The Biosynthetic Pathway of Major Avenanthramides in Oat


Abstract
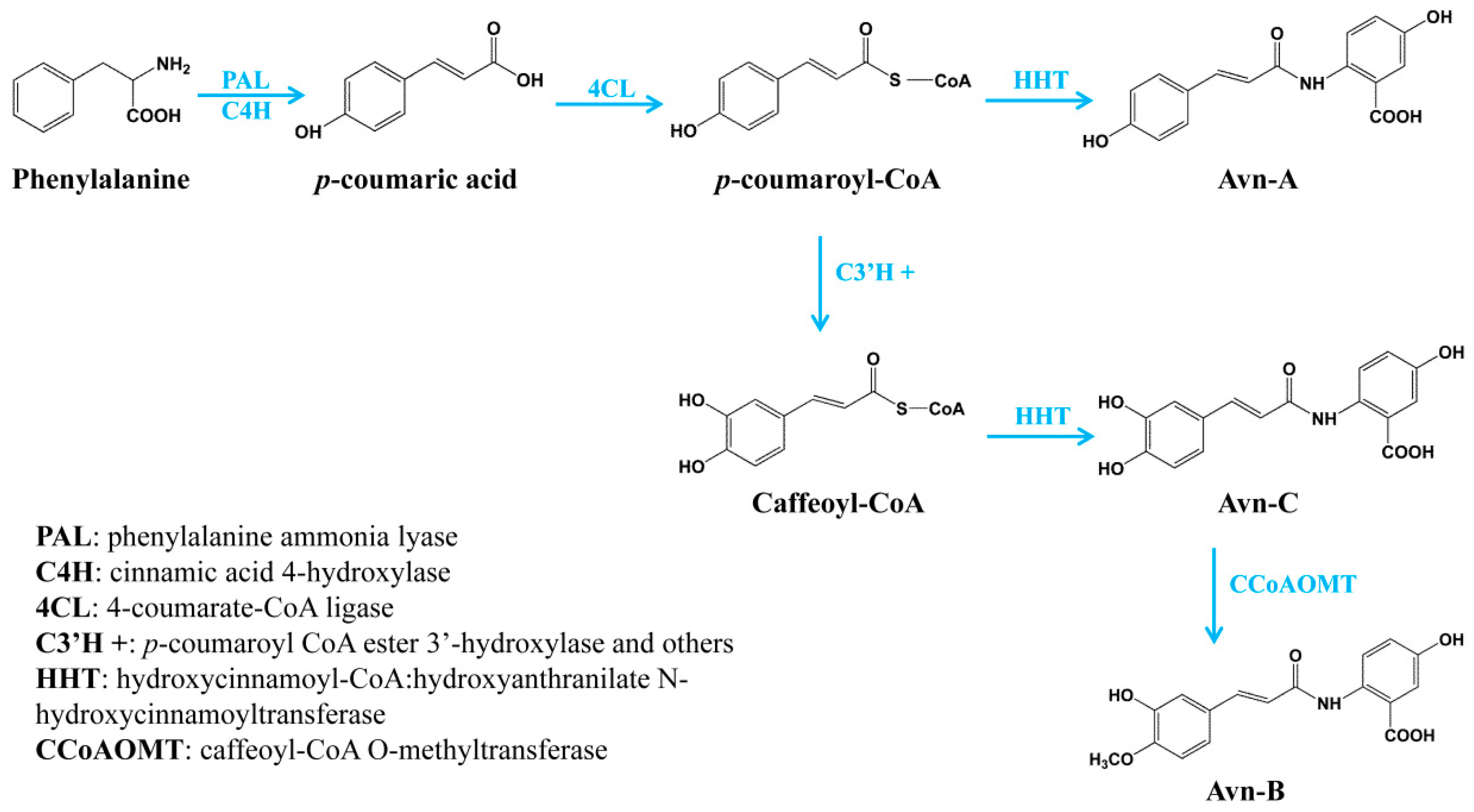
:1. Introduction
2. Results
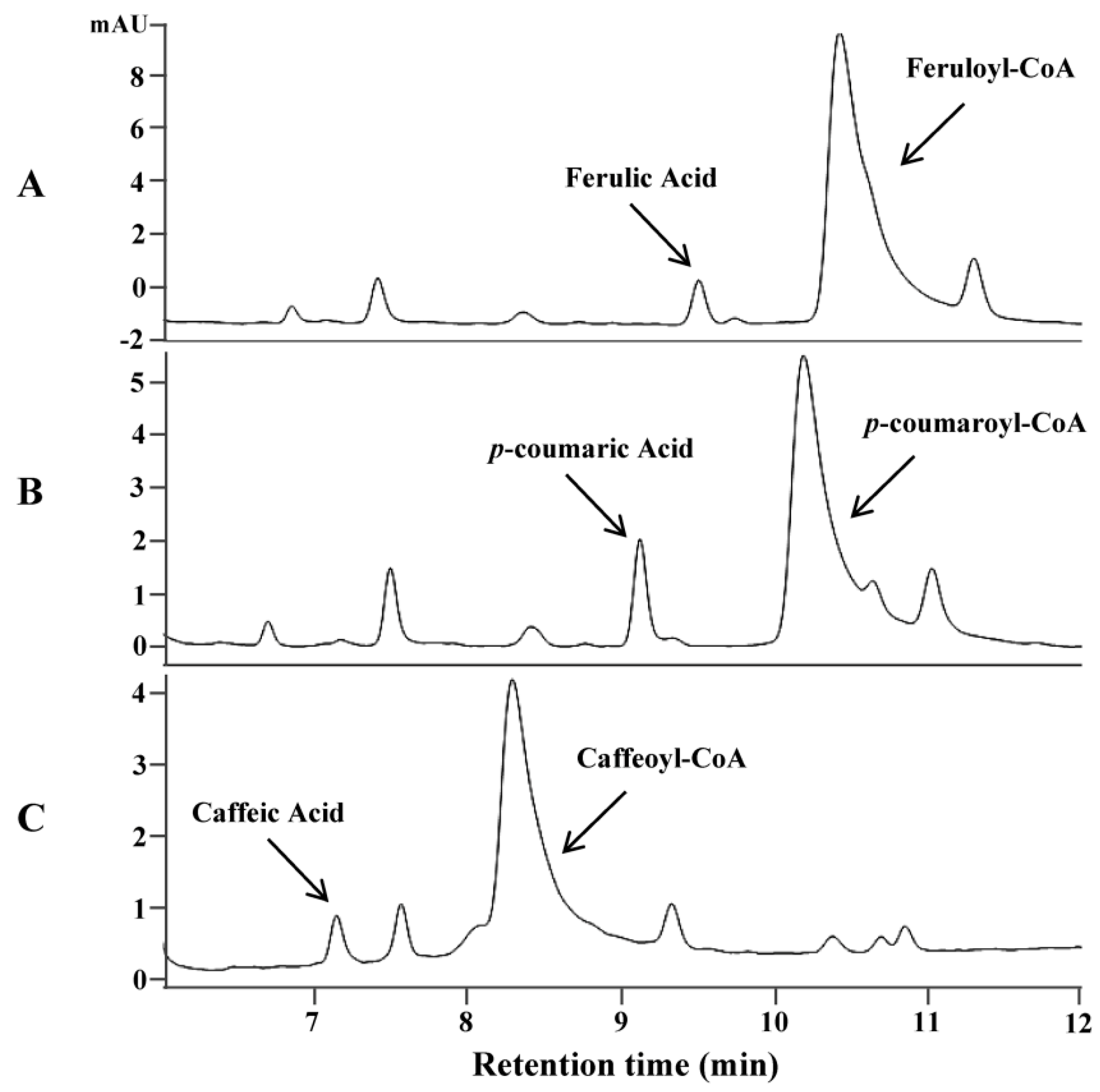
2.1. Identification and Functional Analysis of Genes Encoding 4CLs in the Biosynthesis of Hydroxycinnamate Thioesters
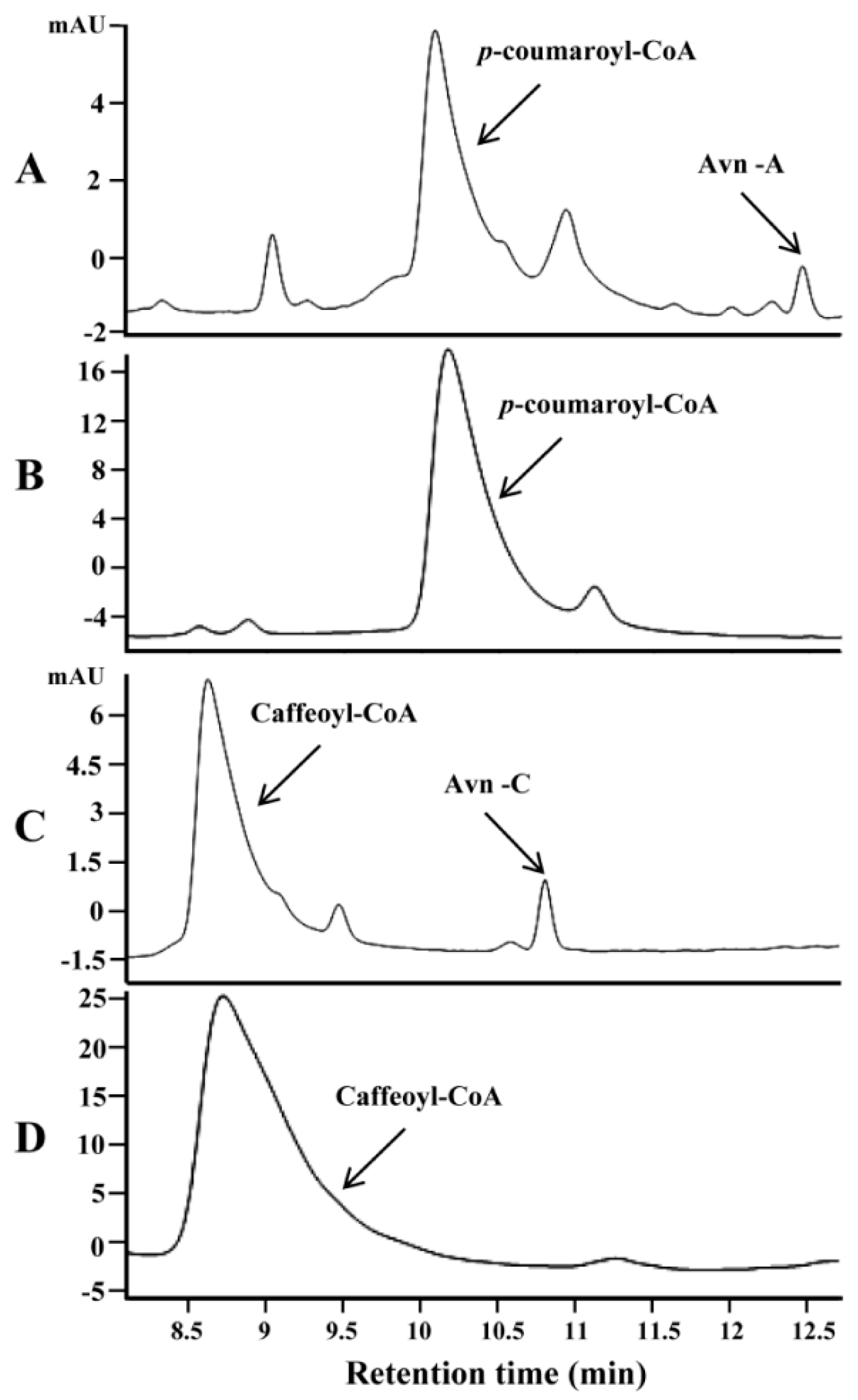
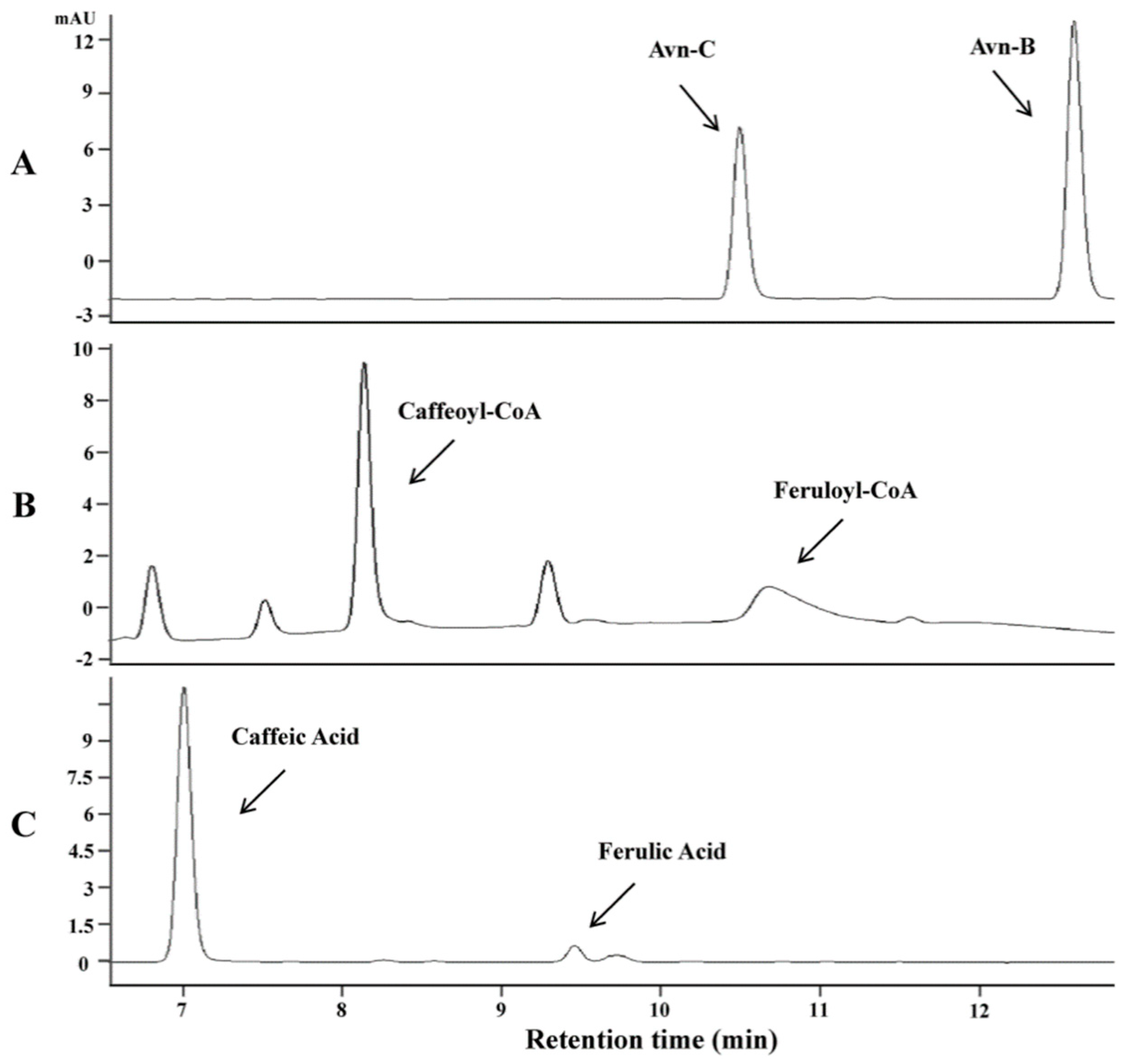
2.2. Identification and Functional Analysis of New Genes Encoding HHT in the Biosynthesis of Avn-A and Avn-C
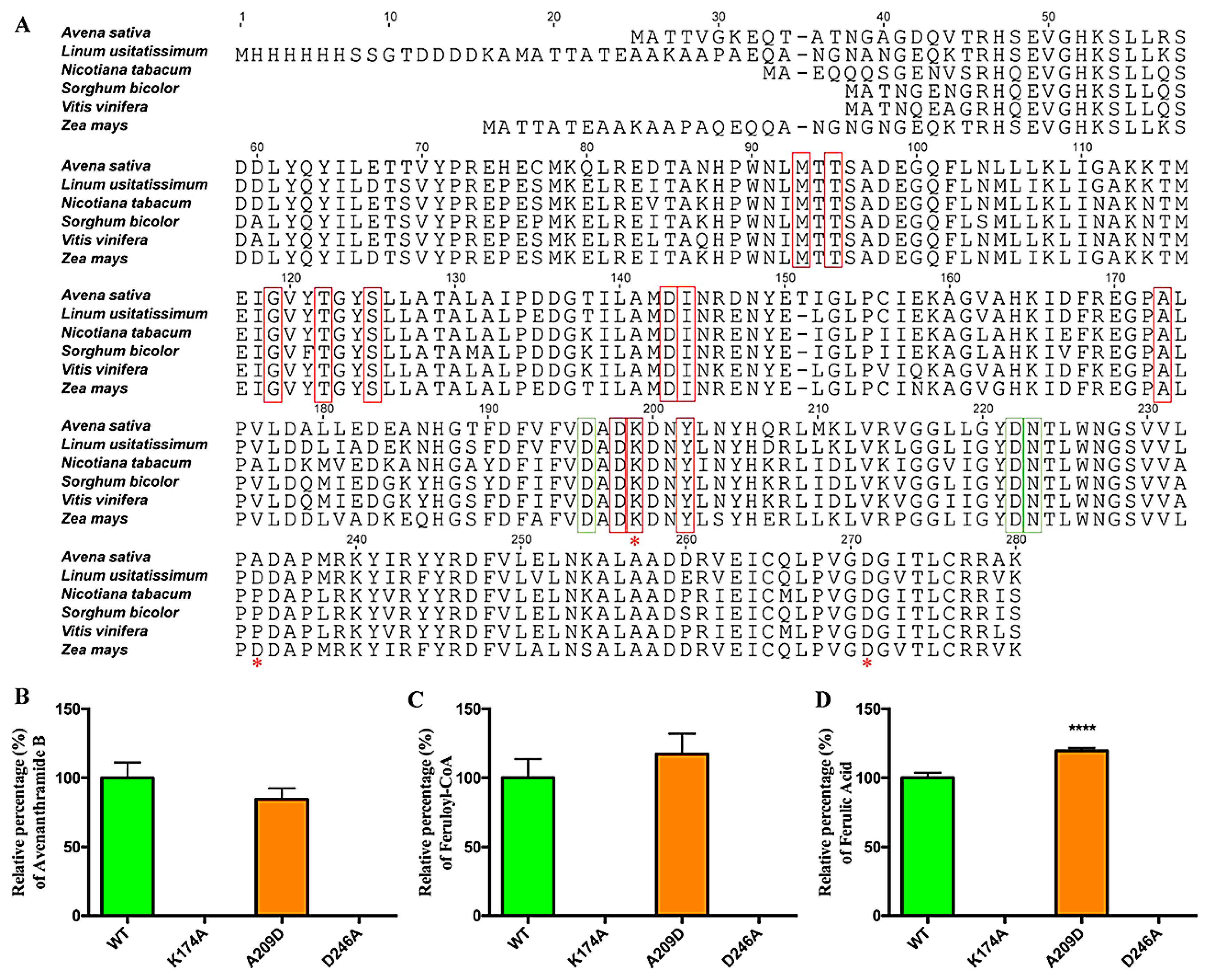
2.3. Identification and Functional Analysis of a Gene Encoding a CCoAOMT Enzyme in the Biosynthesis of Avn-B
3. Discussions
4. Materials and Methods
4.1. Plant Materials
4.2. HPLC Analysis
4.3. RNA Extraction, cDNA Synthesis and Genes Cloning
4.4. Protein Expression and Purification
4.5. Enzyme Assays
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Webster, F.H.; Wood, P.J. Oats: Chemistry and Technology; American Association of Cereal Chemists: St. Paul, MN, USA, 2011. [Google Scholar]
- Sadiq-Butt, M.; Tahir-Nadeem, M.; Khan, M.K.I.; Shabir, R.; Butt, M.S. Oat: Unique among the cereals. Eur. J. Nutr. 2008, 47, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; De, S.; Belkheir, A. Avena sativa (oat), a potential neutraceutical and therapeutic agent: An overview. Crit. Rev. Food Sci. Nutr. 2013, 53, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zubik, L.; Colllins, W.; Marko, M.; Meydani, M. The antiatherogenic potential of oat phenolic compounds. Atherosclerosis 2004, 175, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Sur, R.; Nigam, A.; Grote, D.; Liebel, F.; Southall, M.D. Avenanthramides, polyphenols from oats, exhibit anti-inflammatory and anti-itch activity. Arch. Dermatol. Res. 2008, 300, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M. Potential health benefits of avenanthramides of oats. Nutr. Rev. 2009, 67, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Mayama, S.; Tani, T.; Matsuura, Y. The production of phytoalexins by oat in response to crown rust, Puccinia coronate f. sp. avenae. Physiol. Plant Pathol. 1981, 19, 217–226. [Google Scholar] [CrossRef]
- Collins, F.W. Oat phenolics: Avenanthramides, novel substituted N-cinnamoylanthranilate alkaloids from oat groats and hulls. J. Agric. Food Chem. 1989, 37, 60–66. [Google Scholar] [CrossRef]
- Wise, M.L. Effect of chemical systemic acquired resistance elicitors on avenanthramide biosynthesis in oat (Avena sativa). J. Agric. Food Chem. 2011, 59, 7028–7038. [Google Scholar] [CrossRef]
- Collins, F.W.; Mullin, W.J. High-performance liquid chromatographic determination of avenanthramides, N-aroylanthranilic acid alkaloids from oats. J. Chromatogr. 1988, 445, 363–370. [Google Scholar] [CrossRef]
- Ishihara, A.; Matsukawa, T.; Miyagawa, H.; Ueno, T.; Mayama, S.; Iwamura, H. Induction of hydroxycinnamoyl-CoA: Hydroxyanthranilate N-hydroxycinnamoyl-transferase (HHT) activity in oat leaves by victorin C. Z. Naturforsch 1997, 52, 756–760. [Google Scholar] [CrossRef]
- Yang, Q.; Trinh, H.X.; Imai, S.; Ishihara, A.; Zhang, L.; Nakayashiki, H.; Tosa, Y.; Mayama, S. Analysis of the involvement of hydroxyanthranilate hydroxycinnamoyltransferase and caffeoyl-CoA 3-O-methyltransferase in phytoalexin biosynthesis in oat. Mol. Plant Microb. Interact. 2004, 17, 81–89. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, D.; Hu, J.; Zhou, X.; Ye, X.; Reichel, K.L.; Stewart, N.R.; Syrenne, R.D.; Yang, X.; Gao, P.; et al. Comparative genome analysis of lignin biosynthesis gene families across the plant kingdom. BMC Bioinform. 2009, 10, S3. [Google Scholar] [CrossRef] [PubMed]
- Landmann, C.; Hucherig, S.; Fink, B.; Hoffmann, T.; Dittlein, D.; Coiner, H.A.; Schwab, W. Substrate promiscuity of a rosmarinic acid synthase from lavender (Lavandula angustifolia L.). Planta 2011, 234, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. The evolution of phenylpropanoid metabolism in the green lineage. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Molina, I.; Kosma, D. Role of HXXXD-motif/BAHD acyltransferases in the biosynthesis of extracellular lipids. Plant Cell Rep. 2015, 34, 587–601. [Google Scholar] [CrossRef]
- Gross, G.G.; Zenk, M.G. Isolation and properties of hydroxycinnamate: CoA ligase from lignifying tissue of forsythia. Eur. J. Biochem. 1974, 42, 453–459. [Google Scholar] [CrossRef]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Stockigt, J.; Zenk, M.H. Chemical syntheses and properties of hydroxycinnamoyl-Coenzyme A derivatives. Z. Naturforsch. 1975, 30, 352–358. [Google Scholar] [CrossRef]
- Gutierrez-Gonzalez, J.J.; Tu, Z.J.; Garvin, D.F. Analysis and annotation of the hexaploidy oat seed transcriptome. BMC Genom. 2013, 14, 471. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, H.; Zhu, Y.; McBride, J.; Fu, J.; Sang, S. Oat avenanthramide-C (2c) is biotransformed by mice and the human microbiota into bioactive metabolites. J. Nutr. 2015, 145, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.M.; Sattler, S.A.; Regner, M.; Jones, J.P.; Ralph, J.; Vermerris, W.; Sattler, S.E.; Chulhee, K. The structure and catalytic mechanism of sorghum bicolor caffeoyl-CoA O-methyltransferase. Plant Physiol. 2016, 172, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.L.; Zubieta, C.; Dixon, R.A.; Noel, J.P. Crystal structures of alfalfa caffeoyl coenzyme A 3-O-methyltransferase. Plant Physiol. 2005, 137, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Zubieta, C.; Kota, P.; Ferrer, J.L.; Dixon, R.A.; Noel, J.P. Structural basis for the modulation of lignin monomer methylation by caffeic acid/5-hydrocyferulic acid 3/5-O-methyltransferase. Plant Cell 2002, 14, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Green, A.R.; Lewis, K.M.; Barr, J.T.; Jones, J.P.; Lu, F.; Ralph, J.; Vermerris, W.; Sattler, S.E.; Kang, C. Determination of the structure and catalytic mechanism of sorghum bicolor caffeic acid O-methyltransferase and the structure impact of three brown midrib12 mutations. Plant Physiol. 2014, 165, 1440–1456. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Maury, S.; Bergdoll, M.; Thion, L.; Erard, M.; Legrand, M. Identification of the enzymatic active site of tobacco caffeoyl-coenzyme A O-methyltransferase by site-directed mutagenesis. J. Biol. Chem. 2001, 276, 36831–36838. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.P.; Dixon, R.A.; Pichersky, E.; Zubieta, C.; Ferrer, J.L. Structural, functional and evolutionary basis for methylation of plant small molecules. Recent Adv. Phytochem. 2003, 37, 37–58. [Google Scholar]
- Schoch, G.; Goepfert, S.; Morant, M.; Hehn, A.; Meyer, D.; Ullmann, P.; Werck-Reichhart, D. CYP98A3 from Arabidopsis thaliana is a 3′-hydroxylase of phenolic esters, a missing link in the phenylpropanoid pathway. J. Biol. Chem. 2001, 276, 36566–36574. [Google Scholar] [CrossRef] [PubMed]
- Bassard, J.E.; Mutterer, J.; Duval, F.; Werck-Reichhart, D. A novel method for monitoring the localization of cytochromes P450 and other endoplasmic reticulum membrane associated proteins: A tool for investigating the formation of metabolons. FEBS J. 2012, 279, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Atila, M.; Katselis, G.; Chumala, P.; Luo, Y. Characterization of N-Succinylation of L-Lysylphosphatidylglycerol in Bacillus subtilis using tandem mass spectrometry. J. Am. Soc. Mass. Spectrom. 2016, 27, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Benaragama, I.; Meesapyodsuk, D.; Beattie, A.D.; Qiu, X. Identification and functional analysis of new peroxygenases in oat. Planta 2017, 246, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Obel, N.; Scheller, H.V. Enzymatic synthesis and purification of caffeoyl-CoA, p-coumaroyl-CoA and feruloyl-CoA. Anal. Biochem. 2000, 286, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Verardo, V.; Serea, C.; Segal, R.; Caboni, M.F. Free and bound minor polar compounds in oats: Different extraction methods and analytical determinations. J. Cereal Sci. 2011, 54, 211–217. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Km (μM) | Vmax (nmol/μg/min) | Vmax/Km (nmol/μg/mim/μM) |
|---|---|---|---|
| Avenanthramide C | 167.0 ± 25.2 | 514.4 ± 41.0 | 3.08 |
| Caffeoyl-CoA | 2319.0 ± 867.6 | 1620.0 ± 390.0 | 0.70 |
| Caffeic Acid | 1903.0 ± 173.5 | 597.6 ± 40.0 | 0.31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chen, Y.; Meesapyodsuk, D.; Qiu, X. The Biosynthetic Pathway of Major Avenanthramides in Oat. Metabolites 2019, 9, 163. https://doi.org/10.3390/metabo9080163
Li Z, Chen Y, Meesapyodsuk D, Qiu X. The Biosynthetic Pathway of Major Avenanthramides in Oat. Metabolites. 2019; 9(8):163. https://doi.org/10.3390/metabo9080163
Chicago/Turabian StyleLi, Zhiyong, Yi Chen, Dauenpen Meesapyodsuk, and Xiao Qiu. 2019. "The Biosynthetic Pathway of Major Avenanthramides in Oat" Metabolites 9, no. 8: 163. https://doi.org/10.3390/metabo9080163
APA StyleLi, Z., Chen, Y., Meesapyodsuk, D., & Qiu, X. (2019). The Biosynthetic Pathway of Major Avenanthramides in Oat. Metabolites, 9(8), 163. https://doi.org/10.3390/metabo9080163