Flavan-3-ols Content in Red Raspberry Leaves Increases under Blue Led-Light Irradiation

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Raspberry Cultivation in a Plant Factory Unit and LED Light Irradiation
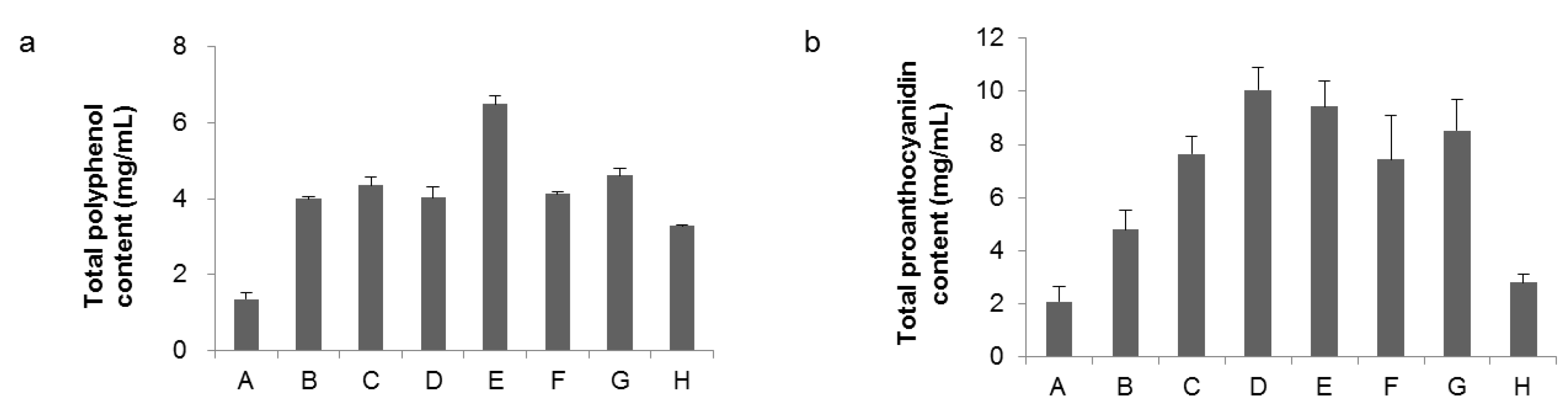
2.2. Analysis of Total Polyphenols and Total Proanthocyanidin Amounts in Extracts from Raspberry Leaves
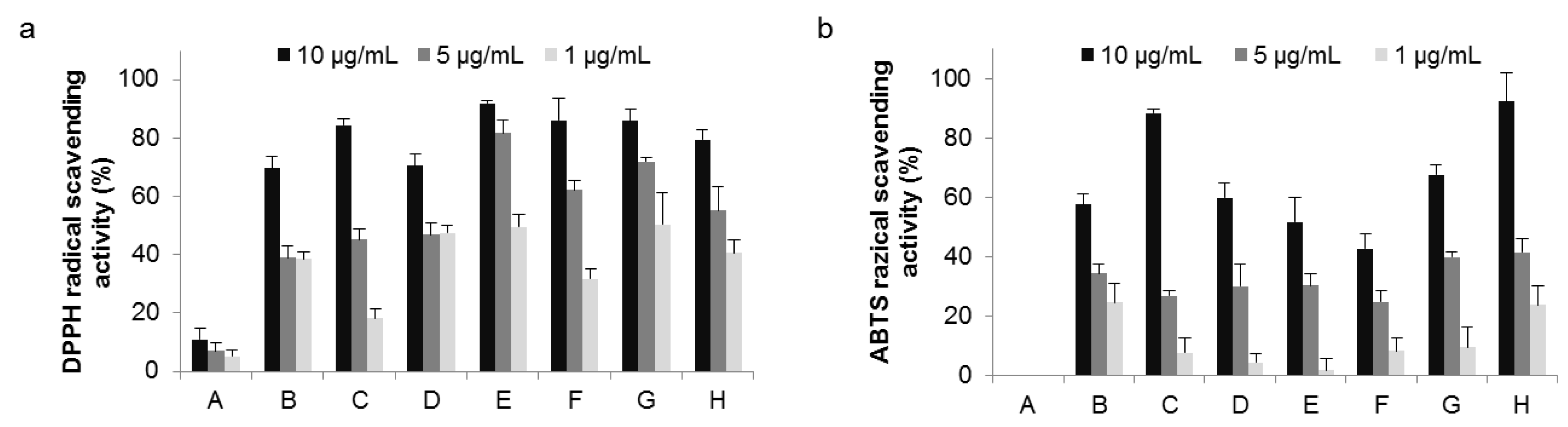
2.3. Radical Scavenging Activity
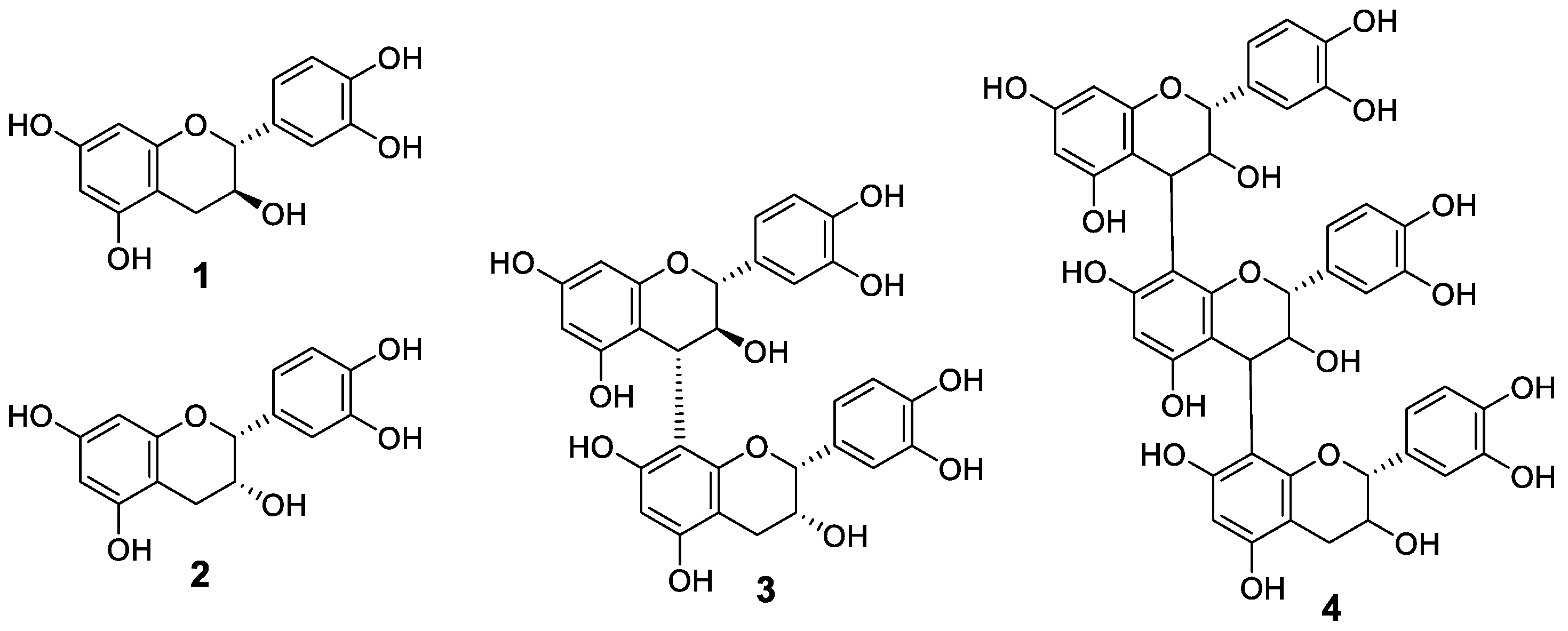
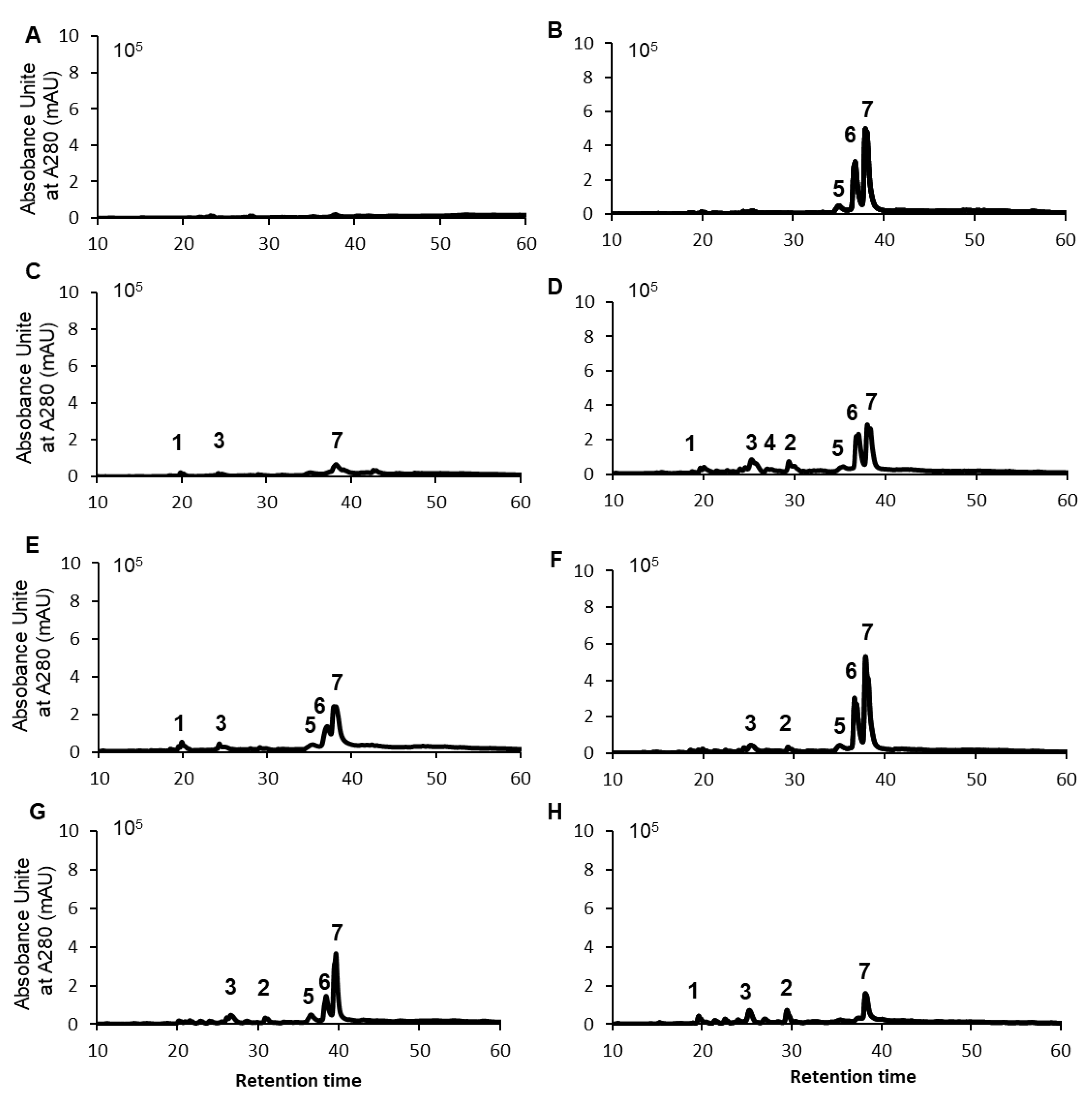
2.4. HPLC Analysis and Identification of Polyphenols Contained in Raspberry Leaf Extracts
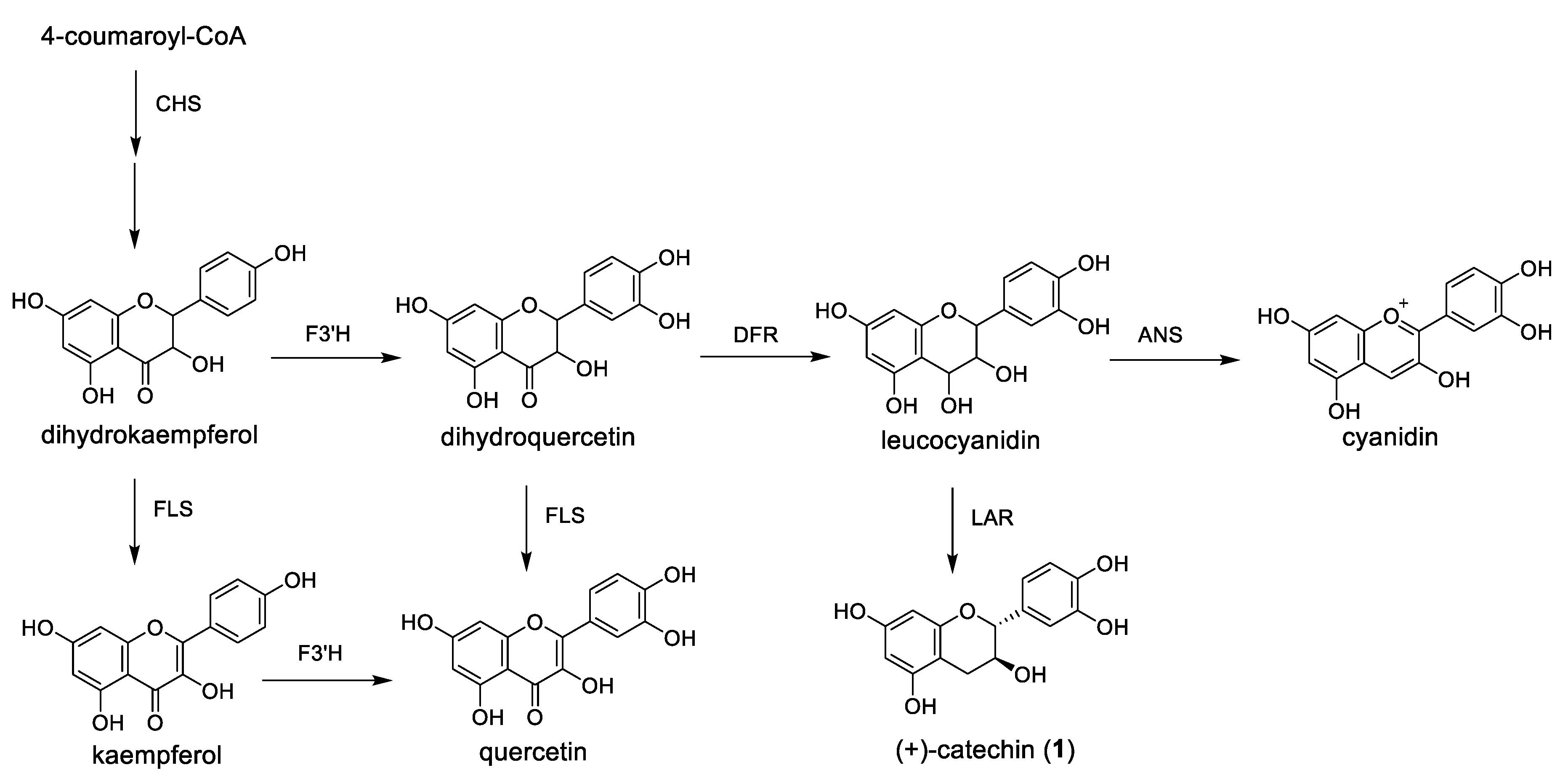
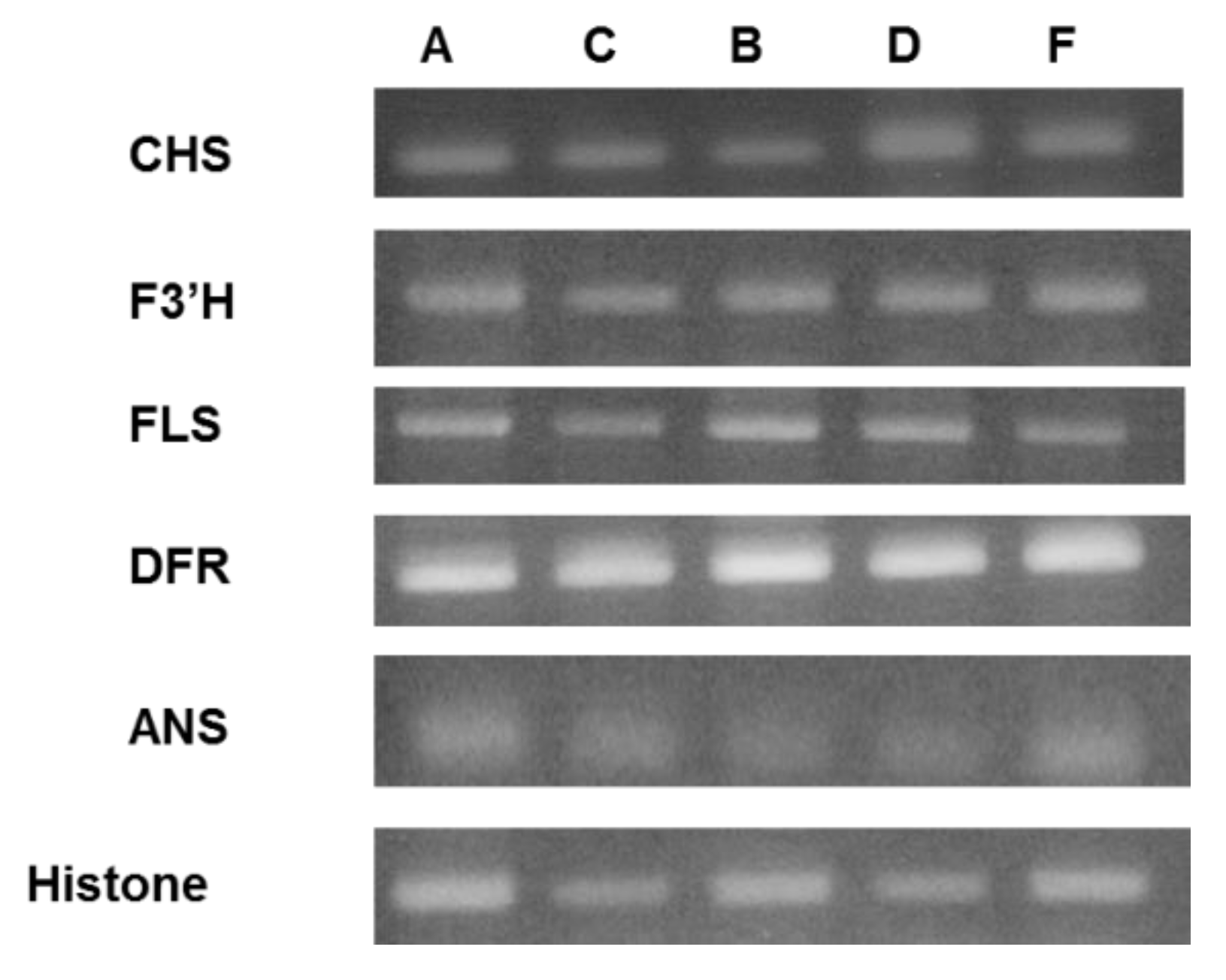
2.5. Semi-Quantitative RT-PCR Analysis of Expression Level of the Flavan-3-ol Derivative Biosynthetic Enzyme
3. Materials and Methods
3.1. Reagents and Materials
3.2. Cultivation of Raspberry
3.3. Preparation of Crude Extract Solution of Raspberry Leaves
3.4. Total Polyphenol Analysis
3.5. Total Proanthocyanidin Analysis
3.6. DPPH Radical Scavenging Assay
3.7. ABTS Radical Scavenging Assay
3.8. LC-MS Analysis Conditions
3.9. Semi-Quantitative RT-PCR Analysis of Flavan-3-ol Derivative Biosynthetic-Enzyme-Related Gene Expression
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harborne, J.B. The Flavonoids: Advances in Research from 1986; Chapman and Hall: London, UK, 1993. [Google Scholar]
- Harborne, J.B.; Baxter, H. The Handbook of Natural Flavonoids; Wiley: New York, NY, USA, 1999. [Google Scholar]
- Zhao, J.; Pang, Y.; Dixon, R.A. The mysteries of proanthocyanidin transport and polymerization. Plant Physiol. 2010, 153, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, Z.; Maximova, S.; Payne, M.J.; Guiltinan, M.J. Proanthocyanidin synthesis in Theobroma cacao: Genes encoding anthocyanidin synthase, anthocyanidin reductase, and leucoanthocyanidin reductase. BMC Plant Biol. 2013, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.F.; Ji, W.; Gao, M.Y.; Niu, T.Q.; Xing, Y.F.; Niu, X.Y. Accumulation of flavanols and expression of leucoanthocyanidin reductase induced by postharvest UV-C irradiation in grape berry. Genet. Mol. Res. 2015, 14, 7687–7695. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, Y.B.; Li, X.; Choi, S.R.; Park, S.; Park, J.S.; Lim, Y.P.; Park, S.U. Accumulation of polyphenolpropanoids by white, blue, and red light irradiation and their organ-specific distribution in Chinese cabbage (Brassica rapa ssp. pekinensis). J. Agric. Food Chem. 2015, 63, 6772–6778. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Nakajima, N.; Tanaka, A.; Ubukata, M. Synthetic studies of proanthocyanidins. Part 2. Stereoselective gram-scale synthesis of procyanidin-B3. Tetrahedron 2002, 58, 7829–7837. [Google Scholar] [CrossRef]
- Saito, A.; Nakajima, N.; Tanaka, A.; Ubukata, M. Synthetic studies of proanthocyanidins. Part 4. The synthesis of procyanidin B1 and B4. TMSOTf-catalyzed cyclization of catechin and epicatechin condensation. Heterocycles 2003, 61, 287–298. [Google Scholar]
- Saito, A.; Doi, Y.; Tanaka, A.; Matsuura, N.; Ubukata, M.; Nakajima, N. Systematic synthesis of four epicatechin series procyanidin trimers and their inhibitory activity on the Maillard reaction and antioxidant activity. Bioorg. Med. Chem. 2004, 12, 4783–4790. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Mizushina, Y.; Tanaka, A.; Nakajima, N. Versatile synthesis of epicatechin series procyanidin oligomers, and their antioxidant and DNA polymerase inhibitory activity. Tetrahedron 2009, 65, 7422–7428. [Google Scholar] [CrossRef]
- Hamada, Y.; Takano, S.; Ayano, Y.; Tokunaga, M.; Koashi, T.; Okamoto, S.; Doi, S.; Ishida, M.; Kawasaki, T.; Hamada, M.; et al. Structure–activity relationship of oligomeric flavan-3-ols: Importance of upper-unit B-ring hydroxyl groups in the dimeric structure for strong activities. Molecules 2015, 20, 18870–18885. [Google Scholar] [CrossRef]
- Higashino, Y.; Okamoto, T.; Mori, K.; Kawasaki, T.; Hamada, M.; Nakajima, N.; Saito, A. Regioselective synthesis of procyanidin B6, a 4-6-condensed (+)-catechin dimer, by intramolecular condensation. Molecules 2018, 23, 205. [Google Scholar] [CrossRef]
- Saito, A.; Emoto, M.; Tanaka, A.; Doi, Y.; Shoji, K.; Mizushina, Y.; Ikawa, H.; Yoshida, H.; Matsuura, N.; Nakajima, N. Stereoselective synthesis of procyanidin B3-3-O-gallate and 3,3”-di-O-gallate, and their abilities as antioxidant and DNA polymerase inhibitor. Tetrahedron 2004, 60, 12043–12049. [Google Scholar] [CrossRef]
- Saito, A.; Mizushina, Y.; Ikawa, H.; Yoshida, H.; Doi, Y.; Tanaka, A.; Nakajima, N. Systematic synthesis of galloyl-substituted procyanidin B1 and B2, and their ability of DPPH radical scavenging activity and inhibitory activity of DNA polymerases. Bioorg. Med. Chem. 2005, 13, 2759–2771. [Google Scholar] [CrossRef]
- Sakuda, H.; Saito, A.; Mizushina, Y.; Yoshida, H.; Tanaka, A.; Nakajima, N. Synthesis of galloyl-substituted procyanidin B4 series, and their deep radical scavenging activity and DNApolymerase inhibitory activity. Heterocycles 2006, 67, 175–188. [Google Scholar]
- Mori, K.; Ayano, Y.; Hamada, Y.; Hojima, T.; Tanaka, R.; Higashino, Y.; Izuno, M.; Okamoto, T.; Kawasaki, T.; Hamada, M.; et al. Role of 2,3-cis structure of (–)-epicatechin-3,5-O-digallate in inhibition of HeLa S3 cell proliferation. Nat. Prod. Chem. Res. 2015, 3, 172. [Google Scholar]
- Hojima, T.; Komeda, S.; Higashino, Y.; Hamada, M.; Nakajima, N.; Kawasaki, T.; Saito, A. Role of 3,5-digalloyl and 3′,4′-dihydroxyl structure of (–)-epicatechin-3,5-digallate in inhibition of HeLa S3 cell proliferation. Nat. Prod. Chem. Res. 2017, 5, 250. [Google Scholar] [CrossRef]
- Matsubara, K.; Saito, A.; Tanaka, A.; Nakajima, N.; Akagi, R.; Mori, M.; Mizushina, Y. Epicatechin conjugated with fatty acid is a potent inhibitor of DNA polymerase and angiogenesis. Life Sci. 2007, 80, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, S.; Doi, S.; Harui, K.; Okamoto, T.; Okamoto, S.; Uenishi, J.; Kawasaki, T.; Nakajima, N.; Saito, A. Development of a new synthetic strategy for procyanidin dimer condensation using peracetylated electrophiles. Heterocycles 2014, 88, 1595–1602. [Google Scholar]
- Okamoto, S.; Ishihara, S.; Okamoto, T.; Doi, S.; Harui, K.; Higashino, Y.; Kawasaki, T.; Nakajima, N.; Saito, A. Inhibitory activity of synthesized acetylated procyanidin B1 analogues against HeLa S3 cells proliferation. Molecules 2014, 19, 1775–1785. [Google Scholar] [CrossRef]
- Tsukuda, S.; Watashi, K.; Hojima, T.; Isogawa, M.; Iwamoto, M.; Omagari, K.; Suzuki, R.; Aizaki, H.; Kojima, S.; Sugiyama, M.; et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology 2017, 65, 1104–1116. [Google Scholar] [CrossRef]
- Hummer, K.E. Rubus pharmacology: Antiquity to the present. HortScience 2010, 45, 1587–1591. [Google Scholar] [CrossRef]
- Rao, A.V.; Synder, D.M. Rasberries and human health: A review. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef] [PubMed]
- Szymanowska, U.; Baraniak, B.; Bogucka-Kocka, A. Antioxidant, anti-inflammatory, and postulated cytotoxic activity of phenolic and anthocyanidin-rich fractions from polana raspberry (Rubus ideaus L.) fruit and juice-in vitro study. Molecules 2018, 23, 1812. [Google Scholar] [CrossRef] [PubMed]
- Olle, M.; Ngouajio, M.; Siomos, A. Vegetable quality and productivity as influenced by growing medium: A review. Žemdirb. Agric. 2012, 99, 399–408. [Google Scholar]
- Beekwilder, J.; Jonker, H.; Meesters, P.; Hall, R.D.; Van der Meer, I.M.; Ric de Vos, C.H. Antioxidants in Raspberry: On line analysis links antioxidant ancivity to diversity of individual metabolites. J. Agric. Food Chem. 2005, 53, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M.; Glod, D.; Kula, M.; Majdam, M.; Halasa, R.; Matkowski, A.; Kozlowska, W.; Kawiak, A. Chemical composition and biological activity of Rubus ideaus shoots—A traditional herbal remedy of Eastern Europe. BMC Compl. Altern. Med. 2014, 14, 480. [Google Scholar] [CrossRef] [PubMed]
- Kassaim, A.; Poette, J.; Paterson, A.; Zait, D.; McCallum, S.; Woodhead, M.; Snith, K.; Hackett, C.; Graham, J. Environmental and seasonal influences on red raspberry anthocyanin antioxidant contents and identification of quantitive traits loci (QTL). Mol. Nutr. Food Res. 2009, 53, 625–634. [Google Scholar] [CrossRef]
- Xu, Y.; Charkes, M.T.; Luo, Z.; Mimee, B.; Veronneau, P.Y.; Rolland, D.; Roussel, D. Preharvest Ultraviolet C irradiation increased the level of polyphenol accumulation and flavonoid pathway gene expression in strawberry fruit. J. Agric. Food Chem. 2017, 65, 9970–9979. [Google Scholar] [CrossRef]
- Nadeem, M.; Abbasi, B.H.; Younas, M.; Ahmad, W.; Zahir, A.; Hano, C. LED-enhanced biosynthesis of biologically active ingredients in callus cultures of Ocimum basilicum. J. Photochem. Photobiol. B. 2019, 190, 172–178. [Google Scholar] [CrossRef]
- Saito, A. Challenges and complexity of functionality of flavan-3-ol derivatives. Biosci. Biotechnol. Biochem. 2017, 81, 1055–1060. [Google Scholar] [CrossRef]
- McCallum, S.; Woodhead, M.; Hackett, C.A.; Kassim, A.; Paterson, A.; Graham, J. Genetic and environmental effects influencing fruit color and QTL analysis in raspberry. Theor. Appl. Genet. 2010, 121, 611–627. [Google Scholar] [CrossRef]
- Folin, O.; Denis, W. A Colorimetric method for the determination of phenols (and derivatives) in urine. J. Biol. Chem. 1915, 22, 305–308. [Google Scholar]
- Juilcunen-Tiittoo, R. Phenolic constituents in the leaves of northern willows: Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Harbertrson, J.F.; Kennedy, J.A.; Adams, D.O. Tannin in skins and seed of cabernet sauvignon, syrah, and pinot noir berries during ripening. Am. J. Enol. Vitic. 2002, 52, 54–59. [Google Scholar]
- Cáceres-Mella, Á.; Peña-Neira, Á.; Narváez-Bastias, J.; Jara-Campos, C.; López-Solís, R.; Canals, J.M. Comparison of analysis methods for measuring proanthocyanidins in wines and their relationship with perceived astringcy. Int. J. Food Sci. Technol. 2013, 48, 2588–2594. [Google Scholar] [CrossRef]
- Nanjo, F.; Goto, K.; Seto, R.; Suzuki, M.; Sakai, M.; Hara, Y. Scavenging effects of tea catechins and their derivatives on 1,1-diphenyl-2-picrylhydrazyl radical. Free Radic. Biol. Med. 1996, 21, 895–902. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Zheng, D.; Schröder, G.; Schröder, J.; Hrazdina, G. Molecular and biochemical characterization of three aromatic polyketide synthase genes from Rubus idaeus. Plant Mol. Biol. 2001, 46, 1–15. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| - | A 1 | B 1 | C | D | E | F | G | H |
|---|---|---|---|---|---|---|---|---|
| PFD-B (Blue) (μmol m−2 s−1) | 7.23 | 7.23 | 30.24 | 30.24 | 40.09 | 3.86 | 16.54 | - |
| PFD-R (Red) (μmol m−2 s−1) | 8.33 | 8.33 | - | - | - | 4.43 | 27.87 | - |
| Temperature (°C) | 18 | 22 | 18 | 22 | 22 | 22 | 22 | 22 |
| Extract (g) 2 | 0.37 | 0.30 | 0.32 | 0.28 | 0.25 | 0.30 | 0.33 | 0.30 |
| - | A | B | C | D | E | F | G | H |
|---|---|---|---|---|---|---|---|---|
| (+)-catechin (1) | 0.50 | 0.44 | 0.45 | 0.48 | 0.60 | 0.46 | 0.45 | 0.52 |
| (–)-epicatechin (2) | 0.10 | 0.09 | 0.06 | 0.10 | 0.11 | 0.09 | 0.11 | 0.11 |
| procyanidin B4 (3) | 0.59 | 0.58 | 0.48 | 1.25 | 1.00 | 0.79 | 0.87 | 1.35 |
| flavan-3-ol trimer (4) | 0.34 | 0.32 | 0.30 | 0.38 | 0.38 | 0.34 | 0.37 | 0.39 |
| - | A | B | C | D | E | F | G | H |
|---|---|---|---|---|---|---|---|---|
| (+)-catechin (1) | 1.85 | 1.32 | 1.44 | 1.34 | 1.5 | 1.38 | 1.49 | 1.56 |
| (–)-epicatechin (2) | 0.37 | 0.27 | 0.19 | 0.28 | 0.28 | 0.27 | 0.36 | 0.33 |
| procyanidin B4 (3) | 2.18 | 1.74 | 1.54 | 3.5 | 2.5 | 2.37 | 2.87 | 4.05 |
| flavan-3-ol trimer (4) | 1.26 | 0.96 | 0.96 | 1.06 | 0.95 | 1.02 | 1.22 | 1.17 |
| Gene | Accession No. of Sequence Used for Primer Design | Primer Sequence |
|---|---|---|
| CHS | AF292367 | F: AACCCTTGTTTCTTCGTACCATTA |
| R: GATGGGTAGCTAGTACTTACACAT | ||
| ANS | KX950789.1 | F: ATC GTC ATG CAC ATA GGC GAC ACC |
| R: CCT TGG GCG GCT CAG AGA AAA | ||
| FLS | GT029981 | F: AGG TGA ACA GGT GGA GTT GG |
| R: TGA AGA CCA TCA TCG AAT GC | ||
| DFR | GT029979 | F: ATG CGA AAC AAC TTG CAT TT |
| R: GCT ACG ATT CAC GAC ATT GC | ||
| F3′H | GT029980 | F: TGA TGA AGC TTT ATA AGC ATG TGA GG |
| R: GGG TCC ACT CTC TTG GTG AA | ||
| Histone | AF304365.1 | F: CAA GGA AGC AAT TGG CTA CCA AGG |
| R: AGT TGG ATA TCC TTG GGC ATA ATA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobori, R.; Hashimoto, S.; Koshimizu, H.; Yakami, S.; Hirai, M.; Noro, K.; Kawasaki, T.; Saito, A. Flavan-3-ols Content in Red Raspberry Leaves Increases under Blue Led-Light Irradiation. Metabolites 2019, 9, 56. https://doi.org/10.3390/metabo9030056
Kobori R, Hashimoto S, Koshimizu H, Yakami S, Hirai M, Noro K, Kawasaki T, Saito A. Flavan-3-ols Content in Red Raspberry Leaves Increases under Blue Led-Light Irradiation. Metabolites. 2019; 9(3):56. https://doi.org/10.3390/metabo9030056
Chicago/Turabian StyleKobori, Ryo, Seiya Hashimoto, Hayato Koshimizu, Shuich Yakami, Mizuki Hirai, Kenta Noro, Takashi Kawasaki, and Akiko Saito. 2019. "Flavan-3-ols Content in Red Raspberry Leaves Increases under Blue Led-Light Irradiation" Metabolites 9, no. 3: 56. https://doi.org/10.3390/metabo9030056
APA StyleKobori, R., Hashimoto, S., Koshimizu, H., Yakami, S., Hirai, M., Noro, K., Kawasaki, T., & Saito, A. (2019). Flavan-3-ols Content in Red Raspberry Leaves Increases under Blue Led-Light Irradiation. Metabolites, 9(3), 56. https://doi.org/10.3390/metabo9030056