Foley, W.J. Four species of arboreal folivore show differential tolerance to a secondary metabolite. Oecologia 2014, 176, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Stapley, J.; Foley, W.; Cunningham, R.; Eschler, B. How well can common brushtail possums regulate their intake of Eucalyptus toxins? J. Comp. Physiol. B 2000, 170, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, M.; Foley, W.J.; Floyd, R.B. Role of volatile and non-volatile plant secondary metabolites in host tree selection by Christmas beetles. J. Chem. Ecol. 2011, 37, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Wallis, I.R.; Keszei, A.; Henery, M.L.; Moran, G.F.; Forrester, R.; Maintz, J.; Marsh, K.J.; Andrew, R.L.; Foley, W.J. A chemical perspective on the evolution of variation in Eucalyptus globulus. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 305–318. [Google Scholar] [CrossRef]
- Kang, H.-M.; Saltveit, M.E. Antioxidant capacity of lettuce leaf tissue increases after wounding. J. Agric. Food Chem. 2002, 50, 7536–7541. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.F.; Villarreal, J.E.; Cisneros-Zevallos, L. The increase in antioxidant capacity after wounding depends on the type of fruit or vegetable tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar] [CrossRef]
- López-Gálvez, G.; Saltveit, M.; Cantwell, M. Wound-induced phenylalanine ammonia lyase activity: Factors affecting its induction and correlation with the quality of minimally processed lettuces. Postharvest Biol. Technol. 1996, 9, 223–233. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Luo, W.; Tanaka, G.; Konishi, Y.; Matsuura, H.; Takahashi, K. Wounding stress induces phenylalanine ammonia lyases, leading to the accumulation of phenylpropanoids in the model liverwort Marchantia polymorpha. Phytochemistry 2018, 155, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Alexieva, V.; Ivanov, S.; Sergiev, I.; Karanov, E. Interaction between stresses. Bulg. J. Plant Physiol. 2003, 29, 1–17. [Google Scholar]
- Niinemets, Ü.; Kahru, A.; Mander, Ü.; Nõges, P.; Nõges, T.; Tuvikene, A.; Vasemägi, A. Interacting environmental and chemical stresses under global change in temperate aquatic ecosystems: Stress responses, adaptation, and scaling. Reg. Environ. Chang. 2017, 17, 2061–2077. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Van Doorslaer, S.; Dedonder, A.; de Blocks, M.; Callens, F. Oxidative stress in plants: EPR monitoring in DMPO-DMSO based extracts. J. Plant Physiol. 1999, 154, 132–136. [Google Scholar] [CrossRef]
- Gao, Y.; Li, G.-T.; Li, Y.; Hai, P.; Wang, F.; Liu, J.-K. Guajadials CF, four unusual meroterpenoids from Psidium guajava. Nat. Prod. Bioprospecting 2013, 3, 14–19. [Google Scholar] [CrossRef]
- Jian, Y.Q.; Huang, X.J.; Zhang, D.M.; Jiang, R.W.; Chen, M.F.; Zhao, B.X.; Wang, Y.; Ye, W.C. Guapsidial A and guadials B and C: Three new meroterpenoids with unusual skeletons from the leaves of Psidium guajava. Chem. Eur. J. 2015, 21, 9022–9027. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Wang, Y.; Jian, Y.-Q.; Huang, X.-J.; Zhang, D.-M.; Tang, Q.-F.; Jiang, R.-W.; Sun, X.-G.; Lv, Z.-P.; Zhang, X.-Q. Guadial A and psiguadials C and D, three unusual meroterpenoids from Psidium guajava. Org. Lett. 2012, 14, 5262–5265. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.-H.; Dong, Z.; Guo, Y.-Q.; Cheng, Z.-B.; Zhou, C.-J.; Yin, S. Psiguajadials A–K: Unusual Psidium Meroterpenoids as Phosphodiesterase-4 Inhibitors from the Leaves of Psidium guajava. Sci. Rep. 2017, 7, 1047. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-L.; Hsieh, K.-L.; Liu, J.-K. Guajadial: An unusual meroterpenoid from guava leaves Psidium guajava. Org. Lett. 2007, 9, 5135–5138. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Xue, J.-J.; Fan, C.-Q.; Miao, Z.-H.; Ding, J.; Yue, J.-M. Eucalyptals A− C with a new skeleton isolated from Eucalyptus globulus. Org. Lett. 2007, 9, 5549–5552. [Google Scholar] [CrossRef] [PubMed]
- Külheim, C.; Yeoh, S.H.; Wallis, I.R.; Laffan, S.; Moran, G.F.; Foley, W.J. The molecular basis of quantitative variation in foliar secondary metabolites in Eucalyptus globulus. New Phytol. 2011, 191, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- Henery, M.L.; Moran, G.F.; Wallis, I.R.; Foley, W.J. Identification of quantitative trait loci influencing foliar concentrations of terpenes and formylated phloroglucinol compounds in Eucalyptus nitens. New Phytol. 2007, 176, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Tounekti, T.; Hernández, I.; Munné-Bosch, S. Salicylic acid biosynthesis and role in modulating terpenoid and flavonoid metabolism in plant responses to abiotic stress. In Salicylic Acid; Springer: Berlin/Heidelberg, Germany, 2013; pp. 141–162. [Google Scholar]
- Close, D.C.; Davies, N.W.; Beadle, C.L. Temporal variation of tannins (galloylglucoses), flavonols and anthocyanins in leaves of Eucalyptus nitens seedlings: Implications for light attenuation and antioxidant activities. Funct. Plant Biol. 2001, 28, 269–278. [Google Scholar] [CrossRef]
- Tanaka, T.; Mikamiyama, H.; Maeda, K.; Iwata, C.; In, Y.; Ishida, T. Total synthesis of (−)-macrocarpal C. Stereoselective coupling reaction with a novel hexasubstituted benzene Cr (CO)3 complex as a biomimetic chiral benzyl cation equivalent. J. Org. Chem. 1998, 63, 9782–9793. [Google Scholar] [CrossRef]
- Copolovici, L.; Niinemets, Ü. Flooding induced emissions of volatile signalling compounds in three tree species with differing waterlogging tolerance. Plant Cell Environ. 2010, 33, 1582–1594. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.; Parbery, D.; Strauss, W. The effects of phytotoxic gases on native Australian plant species: Part 2. Acute injury due to ozone. Environ. Pollut. (1970) 1975, 9, 181–192. [Google Scholar] [CrossRef]
- Beauchamp, J.; Wisthaler, A.; Hansel, A.; Kleist, E.; Miebach, M.; Niinemets, Ü.; Schurr, U.; Wildt, J. Ozone induced emissions of biogenic VOC from tobacco: Relationships between ozone uptake and emission of LOX products. Plant Cell Environ. 2005, 28, 1334–1343. [Google Scholar] [CrossRef]
- Calfapietra, C.; Wiberley, A.E.; Falbel, T.G.; Linskey, A.R.; Mugnozza, G.S.; Karnosky, D.F.; Loreto, F.; Sharkey, T.D. Isoprene synthase expression and protein levels are reduced under elevated O3 but not under elevated CO2 (FACE) in field-grown aspen trees. Plant Cell Environ. 2007, 30, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Harley, P.C.; Niinemets, Ü. Ozone-induced foliar damage and release of stress volatiles is highly dependent on stomatal openness and priming by low-level ozone exposure in Phaseolus vulgaris. Plant Cell Environ. 2017, 40, 1984–2003. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.M.d.; Zibrandtsen, J.F.S.; Gunbilig, D.; Sørensen, M.; Cozzi, F.; Boughton, B.A.; Heskes, A.M.; Neilson, E.H.J. Quantification and Localization of Formylated Phloroglucinol Compounds (FPCs) in Eucalyptus Species. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Xu, B.; Chang, S. A comparative study on phenolic profiles and antioxidant activities of legumes as affected by extraction solvents. J. Food Sci. 2007, 72, S159–S166. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
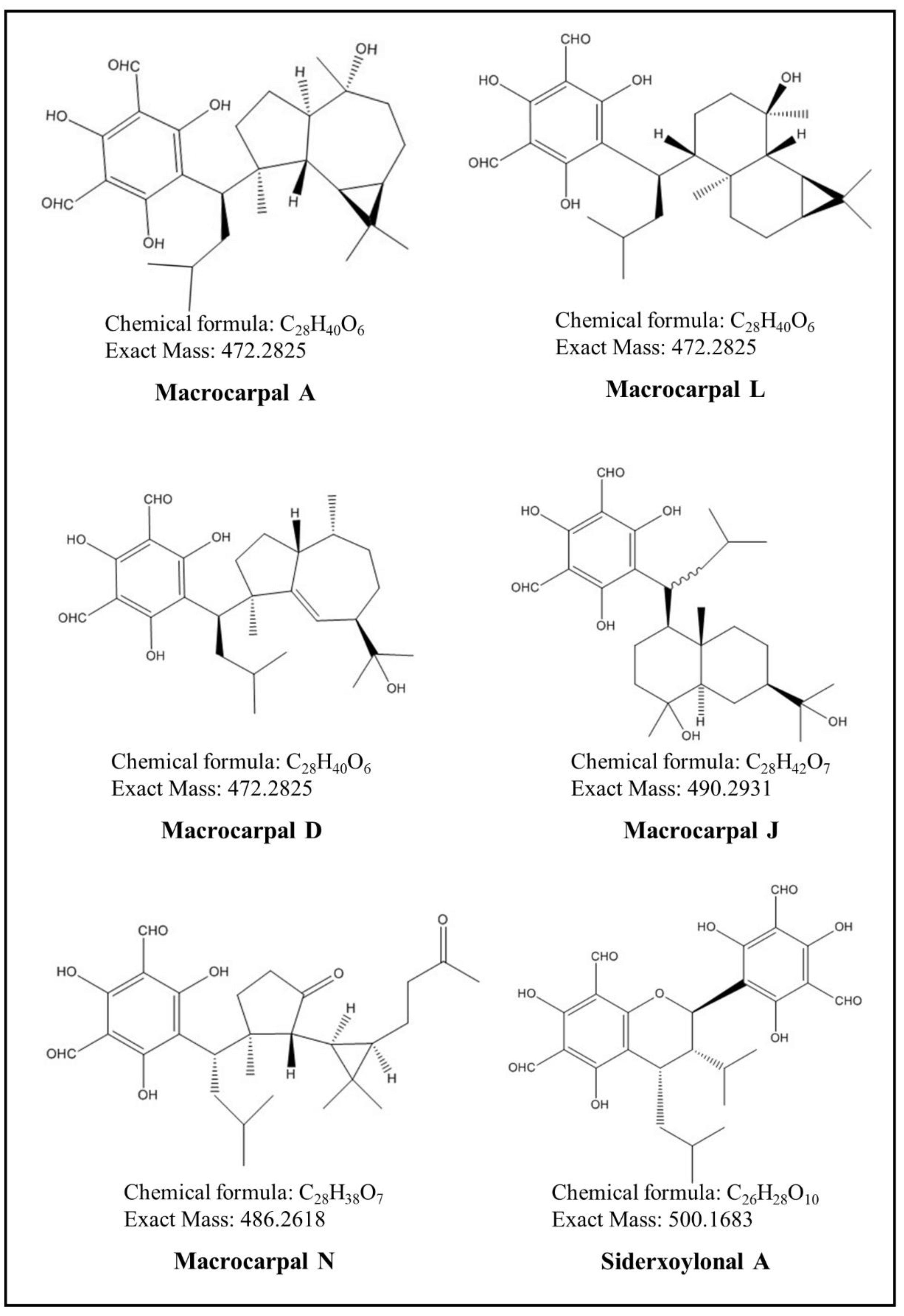
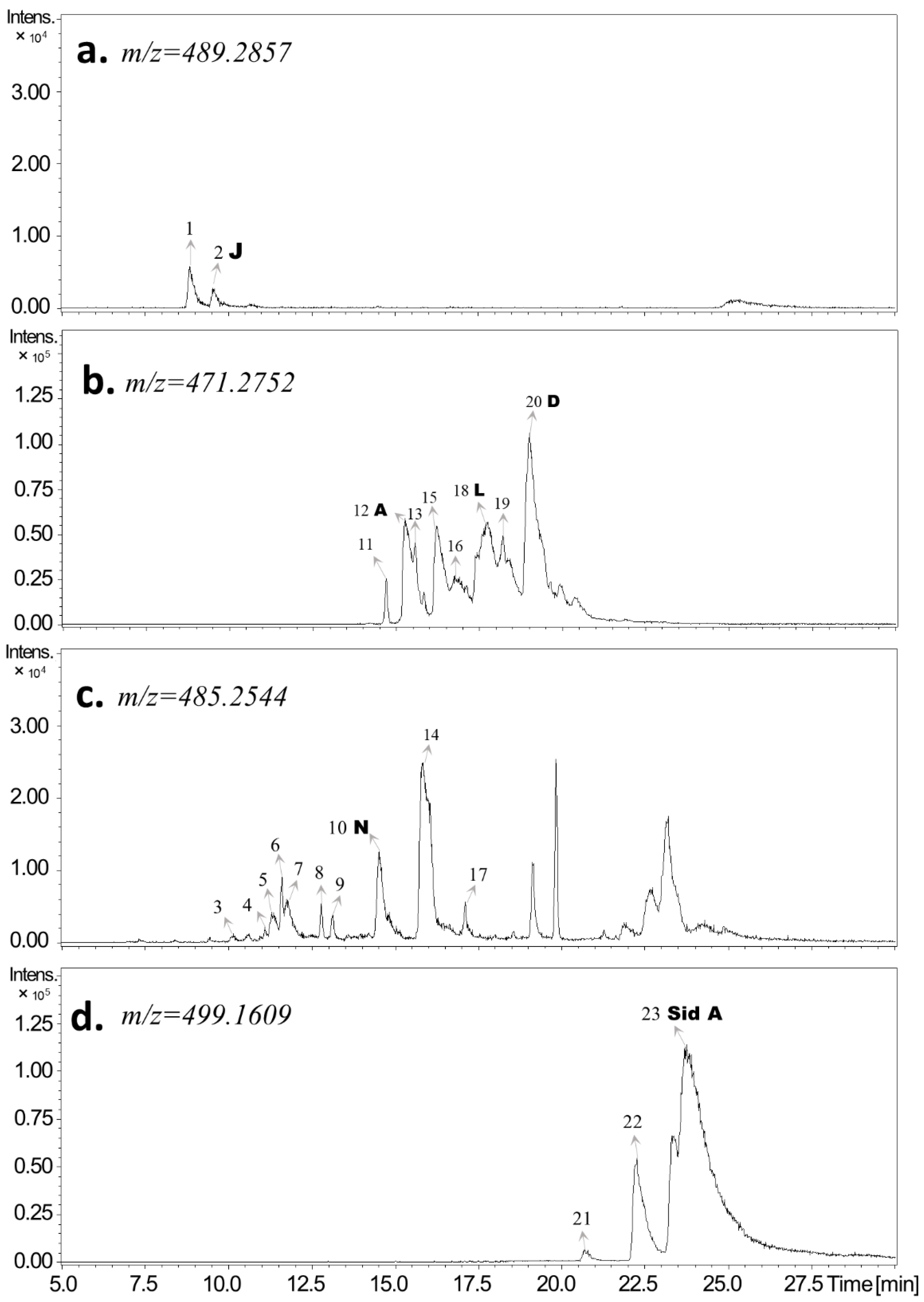
| Peak # | RT [min] | Relative Peak Area% | Calculated [M−H]− m/z | Measured [M−H]− m/z | Error (ppm) | MS/MS (Relative Intensity, in %) | Molecular Formula | Identity |
|---|---|---|---|---|---|---|---|---|
| 1 | 8.9 | 1.2 | 489.2857 | 489.2866 | 1.8 | 489 (100), 490 (35), 491 (8) | C28H42O7 | FPC01 |
| 2 | 9.6 | 0.5 | 489.2857 | 489.2864 | 1.4 | 489 (100), 490 (28), 491 (5) | C28H42O7 | FPC02 Macrocarpal J |
| 3 | 10.2 | 0.1 | 485.2544 | 485.2543 | −0.2 | 485 (100), 486 (41), 487 (14), 249 (8) | C28H38O7 | FPC03 |
| 4 | 11.1 | 0.2 | 485.2544 | 485.2547 | 0.6 | 485 (100), 486 (28), 487 (15), 249 (10) | C28H38O7 | FPC04 |
| 5 | 11.3 | 0.9 | 485.2544 | 485.2550 | 1.2 | 485 (100), 486 (23), 207 (40), 250 (13) | C28H38O7 | FPC05 |
| 6 | 11.6 | 0.9 | 485.2544 | 485.2549 | 1.0 | 485 (100), 486 (26), 487 (7), 207 (5) | C28H38O7 | FPC06 |
| 7 | 11.8 | 1.2 | 485.2544 | 485.2542 | −0.4 | 485 (100), 486 (23), 207 (28), 250 (15) | C28H38O7 | FPC07 |
| 8 | 12.8 | 0.4 | 485.2544 | 485.2547 | 0.6 | 485 (100), 486 (23), 487 (10), 207 (3) | C28H38O7 | FPC08 |
| 9 | 13.1 | 0.4 | 485.2544 | 485.2550 | 1.2 | 485 (100), 486 (40), 487 (8), 207 (10) | C28H38O7 | FPC09 |
| 10 | 14.5 | 1.7 | 485.2544 | 485.2546 | 0.4 | 485 (100), 486 (29), 207 (9), 250 (10) | C28H38O7 | FPC10 Macrocarpal N |
| 11 | 14.7 | 2.1 | 471.2752 | 471.2756 | 0.8 | 471 (100), 472 (32), 473 (4) | C28H40O6 | FPC11 |
| 12 | 15.3 | 15.2 | 471.2752 | 471.2751 | −0.2 | 471 (100), 472 (27), 473 (4), 207 (3) | C28H40O6 | FPC12 Macrocarpal A |
| 13 | 15.6 | 6.9 | 471.2752 | 471.2751 | −0.2 | 471 (100), 472 (28), 473 (5), 207 (3) | C28H40O6 | FPC13 |
| 14 | 15.9 | 7.7 | 485.2544 | 485.2545 | 0.2 | 485 (100), 486 (29), 207 (8), 250 (2) | C28H38O7 | FPC14 |
| 15 | 16.3 | 16.2 | 471.2752 | 471.2751 | −0.2 | 471 (100), 472 (27), 473 (4), 207 (3) | C28H40O6 | FPC15 |
| 16 | 16.9 | 15.8 | 471.2752 | 471.2752 | 0.0 | 471 (100), 472 (29), 473 (5), 207 (3) | C28H40O6 | FPC16 |
| 17 | 17.1 | 0.5 | 485.2544 | 485.2544 | 0.0 | 485 (100), 486 (30), 207 (12), 250 (2) | C28H38O7 | FPC17 |
| 18 | 17.8 | 33.1 | 471.2752 | 471.2750 | −0.4 | 471 (100), 472 (27), 473 (4), 207 (2) | C28H40O6 | FPC18 Macrocarpal L |
| 19 | 18.2 | 24.2 | 471.2752 | 471.2751 | −0.2 | 471 (100), 472 (27), 473 (4), 207 (4) | C28H40O6 | FPC19 |
| 20 | 19.0 | 49.2 | 471.2752 | 471.2750 | −0.4 | 471 (100), 207 (48), 250 (28), 472 (26) | C28H40O6 | FPC20 Macrocarpal D |
| 21 | 22.3 | 15.7 | 499.1609 | 499.1608 | −0.2 | 249 (100), 250 (15), 181 (1), 251 (1) | C26H28O10 | FPC21 |
| 22 | 23.3 | 18.3 | 499.1609 | 499.1610 | 0.2 | 249 (100), 250 (13), 181 (1), 251 (2) | C26H28O10 | FPC22 |
| 23 | 23.8 | 100.0 | 499.1609 | 499.1607 | −0.4 | 249 (100), 250 (12), 181 (3), 251 (1) | C26H28O10 | FPC23 Sideroxylonal A |
| Concentration (mg g−1 DM) | Macrocarpal A | Macrocarpal D | Macrocarpal J | Macrocarpal L | Macrocarpal N | Sideroxylonal A | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | |
| Wounding | 0.04 | 0.84 | 0.96 | 0.37 | N.A.1 | N.A.1 | 62.43 | 0.00 | 0.51 | 0.50 | 35.32 | 0.00 |
| Time | 0.27 | 0.62 | 0.02 | 0.88 | N.A.1 | N.A.1 | 0.09 | 0.78 | 1.11 | 0.33 | 0.00 | 0.95 |
| Time × Time | 0.30 | 0.60 | 0.02 | 0.89 | N.A.1 | N.A.1 | 0.07 | 0.81 | 3.91 | 0.10 | 0.28 | 0.62 |
| Wounding × Time | 0.00 | 0.97 | 5.49 | 0.06 | N.A.1 | N.A.1 | 2006.52 | 0.00 | 10.34 | 0.02 | 12.71 | 0.01 |
| Ozone | 48.01 | 0.00 | 88.77 | 0.00 | 1.84 | 0.22 | 721.19 | 0.00 | 41.56 | 0.00 | 46.06 | 0.00 |
| Time | 10.78 | 0.02 | 2.48 | 0.17 | 0.56 | 0.48 | 2.68 | 0.16 | 2.39 | 0.17 | 0.08 | 0.78 |
| Time × Time | 5.27 | 0.06 | 1.52 | 0.26 | 0.39 | 0.55 | 1.75 | 0.23 | 4.19 | 0.09 | 0.40 | 0.55 |
| Ozone × Time | 18.30 | 0.01 | 2.96 | 0.14 | 13.97 | 0.01 | 21.69 | 0.00 | 41.69 | 0.00 | 17.51 | 0.01 |
| Ozone + Wounding | 6.22 | 0.05 | 1.07 | 0.34 | 4.41 | 0.08 | 0.81 | 0.40 | 23.26 | 0.00 | 140.11 | 0.00 |
| Time | 0.06 | 0.82 | 0.11 | 0.75 | 0.11 | 0.75 | 0.18 | 0.69 | 0.28 | 0.62 | 0.03 | 0.87 |
| Time × Time | 0.13 | 0.73 | 0.23 | 0.65 | 0.20 | 0.67 | 0.36 | 0.57 | 0.99 | 0.36 | 0.00 | 0.99 |
| (Ozone + Wounding) × Time | 20.38 | 0.00 | 1.01 | 0.35 | 3.36 | 0.12 | 0.00 | 0.98 | 5.76 | 0.05 | 14.72 | 0.01 |
| Concentration (mg g−1 DM) | Total Macrocarpals | Total Sideroxylonals | Total FPCs | Total Phenolics | ||||
|---|---|---|---|---|---|---|---|---|
| F | Sig. | F | Sig. | F | Sig. | F | Sig. | |
| Wounding | 4.42 | 0.08 | 27.90 | 0.00 | 15.70 | 0.01 | 115.88 | 0.00 |
| Time | 0.94 | 0.37 | 0.42 | 0.54 | 0.67 | 0.44 | 0.40 | 0.56 |
| Time × Time | 1.04 | 0.35 | 0.65 | 0.45 | 0.87 | 0.39 | 2.17 | 0.19 |
| Wounding × Time | 0.00 | 0.98 | 13.71 | 0.01 | 1.72 | 0.24 | 27.52 | 0.00 |
| Ozone | 20.56 | 0.00 | 5.33 | 0.06 | 64.02 | 0.00 | 12.30 | 0.01 |
| Time | 1.51 | 0.27 | 0.01 | 0.94 | 2.80 | 0.15 | 4.31 | 0.08 |
| Time × Time | 4.75 | 0.07 | 0.26 | 0.63 | 5.33 | 0.06 | 6.55 | 0.04 |
| Ozone × Time | 0.09 | 0.77 | 2.56 | 0.16 | 0.54 | 0.49 | 10.47 | 0.02 |
| Ozone + Wounding | 0.01 | 0.94 | 16.44 | 0.01 | 1.45 | 0.27 | 924.63 | 0.00 |
| Time | 1.99 | 0.21 | 0.04 | 0.85 | 1.29 | 0.30 | 0.07 | 0.79 |
| Time × Time | 4.65 | 0.07 | 0.29 | 0.61 | 2.78 | 0.15 | 0.39 | 0.55 |
| (Ozone + Wounding) × Time | 0.28 | 0.62 | 2.79 | 0.15 | 1.50 | 0.27 | 176.34 | 0.00 |
| Composition (%) | Macrocarpal A | Macrocarpal D | Macrocarpal J | Macrocarpal L | Macrocarpal N | Sideroxylonal A | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | F | Sig. | |
| Wounding | 13.53 | 0.01 | 29.27 | 0.00 | N.A.1 | N.A.1 | 16.96 | 0.01 | 4.81 | 0.07 | 302.54 | 0.00 |
| Time | 7.13 | 0.04 | 5.43 | 0.06 | N.A.1 | N.A.1 | 4.32 | 0.08 | 0.97 | 0.36 | 0.07 | 0.81 |
| Time × Time | 4.50 | 0.08 | 6.04 | 0.05 | N.A.1 | N.A.1 | 4.73 | 0.07 | 0.04 | 0.85 | 0.20 | 0.67 |
| Wounding × Time | 1.72 | 0.24 | 15.94 | 0.01 | N.A.1 | N.A.1 | 5.33 | 0.06 | 29.86 | 0.00 | 131.52 | 0.00 |
| Ozone | 12.74 | 0.01 | 1.18 | 0.32 | 106.17 | 0.00 | 0.51 | 0.50 | 0.23 | 0.65 | 39.21 | 0.00 |
| Time | 1.19 | 0.32 | 4.19 | 0.09 | 2.74 | 0.15 | 2.04 | 0.20 | 3.22 | 0.13 | 0.39 | 0.55 |
| Time × Time | 0.40 | 0.55 | 2.10 | 0.20 | 1.54 | 0.26 | 1.61 | 0.25 | 4.45 | 0.08 | 2.83 | 0.14 |
| Ozone × Time | 7.78 | 0.03 | 12.09 | 0.01 | 0.13 | 0.73 | 4.83 | 0.07 | 7.98 | 0.03 | 13.54 | 0.01 |
| Ozone + Wounding | 22.29 | 0.00 | 0.89 | 0.38 | 47.33 | 0.00 | 4.23 | 0.09 | 7.10 | 0.04 | 16.99 | 0.01 |
| Time | 2.10 | 0.20 | 0.10 | 0.77 | 0.17 | 0.69 | 0.01 | 0.91 | 35.06 | 0.01 | 0.00 | 0.95 |
| Time ×* Time | 1.14 | 0.33 | 0.25 | 0.64 | 0.02 | 0.90 | 0.05 | 0.83 | 67.31 | 0.00 | 0.29 | 0.61 |
| (Ozone + Wounding) × Time | 88.99 | 0.00 | 0.48 | 0.52 | 2.27 | 0.18 | 4.11 | 0.09 | 2.21 | 0.19 | 160.49 | 0.00 |
| % of Total FPCs | Macrocarpal A | Macrocarpal D | Macrocarpal J | Macrocarpal L | Macrocarpal N | Sideroxylonal A | |
|---|---|---|---|---|---|---|---|
| 0.5 | C | 1.42 ± 0.82 | 7.61 ± 4.39 | 0.06 ± 0.03 | 9.03 ± 5.21 | 0.36 ± 0.21 | 29.31 ± 16.92 |
| W | 1.02 ± 0.59 | 4.05 ± 2.34 | 0.00 ± 0.00 | 4.92 ± 2.84 | 0.39 ± 0.22 | 23.82 ± 13.75 | |
| O | 2.13 ± 1.23 | 6.52 ± 3.76 | 0.00 ± 0.00 | 7.97 ± 4.60 | 0.17 ± 0.10 | 15.88 ± 9.17 | |
| O + W | 0.85 ± 0.49 | 4.58 ± 2.64 | 0.02 ± 0.01 | 6.31 ± 3.64 | 0.14 ± 0.08 | 20.07 ± 11.58 | |
| 25 | C | 1.27 ± 0.73 | 5.54 ± 3.20 | 0.07 ± 0.04 | 6.43 ± 3.71 | 0.29 ± 0.17 | 22.80 ± 13.16 |
| W | 0.90 ± 0.52 | 3.59 ± 2.07 | 0.00 ± 0.00 | 4.63 ± 2.67 | 0.36 ± 0.21 | 30.04 ± 17.34 | |
| O | 2.68 ± 1.55 | 7.25 ± 4.18 | 0.00 ± 0.00 | 8.26 ± 4.77 | 0.38 ± 0.22 | 16.44 ± 9.49 | |
| O + W | 2.37 ± 1.37 | 7.62 ± 4.40 | 0.00 ± 0.00 | 9.49 ± 5.48 | 0.45 ± 0.26 | 20.23 ± 11.68 | |
| 50 | C | 1.10 ± 0.63 | 4.70 ± 2.71 | 0.08 ± 0.04 | 6.68 ± 3.85 | 0.11 ± 0.06 | 27.45 ± 15.84 |
| W | 1.10 ± 0.64 | 4.67 ± 2.70 | 0.00 ± 0.00 | 5.54 ± 3.20 | 0.37 ± 0.21 | 18.52 ± 10.69 | |
| O | 1.67 ± 0.96 | 5.89 ± 3.40 | 0.00 ± 0.00 | 8.17 ± 4.71 | 0.09 ± 0.05 | 19.35 ± 11.17 | |
| O + W | 1.84 ± 1.06 | 7.07 ± 4.08 | 0.01 ± 0.01 | 8.27 ± 4.77 | 0.38 ± 0.22 | 22.63 ± 13.06 | |
| 75 | C | 1.49 ± 0.86 | 6.09 ± 3.51 | 0.09 ± 0.05 | 8.43 ± 4.87 | 0.18 ± 0.10 | 21.49 ± 12.41 |
| W | 1.19 ± 0.69 | 5.61 ± 3.24 | 0.00 ± 0.00 | 6.14 ± 3.54 | 0.42 ± 0.24 | 18.88 ± 10.90 | |
| O | 1.72 ± 0.99 | 7.34 ± 4.24 | 0.00 ± 0.00 | 8.60 ± 4.96 | 0.28 ± 0.16 | 18.31 ± 10.57 | |
| O + W | 0.92 ± 0.53 | 4.87 ± 2.81 | 0.01 ± 0.01 | 5.87 ± 3.39 | 0.13 ± 0.07 | 20.70 ± 11.95 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Marques dos Santos, B.; Kanagendran, A.; Neilson, E.H.J.; Niinemets, Ü. Ozone and Wounding Stresses Differently Alter the Temporal Variation in Formylated Phloroglucinols in Eucalyptus globulus Leaves. Metabolites 2019, 9, 46. https://doi.org/10.3390/metabo9030046
Liu B, Marques dos Santos B, Kanagendran A, Neilson EHJ, Niinemets Ü. Ozone and Wounding Stresses Differently Alter the Temporal Variation in Formylated Phloroglucinols in Eucalyptus globulus Leaves. Metabolites. 2019; 9(3):46. https://doi.org/10.3390/metabo9030046
Chicago/Turabian StyleLiu, Bin, Bruna Marques dos Santos, Arooran Kanagendran, Elizabeth H. Jakobsen Neilson, and Ülo Niinemets. 2019. "Ozone and Wounding Stresses Differently Alter the Temporal Variation in Formylated Phloroglucinols in Eucalyptus globulus Leaves" Metabolites 9, no. 3: 46. https://doi.org/10.3390/metabo9030046
APA StyleLiu, B., Marques dos Santos, B., Kanagendran, A., Neilson, E. H. J., & Niinemets, Ü. (2019). Ozone and Wounding Stresses Differently Alter the Temporal Variation in Formylated Phloroglucinols in Eucalyptus globulus Leaves. Metabolites, 9(3), 46. https://doi.org/10.3390/metabo9030046