
Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres


 ,
,  , , and
, , and 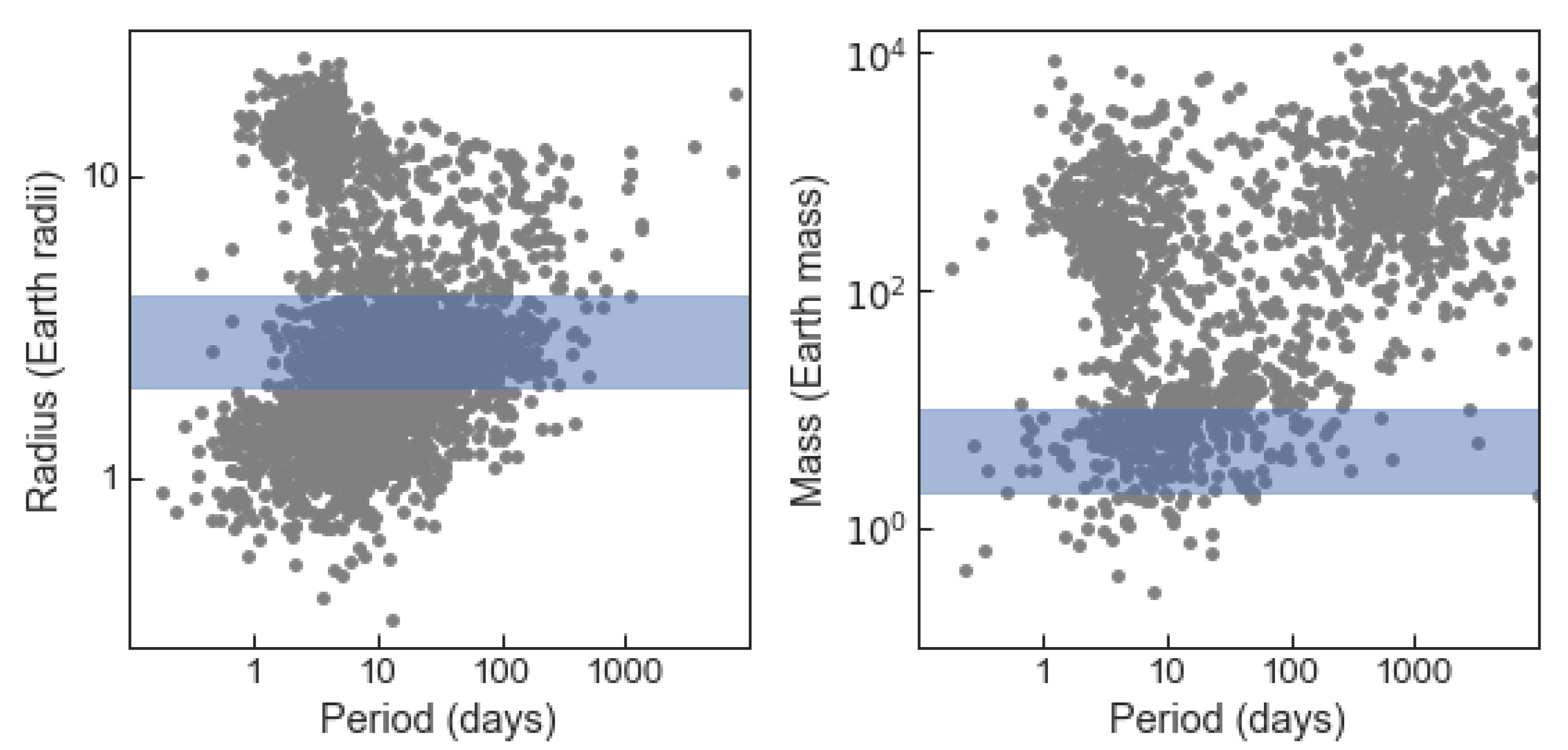{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Motivation to Expand the Pool of Planets Considered to Be Potentially Habitable
1.2. Earth’s Aerial Biosphere
1.3. Speculation on Solar System Planet and Brown Dwarf Aerial Biospheres
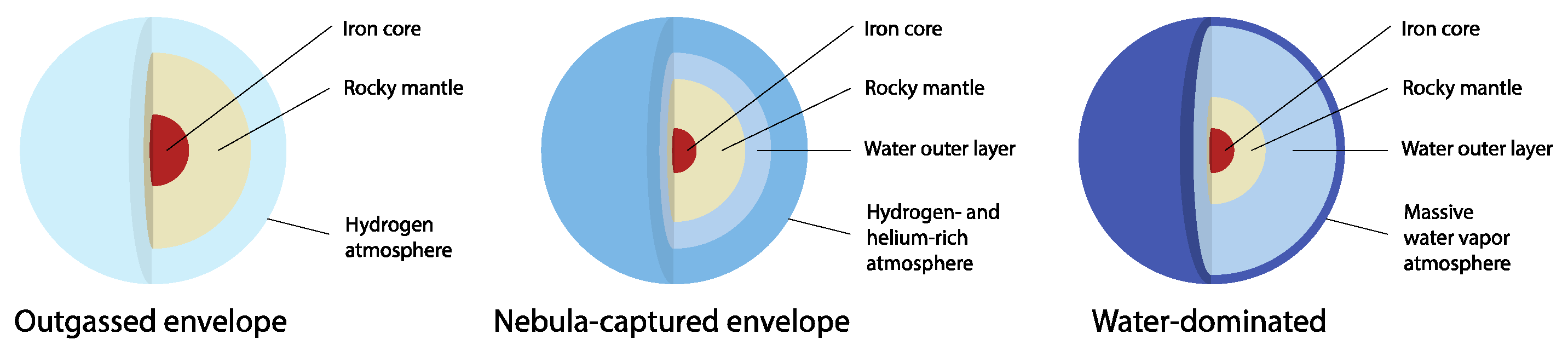
2. Sub Neptune Atmospheres Can Satisfy the Basic Requirements for Life
- A thermodynamic disequilibrium;
- An environment capable of maintaining covalent bonds (in chemical compounds), especially between carbon, hydrogen, and other atoms;
- A liquid environment;
- A molecular system that can support Darwinian evolution.
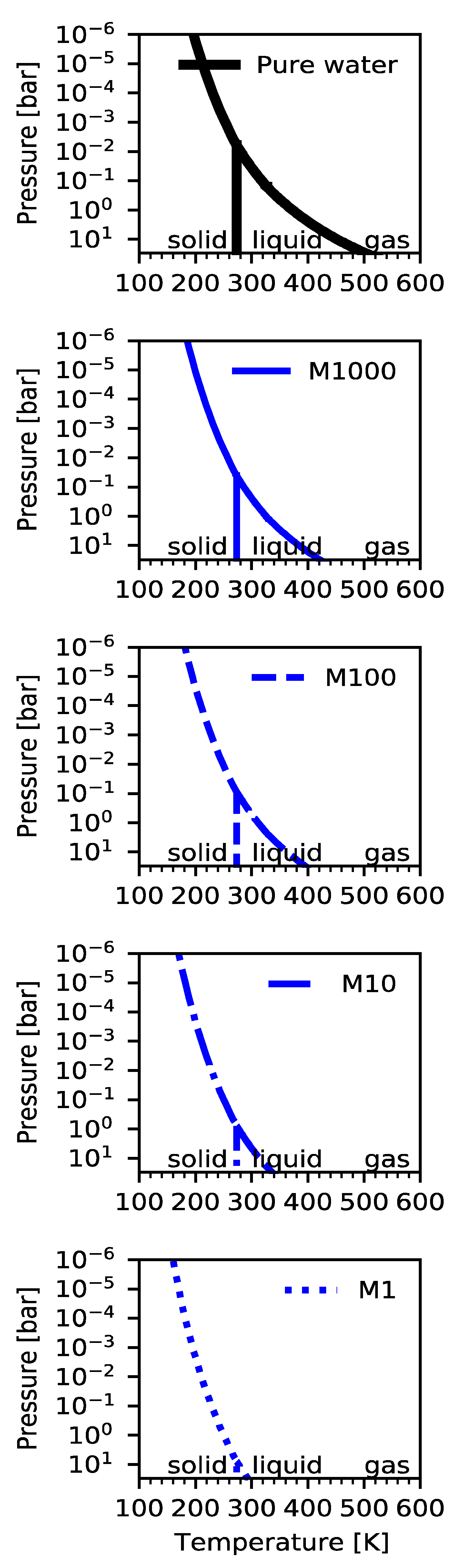
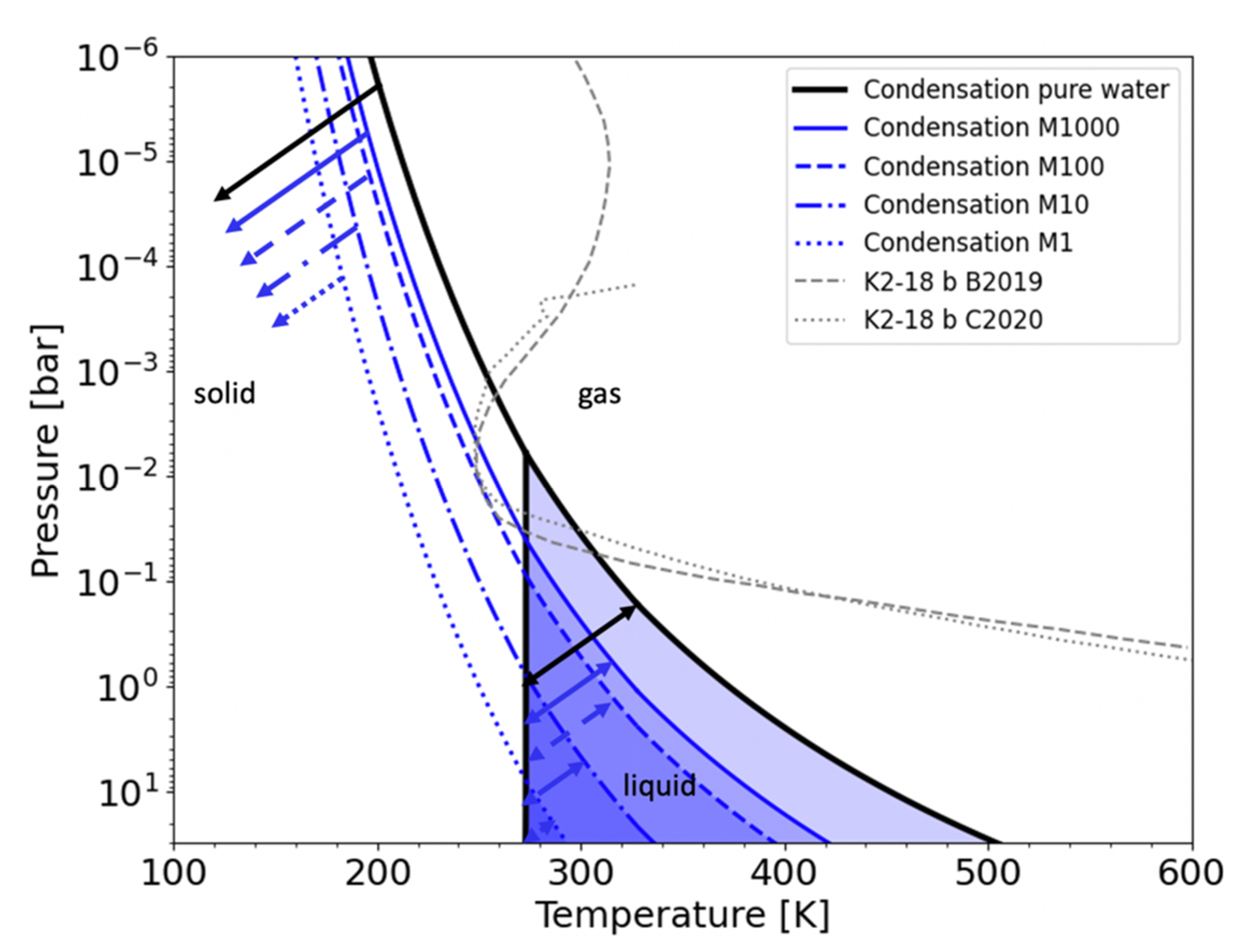
3. Atmospheric Liquid Water Clouds Require High Water Content or Cold Lower Layers
4. How Can Life Persist Aloft?
4.1. Can Passive Life Particles Spend Enough Time Amongst Liquid Cloud Droplets?
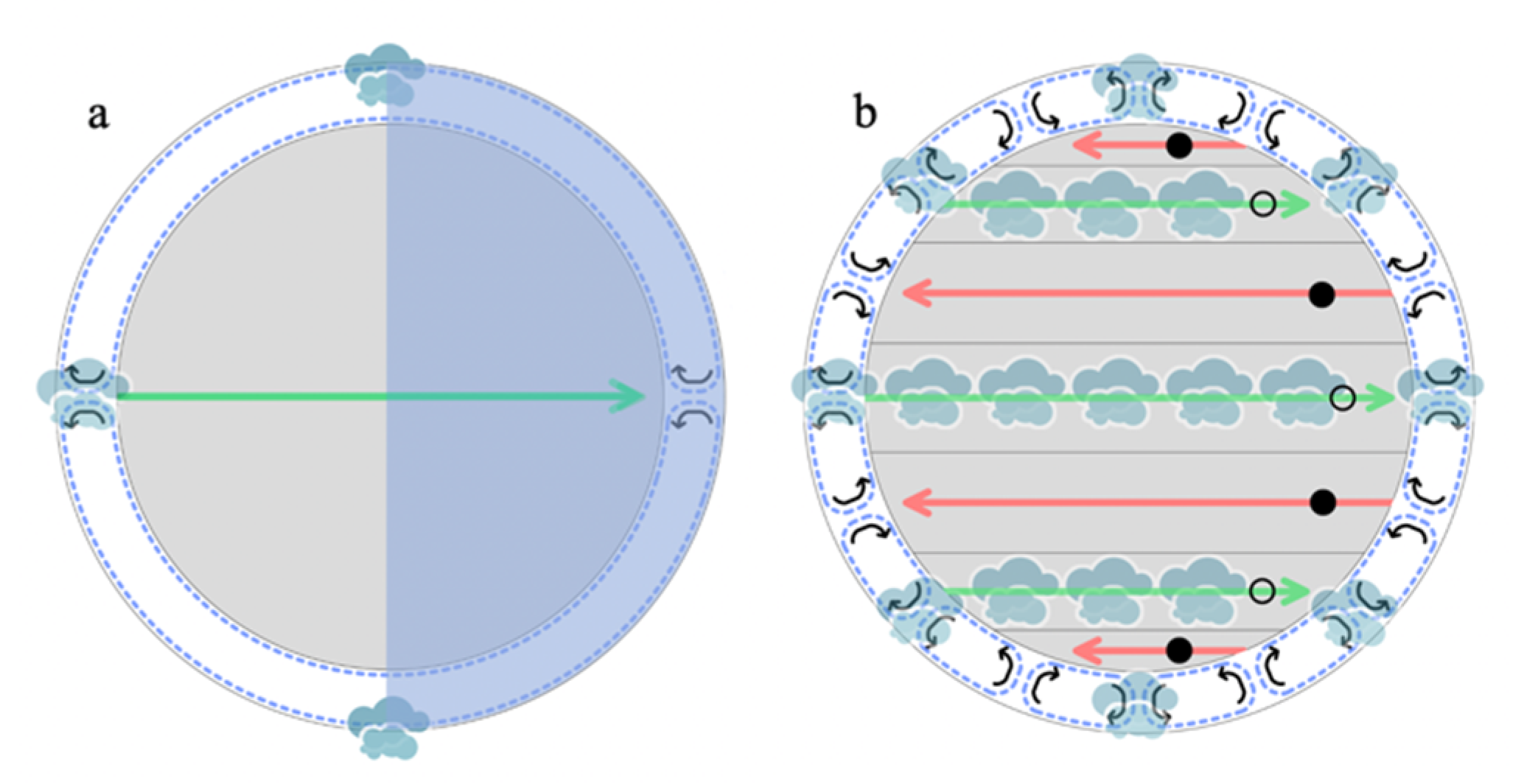
4.1.1. Cloud Location and Downward Transport in a Slow Rotator: K2-18b
4.1.2. Cloud Location and Downward Transport in Rapid Rotators
4.1.3. Other Physical Factors Affecting Aerial Life
4.2. Active Mechanisms for Complex Life to Persist Aloft
5. Challenges to Life in a Sub Neptune Atmosphere beyond Persistence Aloft
5.1. Origin of Life in an Atmosphere
5.2. Nutrient Scarcity
5.3. High-Energy Radiation or Lack Thereof for Planets Orbiting M Dwarf Stars
6. Possible Metabolic Strategies and Biosignatures
6.1. Hydrogenic Photosynthesis and CO as a Bioindicator
6.2. Methanogenesis
6.3. Light-Harvesting Pigments
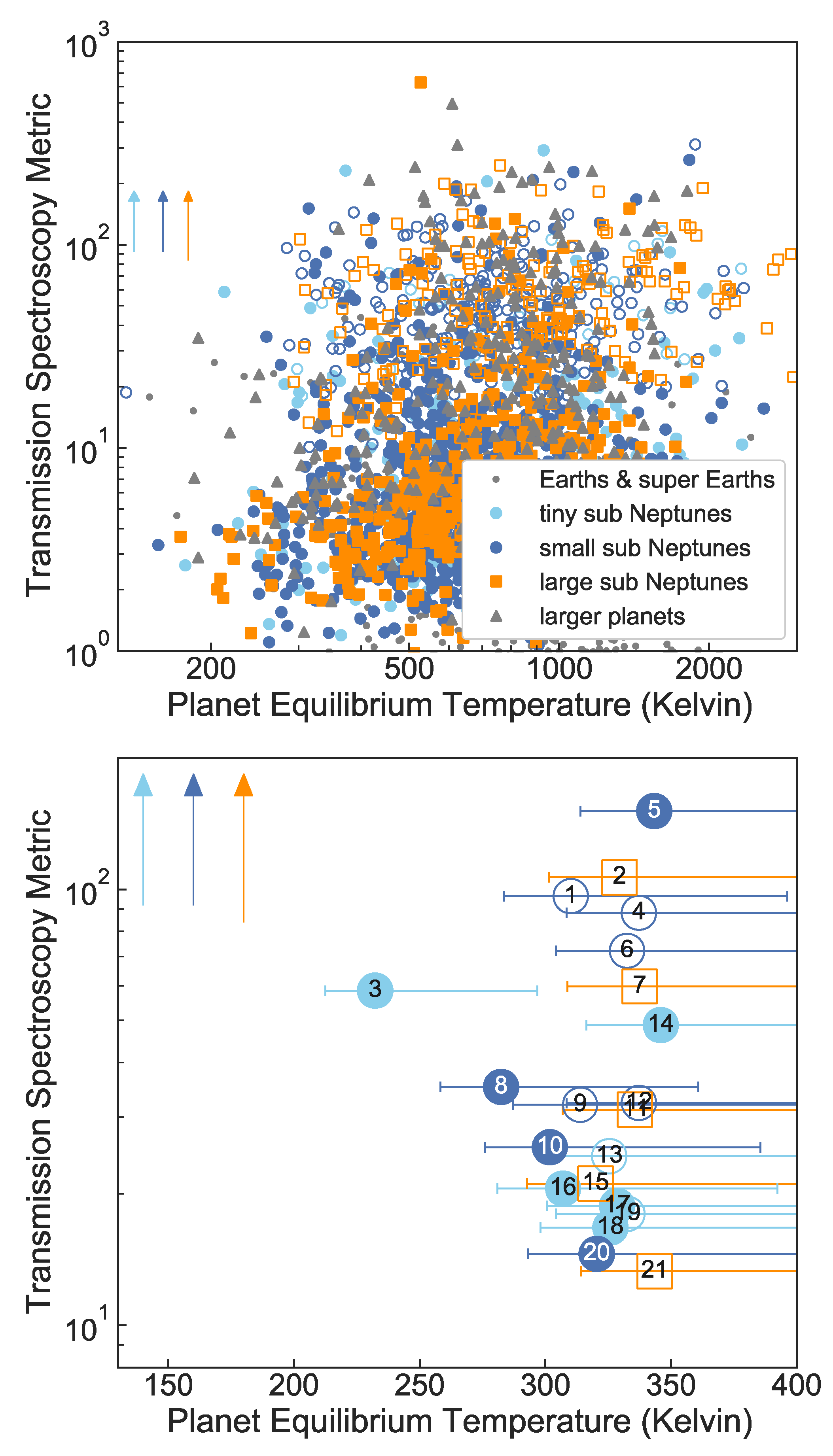
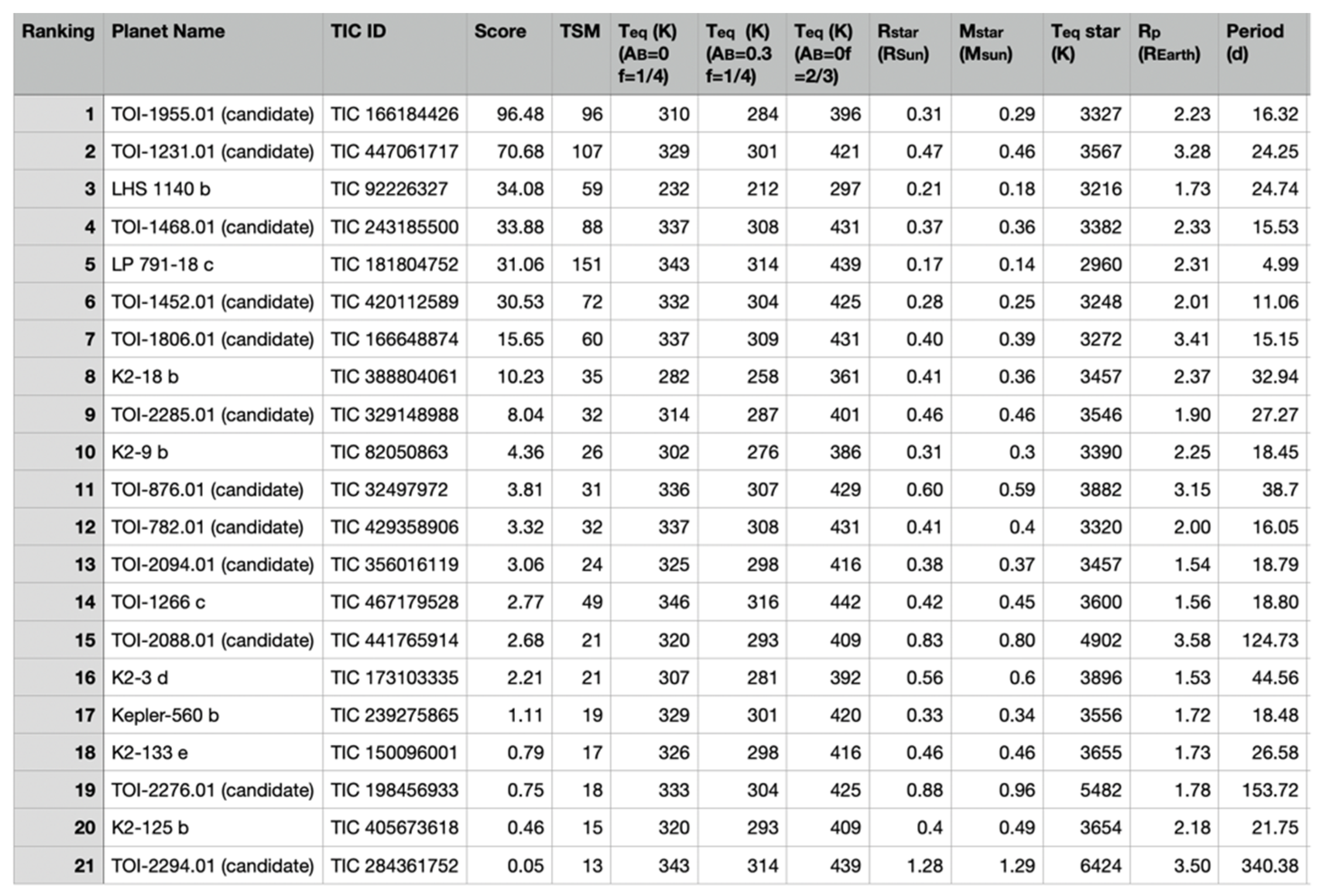
7. Target List of Temperate Sub Neptunes Suitable for Atmospheric Observations
7.1. Temperature and Scaled Semi-Major Axis Suitability Metric
7.2. Target Star List
- Rp > 1.5 REarth,
- 200 K Teq < 320 K, and
- TSM > 12.
- Rp 1.5 REarth “Earths and super Earths”;
- 1.5 REarth < Rp 1.8 REarth “tiny sub Neptunes”;
- 1.8 REarth < Rp 2.75 REarth “small sub Neptunes”;
- 2.75 REarth < Rp 4 REarth “large sub Neptunes”.
8. Summary and Future Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, J.P.; Mather, J.C.; Clampin, M.; Doyon, R.; Greenhouse, M.A.; Hammel, H.B.; Hutchings, J.B.; Jakobsen, P.; Lilly, S.J.; Long, K.S.; et al. The James Webb Space Telescope. Space Sci. Rev. 2006, 123, 485–606. [Google Scholar] [CrossRef] [Green Version]
- Gilmozzi, R.; Spyromilio, J. The European Extremely Large Telescope (E-ELT). Messenger 2007, 127, 11. [Google Scholar]
- Johns, M.; McCarthy, P.; Raybould, K.; Bouchez, A.; Farahani, A.; Filgueira, J.; Jacoby, G.; Shectman, S.; Sheehan, M. Giant magellan telescope: Overview. Int. Soc. Opt. Photonics 2012, 8444, 84441H. [Google Scholar]
- Sanders, G.H. The Thirty Meter Telescope (TMT): An International Observatory. J. Astrophys. Astron. 2013, 34, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Seager, S.; Turnbull, M.; Sparks, W.; Thomson, M.; Shaklan, S.B.; Roberge, A.; Kuchner, M.; Kasdin, N.J.; Domagal-Goldman, S.; Cash, W.; et al. The Exo-S probe class starshade mission. Tech. Instrum. Detect. Exopl. Vii 2015, 9605, 96050W. [Google Scholar] [CrossRef]
- Gaudi, B.S.; Seager, S.; Mennesson, B.; Kiessling, A.; Warfield, K.; Cahoy, K.; Clarke, J.T.; Domagal-Goldman, S.; Feinberg, L.; Guyon, O.; et al. The Habitable Exoplanet Observatory (HabEx) Mission Concept Study Final Report. arXiv 2020, arXiv:2001.06683. [Google Scholar]
- The LUVOIR Team The LUVOIR Mission Concept Study Final Report. arXiv 2019, arXiv:1912.06219.
- Meixner, M.; Cooray, A.; Leisawitz, D.; Staguhn, J.; Armus, L.; Battersby, C.; Bauer, J.; Bergin, E.; Bradford, C.M.; Ennico-Smith, K.; et al. Origins Space Telescope Mission Concept Study Report. arXiv 2019, arXiv:1912.06213. [Google Scholar]
- Adams, E.R.; Seager, S.; Elkins-Tanton, L. Ocean planet or thick atmosphere: On the mass-radius relationship for solid exoplanets with massive atmospheres. Astrophys. J. 2008, 673. [Google Scholar] [CrossRef] [Green Version]
- Lopez, E.D.; Fortney, J.J. Understanding the Mass-Radius Relation for Sub-neptunes: Radius as a Proxy for Composition. Astrophys. J. 2014, 792, 1. [Google Scholar] [CrossRef] [Green Version]
- Montet, B.T.; Morton, T.D.; Foreman-Mackey, D.; Johnson, J.A.; Hogg, D.W.; Bowler, B.P.; Latham, D.W.; Bieryla, A.; Mann, A.W. Stellar and planetary properties of K2 campaign 1 candidates and validation of 17 planets, including a planet receiving Earth-like insolation. Astrophys. J. 2015, 809, 25. [Google Scholar] [CrossRef] [Green Version]
- Benneke, B.; Wong, I.; Piaulet, C.; Knutson, H.A.; Lothringer, J.; Morley, C.V.; Crossfield, I.J.M.; Gao, P.; Greene, T.P.; Dressing, C. Water vapor and clouds on the habitable-zone sub-Neptune exoplanet K2-18b. Astrophys. J. Lett. 2019, 887, L14. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.A.; Seager, S. Three possible origins for the gas layer on GJ 1214B. Astrophys. J. 2010, 716. [Google Scholar] [CrossRef]
- Madhusudhan, N.; Nixon, M.C.; Welbanks, L.; Piette, A.A.A.; Booth, R.A. The Interior and Atmosphere of the Habitable-zone Exoplanet K2-18b. Astrophys. J. 2020, 891, L7. [Google Scholar] [CrossRef]
- Greene, T.P.; Line, M.R.; Montero, C.; Fortney, J.J.; Lustig-Yaeger, J.; Luther, K. Characterizing Transiting Exoplanet Atmospheres with JWST. Astrophys. J. 2016, 817, 17. [Google Scholar] [CrossRef] [Green Version]
- Howard, A.W.; Marcy, G.W.; Bryson, S.T.; Jenkins, J.M.; Rowe, J.F.; Batalha, N.M.; Borucki, W.J.; Koch, D.G.; Dunham, E.W.; Gautier, T.N.; et al. Planet occurrence within 0.25AU of solar-type stars from Kepler. Astrophys. J. Suppl. Ser. 2012, 201. [Google Scholar] [CrossRef] [Green Version]
- Dressing, C.D.; Charbonneau, D. The Occurrence Rate of Small Planets around Small Stars. Astrophys. J. 2013, 767, 95. [Google Scholar] [CrossRef] [Green Version]
- Fressin, F.; Torres, G.; Charbonneau, D.; Bryson, S.T.; Christiansen, J.; Dressing, C.D.; Jenkins, J.M.; Walkowicz, L.M.; Batalha, N.M. The False Positive Rate of Kepler and the Occurrence of Planets. Astrophys. J. 2013, 766, 81. [Google Scholar] [CrossRef] [Green Version]
- Bean, J.L.; Raymond, S.N.; Owen, J.E. The Nature and Origins of Sub-Neptune Size Planets. J. Geophys. Res. 2021, 126, e06639. [Google Scholar] [CrossRef]
- Rogers, L.A. Most 1.6 Earth-radius Planets are Not Rocky. Astrophys. J. 2015, 801, 41. [Google Scholar] [CrossRef] [Green Version]
- Kite, E.S.; Fegley, B., Jr.; Schaefer, L.; Ford, E.B. Atmosphere Origins for Exoplanet Sub-Neptunes. Astrophys. J. 2020, 891, 111. [Google Scholar] [CrossRef]
- Vazan, A.; Ormel, C.W.; Noack, L.; Dominik, C. Contribution of the Core to the Thermal Evolution of Sub-Neptunes. Astrophys. J. 2018, 869, 163. [Google Scholar] [CrossRef] [Green Version]
- Bodenheimer, P.; Lissauer, J.J. Accretion and Evolution of ~2.5 M⊕ Planets with Voluminous H/He Envelopes. Astrophys. J. 2014, 791, 103. [Google Scholar] [CrossRef] [Green Version]
- Bains, W. What do we think life is? A simple illustration and its consequences. Int. J. Astrobiol. 2014, 13, 101–111. [Google Scholar] [CrossRef]
- Baross, J.; Benner, S.A.; Cody, G.D.; Copley, S.D.; Pace, N.R.; Scott, J.H.; Shapiro, R.; Sogin, M.L.; Stein, J.L.; Summons, R.; et al. The Limits of Organic Life in Planetary Systems; National Academies Press: Washington, DC, USA, 2007; ISBN 0309179564. [Google Scholar]
- Ricker, G.R.; Winn, J.N.; Vanderspek, R.; Latham, D.W.; Bakos, G.Á.; Bean, J.L.; Berta-Thompson, Z.K.; Brown, T.M.; Buchhave, L.; Butler, N.R.; et al. Transiting Exoplanet Survey Satellite (TESS). J. Astron. Telesc. Instrum. Syst. 2015, 1, 14003. [Google Scholar] [CrossRef] [Green Version]
- Vaïtilingom, M.; Attard, E.; Gaiani, N.; Sancelme, M.; Deguillaume, L.; Flossmann, A.I.; Amato, P.; Delort, A.-M. Long-term features of cloud microbiology at the puy de Dôme (France). Atmos. Environ. 2012, 56, 88–100. [Google Scholar] [CrossRef]
- Amato, P.; Joly, M.; Besaury, L.; Oudart, A.; Taib, N.; Moné, A.I.; Deguillaume, L.; Delort, A.-M.; Debroas, D. Active microorganisms thrive among extremely diverse communities in cloud water. PLoS ONE 2017, 12, e0182869. [Google Scholar] [CrossRef] [Green Version]
- Barberán, A.; Ladau, J.; Leff, J.W.; Pollard, K.S.; Menninger, H.L.; Dunn, R.R.; Fierer, N. Continental-scale distributions of dust-associated bacteria and fungi. Proc. Natl. Acad. Sci. USA 2015, 112, 5756–5761. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.; Gonzalez-Martin, C.; Hoose, C.; Smith, D. Global-scale atmospheric dispersion of microorganisms. Microbiol. Aerosols 2017, 155–194. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Gosewinkel, U.; Starnawski, P.; Lever, M.; Finster, K. Aeolian dispersal of bacteria in southwest Greenland: Their sources, abundance, diversity and physiological states. FEMS Microbiol. Ecol. 2018, 94, fiy031. [Google Scholar] [CrossRef] [Green Version]
- Burrows, S.M.; Butler, T.; Jöckel, P.; Tost, H.; Kerkweg, A.; Pöschl, U.; Lawrence, M.G. Bacteria in the global atmosphere–Part 2: Modeling of emissions and transport between different ecosystems. Atmos. Chem. Phys. 2009, 9, 9281–9297. [Google Scholar] [CrossRef] [Green Version]
- Bowers, R.M.; McCubbin, I.B.; Hallar, A.G.; Fierer, N. Seasonal variability in airborne bacterial communities at a high-elevation site. Atmos. Environ. 2012, 50, 41–49. [Google Scholar] [CrossRef]
- Amato, P.; Ménager, M.; Sancelme, M.; Laj, P.; Mailhot, G.; Delort, A.-M. Microbial population in cloud water at the Puy de Dôme: Implications for the chemistry of clouds. Atmos. Environ. 2005, 39, 4143–4153. [Google Scholar] [CrossRef] [Green Version]
- DeLong, E.F. Diminutive Cells in The Oceans—Unanswered Questions. In Size Limits of Very Small Microorganisms: Proceedings of a Workshop; Monterey Bay Aquarium Research Institute: Moss Landing, CA, USA, 1999; p. 81. [Google Scholar]
- Seager, S.; Petkowski, J.J.; Gao, P.; Bains, W.; Bryan, N.C.; Ranjan, S.; Greaves, J. The Venusian Lower Atmosphere Haze as a Depot for Desiccated Microbial Life: A Proposed Life Cycle for Persistence of the Venusian Aerial Biosphere. Astrobiology 2020. [Google Scholar] [CrossRef]
- Bryan, N.C.; Christner, B.C.; Guzik, T.G.; Granger, D.J.; Stewart, M.F. Abundance and survival of microbial aerosols in the troposphere and stratosphere. ISME J. 2019, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Amato, P.; Besaury, L.; Joly, M.; Penaud, B.; Deguillaume, L.; Delort, A.-M. Metatranscriptomic exploration of microbial functioning in clouds. Sci. Rep. 2019, 9, 4383. [Google Scholar] [CrossRef] [Green Version]
- Dimmick, R.L.; Wolochow, H.; Chatigny, M.A. Evidence for more than one division of bacteria within airborne particles. Appl. Environ. Microbiol. 1979, 38, 642–643. [Google Scholar] [CrossRef] [Green Version]
- Dimmick, R.L.; Wolochow, H.; Chatigny, M.A. Evidence that bacteria can form new cells in airborne particles. Appl. Environ. Microbiol. 1979, 37, 924–927. [Google Scholar] [CrossRef] [Green Version]
- Sattler, B.; Puxbaum, H.; Psenner, R. Bacterial growth in supercooled cloud droplets. Geophys. Res. Lett. 2001, 28, 239–242. [Google Scholar] [CrossRef]
- Morowitz, H.; Sagan, C. Life in the Clouds of Venus? Nature 1967, 215, 1259. [Google Scholar] [CrossRef]
- Limaye, S.S.; Mogul, R.; Smith, D.J.; Ansari, A.H.; Słowik, G.P.; Vaishampayan, P. Venus’ Spectral Signatures and the Potential for Life in the Clouds. Astrobiology 2018, 18, 1181–1198. [Google Scholar] [CrossRef]
- Cavalazzi, B.; Barbieri, R.; Gómez, F.; Capaccioni, B.; Olsson-Francis, K.; Pondrelli, M.; Rossi, A.P.; Hickman-Lewis, K.; Agangi, A.; Gasparotto, G.; et al. The Dallol Geothermal Area, Northern Afar (Ethiopia)-An Exceptional Planetary Field Analog on Earth. Astrobiology 2019, 19, 553–578. [Google Scholar] [CrossRef]
- Kotopoulou, E.; Delgado Huertas, A.; Garcia-Ruiz, J.M.; Dominguez-Vera, J.M.; Lopez-Garcia, J.M.; Guerra-Tschuschke, I.; Rull, F. A polyextreme hydrothermal system controlled by iron: The case of Dallol at the Afar Triangle. ACS Earth Sp. Chem. 2018, 3, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagan, C.; Salpeter, E.E. Particles, environments and possible ecologies in the Jovian atmosphere. Astrophys. J. Suppl. Ser. 1976, 32, 737–755. [Google Scholar] [CrossRef] [Green Version]
- Yates, J.S.; Palmer, P.I.; Biller, B.; Cockell, C.S. Atmospheric habitable zones in Y dwarf atmospheres. Astrophys. J. 2017, 836, 184. [Google Scholar] [CrossRef] [Green Version]
- Lingam, M.; Loeb, A. Brown Dwarf atmospheres as the potentially most detectable and abundant sites for life. Astrophys. J. 2019, 883, 143. [Google Scholar] [CrossRef] [Green Version]
- Tsiaras, A.; Waldmann, I.P.; Tinetti, G.; Tennyson, J.; Yurchenko, S.N. Water vapour in the atmosphere of the habitable-zone eight-Earth-mass planet K2-18 b. Nat. Astron. 2019, 3, 1086–1091. [Google Scholar] [CrossRef] [Green Version]
- Blain, D.; Charnay, B.; Bézard, B. 1D atmospheric study of the temperate sub-Neptune K2-18b. Astron. Astrophys. 2021, 646, A15. [Google Scholar] [CrossRef]
- Charnay, B.; Blain, D.; Bézard, B.; Leconte, J.; Turbet, M.; Falco, A. Formation and dynamics of water clouds on temperate sub-Neptunes: The example of K2-18b. Astron. Astrophys. 2021, 646, A171. [Google Scholar] [CrossRef]
- Bains, W.; Xiao, Y.; Yu, C. Prediction of the maximum temperature for life based on the stability of metabolites to decomposition in water. Life 2015, 5, 1054–1100. [Google Scholar] [CrossRef] [Green Version]
- McKay, C.P. Requirements and limits for life in the context of exoplanets. Proc. Natl. Acad. Sci. USA 2014, 111, 12628–12633. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [Green Version]
- Kashefi, K.; Lovley, D.R. Extending the upper temperature limit for life. Science 2003, 301, 934. [Google Scholar] [CrossRef]
- Hoehler, T.; Bains, W.; Davila, A.; Parenteau, M.; Pohorille, A. Life’s requirements, habitability, and biological potential. In Planetary Astrobiology; Meadows, V., Marais, D.J., Des Arney, G., Schmidt, B., Eds.; The University of Arizona Press: Tucson, AZ, USA, 2020. [Google Scholar]
- Petkowski, J.J.; Bains, W.; Seager, S. On the Potential of Silicon as a Building Block for Life. Life 2020, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Seager, S.; Huang, J.; Petkowski, J.J.; Pajusalu, M. Laboratory studies on the viability of life in H2-dominated exoplanet atmospheres. Nat. Astron. 2020. [Google Scholar] [CrossRef]
- Engel, T.; Reid, P. Thermodynamics, 4th ed.; Pearson: New York, NY, USA, 2018; ISBN 978-0134804583. [Google Scholar]
- Lodders, K.; Fegley, B. The Planetary Scientist’s Companion/Katharina Lodders, Bruce Fegley; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Piette, A.A.A.; Madhusudhan, N. On the Temperature Profiles and Emission Spectra of Mini-Neptune Atmospheres. Astrophys. J. 2020, 904, 154. [Google Scholar] [CrossRef]
- Pierrehumbert, R.T.; Hammond, M. Atmospheric Circulation of Tide-Locked Exoplanets. Annu. Rev. Fluid Mech. 2019, 51, 275–303. [Google Scholar] [CrossRef]
- Guillot, T. Condensation of methane, ammonia, and water and the inhibition of convection in giant planets. Science 1995, 269, 1697–1699. [Google Scholar] [CrossRef] [PubMed]
- Leconte, J.; Selsis, F.; Hersant, F.; Guillot, T. Condensation-inhibited convection in hydrogen-rich atmospheres. Astron. Astrophys. 2017, 598, A98. [Google Scholar] [CrossRef]
- Fievet, A.; Ducret, A.; Mignot, T.; Valette, O.; Robert, L.; Pardoux, R.; Dolla, A.R.; Aubert, C. Single-cell analysis of growth and cell division of the anaerobe Desulfovibrio vulgaris Hildenborough. Front. Microbiol. 2015, 6, 1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldreich, P.; Soter, S. Q in the Solar System. Icarus 1966, 5, 375–389. [Google Scholar] [CrossRef]
- Guinan, E.F.; Engle, S.G. The K2-18b Planetary System: Estimates of the Age and X-UV Irradiances of a Habitable Zone “Wet” Sub-Neptune Planet. Res. Notes Am. Astron. Soc. 2019, 3, 189. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, K.; Schubert, G.; Anderson, J.D. Origin of Jupiter’s cloud-level zonal winds remains a puzzle even after Juno. Proc. Natl. Acad. Sci. USA 2018, 115, 8499–8504. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, L.N.; Kaspi, Y.; Guillot, T.; Showman, A.P. How Well Do We Understand the Belt/Zone Circulation of Giant Planet Atmospheres? Space Sci. Rev. 2020, 216, 30. [Google Scholar] [CrossRef] [Green Version]
- Showman, A.P.; Tan, X.; Parmentier, V. Atmospheric Dynamics of Hot Giant Planets and Brown Dwarfs. Space Sci. Rev. 2020, 216, 139. [Google Scholar] [CrossRef]
- West, R.A.; Friedson, A.J.; Appleby, J.F. Jovian large-scale stratospheric circulation. Icarus 1992, 100, 245–259. [Google Scholar] [CrossRef]
- Walters, K.R.; Serianni, A.S.; Sformo, T.; Barnes, B.M.; Duman, J.G. A nonprotein thermal hysteresis-producing xylomannan antifreeze in the freeze-tolerant Alaskan beetle Upis ceramboides. Proc. Natl. Acad. Sci. USA 2009, 106, 20210–20215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mykytczuk, N.C.S.; Foote, S.J.; Omelon, C.R.; Southam, G.; Greer, C.W.; Whyte, L.G. Bacterial growth at −15 °C; molecular insights from the permafrost bacterium Planococcus halocryophilus Or1. ISME J. 2013, 7, 1211–1226. [Google Scholar] [CrossRef] [Green Version]
- Maykut, G.; Untersteiner, N. The Geophysics of Sea Ice; Plenium: New York, NY, USA, 1986. [Google Scholar]
- Junge, K.; Eicken, H.; Deming, J.W. Motility of Colwellia psychrerythraea strain 34H at subzero temperatures. Appl. Environ. Microbiol. 2003, 69, 4282–4284. [Google Scholar] [CrossRef] [Green Version]
- Huston, A.L. Bacterial Adaptation to the Cold: In Situ Activities of Extracellular Enzymes in the North Water Polynya and Characterization of a Cold-Active Aminopeptidase from Colwellia Psychrerythraea Strain 34H. Ph.D. Thesis, University of Washington, Washington, DC, USA, 2004. [Google Scholar]
- Farrell, A.H.; Hohenstein, K.A.; Shain, D.H. Molecular adaptation in the ice worm, Mesenchytraeus solifugus: Divergence of energetic-associated genes. J. Mol. Evol. 2004, 59, 666–673. [Google Scholar] [CrossRef]
- Shain, D.H.; Mason, T.A.; Farrell, A.H.; Michalewicz, L.A. Distribution and behavior of ice worms (Mesenchytraeus solifugus) in south-central Alaska. Can. J. Zool. 2001, 79, 1813–1821. [Google Scholar] [CrossRef]
- Shain, D.H.; Carter, M.R.; Murray, K.P.; Maleski, K.A.; Smith, N.R.; McBride, T.R.; Michalewicz, L.A.; Saidel, W.M. Morphologic characterization of the ice worm Mesenchytraeus solifugus. J. Morphol. 2000, 246, 192–197. [Google Scholar] [CrossRef]
- Cáceres, L.; Gómez-Silva, B.; Garró, X.; Rodríguez, V.; Monardes, V.; McKay, C.P. Relative humidity patterns and fog water precipitation in the Atacama Desert and biological implications. J. Geophys. Res. Biogeosciences 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Gruzdev, N.; McClelland, M.; Porwollik, S.; Ofaim, S.; Pinto, R.; Saldinger-Sela, S. Global Transcriptional Analysis of Dehydrated Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2012, 78, 7866–7875. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Guidetti, R.; JoÈnsson, K.I. Long-term anhydrobiotic survival in semi-terrestrial micrometazoans. J. Zool. 2002, 257, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Zagorodnov, V.; Sandman, K.; Reeve, J.N. Recovery and Identification of Viable Bacteria Immured in Glacial Ice. Icarus 2000, 144, 479–485. [Google Scholar] [CrossRef]
- Cano, R.J.; Borucki, M.K. Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber. Science 1995, 268, 1060–1064. [Google Scholar] [CrossRef]
- Meng, F.-W.; Wang, X.-Q.; Ni, P.; Kletetschka, G.; Yang, C.-H.; Li, Y.-P.; Yang, Y.-H. A newly isolated Haloalkaliphilic bacterium from middle–late Eocene halite formed in salt lakes in China. Carbonates Evaporites 2015, 30, 321–330. [Google Scholar] [CrossRef]
- Burch, A.Y.; Zeisler, V.; Yokota, K.; Schreiber, L.; Lindow, S.E. The hygroscopic biosurfactant syringafactin produced by P seudomonas syringae enhances fitness on leaf surfaces during fluctuating humidity. Environ. Microbiol. 2014, 16, 2086–2098. [Google Scholar] [CrossRef]
- Elliot, M.A.; Talbot, N.J. Building filaments in the air: Aerial morphogenesis in bacteria and fungi. Curr. Opin. Microbiol. 2004, 7, 594–601. [Google Scholar] [CrossRef]
- Talbot, N.J. Coming up for air and sporulation. Nature 1999, 398, 295–296. [Google Scholar] [CrossRef]
- Hedenström, A.; Norevik, G.; Warfvinge, K.; Andersson, A.; Bäckman, J.; Åkesson, S. Annual 10-month aerial life phase in the common swift Apus apus. Curr. Biol. 2016, 26, 3066–3070. [Google Scholar] [CrossRef] [Green Version]
- Lyons, T.W.; Rogers, K.; Krishnamurthy, R.; Williams, L.; Marchi, S.; Schwieterman, E.; Planavsky, N.; Reinhard, C. Constraining Prebiotic Chemistry Through a Better Understanding of Earth’s Earliest Environments. arXiv 2020, arXiv:2008.04803. [Google Scholar]
- Bains, W. Getting Beyond the Toy Domain. Meditations on David Deamer’s “Assembling Life”. Life 2020, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R. Origins: A Skeptic’s Guide to the Creation of Life on Earth; Bantam Dell Pub Group: New York, NY, USA, 1987; ISBN 0553343556. [Google Scholar]
- Benner, S.A.; Bell, E.A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.; Mojzsis, S.J.; Omran, A.; Pasek, M.A.; Trail, D. When Did Life Likely Emerge on Earth in an RNA-First Process? ChemSystemsChem 2019, 2, e1900035. [Google Scholar]
- Furukawa, Y.; Chikaraishi, Y.; Ohkouchi, N.; Ogawa, N.O.; Glavin, D.P.; Dworkin, J.P.; Abe, C.; Nakamura, T. Extraterrestrial ribose and other sugars in primitive meteorites. Proc. Natl. Acad. Sci. USA 2019, 116, 24440–24445. [Google Scholar] [CrossRef] [Green Version]
- Callahan, M.P.; Smith, K.E.; Cleaves, H.J.; Ruzicka, J.; Stern, J.C.; Glavin, D.P.; House, C.H.; Dworkin, J.P. Carbonaceous meteorites contain a wide range of extraterrestrial nucleobases. Proc. Natl. Acad. Sci. USA 2011, 108, 13995–13998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Fischer, R.A.; Stixrude, L.P.; Lithgow-Bertelloni, C.R. Constraining the Volume of Earth’s Early Oceans With a Temperature-Dependent Mantle Water Storage Capacity Model. AGU Adv. 2021, 2, e2020AV000323. [Google Scholar] [CrossRef]
- Woese, C.R. A proposal concerning the origin of life on the planet earth. J. Mol. Evol. 1979, 13, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.; Schneider, C.; Okamura, H.; Crisp, A.; Amatov, T.; Dejmek, M.; Carell, T. Wet-dry cycles enable the parallel origin of canonical and non-canonical nucleosides by continuous synthesis. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Gnanamani, E.; Yan, X.; Zare, R.N. Can all bulk-phase reactions be accelerated in microdroplets? Analyst 2017, 142, 1399–1402. [Google Scholar] [CrossRef]
- Nam, I.; Lee, J.K.; Nam, H.G.; Zare, R.N. Abiotic production of sugar phosphates and uridine ribonucleoside in aqueous microdroplets. Proc. Natl. Acad. Sci. USA 2017, 114, 12396–12400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, E.C.; Vaida, V. In situ observation of peptide bond formation at the water-air interface. Proc. Natl. Acad. Sci. USA 2012, 109, 15697–15701. [Google Scholar] [CrossRef] [Green Version]
- Toupance, G.; Raulin, F.; Buvet, R. Formation of prebiochemical compounds in models of the primitive Earth’s atmosphere. Orig. Life 1975, 6, 83–90. [Google Scholar] [CrossRef]
- Shi, W.; Chance, M.R. Metalloproteomics: Forward and reverse approaches in metalloprotein structural and functional characterization. Curr. Opin. Chem. Biol. 2011, 15, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Peucker-Ehrenbrink, B. Accretion of extraterrestrial matter during the last 80 million years and its effect on the marine osmium isotope record. Geochim. Cosmochim. Acta 1996, 60, 3187–3196. [Google Scholar] [CrossRef]
- Emiliani, C. Planet. Earth: Cosmology, Geology, and the Evolution of Life and Environment; Cambridge University Press: Cambridge, UK, 1992; ISBN 9780521401234. [Google Scholar]
- Fung, I.Y.; Meyn, S.K.; Tegen, I.; Doney, S.C.; John, J.G.; Bishop, J.K.B. Iron supply and demand in the upper ocean. Glob. Biogeochem. Cycles 2000, 14, 281–295. [Google Scholar] [CrossRef]
- Li, J.; Mara, P.; Schubotz, F.; Sylvan, J.B.; Burgaud, G.; Klein, F.; Beaudoin, D.; Wee, S.Y.; Dick, H.J.B.; Lott, S. Recycling and metabolic flexibility dictate life in the lower oceanic crust. Nature 2020, 579, 250–255. [Google Scholar] [CrossRef]
- Oza, A.V.; Johnson, R.E.; Lellouch, E.; Schmidt, C.; Schneider, N.; Huang, C.; Gamborino, D.; Gebek, A.; Wyttenbach, A.; Demory, B.-O.; et al. Sodium and Potassium Signatures of Volcanic Satellites Orbiting Close-in Gas Giant Exoplanets. Astrophys. J. 2019, 885, 168. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.; Bagenal, F.; Hill, T.W.; Wilson, J.K. The Io neutral clouds and plasma torus. Jupit. Planet Satell. Magnetos. 2004, 1, 561–591. [Google Scholar]
- Tucker, O.J.; Johnson, R.E.; Young, L.A. Gas transfer in the Pluto-Charon system: A Charon atmosphere. Icarus 2015, 246, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Heller, R.; Williams, D.; Kipping, D.; Limbach, M.A.; Turner, E.; Greenberg, R.; Sasaki, T.; Bolmont, É.; Grasset, O.; Lewis, K.; et al. Formation, Habitability, and Detection of Extrasolar Moons. Astrobiology 2014, 14, 798–835. [Google Scholar] [CrossRef] [Green Version]
- Hoehler, T.M. Biological energy requirements as quantitative boundary conditions for life in the subsurface. Geobiology 2004, 2, 205–215. [Google Scholar] [CrossRef]
- Ranjan, S.; Wordsworth, R.; Sasselov, D.D. The surface UV environment on planets orbiting M dwarfs: Implications for prebiotic chemistry and the need for experimental follow-up. Astrophys. J. 2017, 843, 110. [Google Scholar] [CrossRef] [Green Version]
- Rimmer, P.B.; Xu, J.; Thompson, S.J.; Gillen, E.; Sutherland, J.D.; Queloz, D. The origin of RNA precursors on exoplanets. Sci. Adv. 2018, 4, eaar3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günther, M.N.; Zhan, Z.; Seager, S.; Rimmer, P.B.; Ranjan, S.; Stassun, K.G.; Oelkers, R.J.; Daylan, T.; Newton, E.; Kristiansen, M.H.; et al. Stellar Flares from the First TESS Data Release: Exploring a New Sample of M Dwarfs. Astron. J. 2020, 159, 60. [Google Scholar] [CrossRef] [Green Version]
- Segura, A.; Walkowicz, L.M.; Meadows, V.; Kasting, J.; Hawley, S. The effect of a strong stellar flare on the atmospheric chemistry of an Earth-like planet orbiting an M dwarf. Astrobiology 2010, 10, 751–771. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, M.A.; Weinberger, A.J.; Loyd, R.O.P.; Shkolnik, E.; Barclay, T.; Howard, W.S.; Zic, A.; Osten, R.A.; Cranmer, S.R.; Kowalski, A.F.; et al. Discovery of an Extremely Short Duration Flare from Proxima Centauri Using Millimeter through Far-ultraviolet Observations. Astrophys. J. 2021, 911, L25. [Google Scholar] [CrossRef]
- Ranjan, S.; Sasselov, D.D. Influence of the UV Environment on the Synthesis of Prebiotic Molecules. Astrobiology 2016, 16, 68–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.M.; Keck, J.L.; Battista, J.R. Rising from the Ashes: DNA Repair in Deinococcus radiodurans. PLoS Genet. 2010, 6, e1000815. [Google Scholar] [CrossRef] [PubMed]
- Diaz, B.; Schulze-Makuch, D. Microbial Survival Rates of Escherichia coli and Deinococcus radiodurans Under Low Temperature, Low Pressure, and UV-Irradiation Conditions, and Their Relevance to Possible Martian Life. Astrobiology 2006, 6, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Hawley, S.L.; Davenport, J.R.A.; Kowalski, A.F.; Wisniewski, J.P.; Hebb, L.; Deitrick, R.; Hilton, E.J. Kepler Flares. I. Active and Inactive M Dwarfs. Astrophys. J. 2014, 797, 121. [Google Scholar] [CrossRef] [Green Version]
- Newton, E.R.; Irwin, J.; Charbonneau, D.; Berlind, P.; Calkins, M.L.; Mink, J. The Hα Emission of Nearby M Dwarfs and its Relation to Stellar Rotation. Astrophys. J. 2017, 834, 85. [Google Scholar] [CrossRef]
- Owen, J.E.; Wu, Y. Kepler Planets: A Tale of Evaporation. Astrophys. J. 2013, 775, 105. [Google Scholar] [CrossRef] [Green Version]
- Mordasini, C. Planetary evolution with atmospheric photoevaporation. I. Analytical derivation and numerical study of the evaporation valley and transition from super-Earths to sub-Neptunes. Astron. Astrophys. 2020, 638, A52. [Google Scholar] [CrossRef]
- Atri, D.; Mogan, S.R.C. Stellar flares versus luminosity: XUV-induced atmospheric escape and planetary habitability. Mon. Not. R. Astron. Soc. 2021, 500, L1–L5. [Google Scholar] [CrossRef]
- Seager, S.; Bains, W.; Petkowski, J.J. Toward a List of Molecules as Potential Biosignature Gases for the Search for Life on Exoplanets and Applications to Terrestrial Biochemistry. Astrobiology 2016, 16, 465–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwieterman, E.W.; Kiang, N.Y.; Parenteau, M.N.; Harman, C.E.; DasSarma, S.; Fisher, T.M.; Arney, G.N.; Hartnett, H.E.; Reinhard, C.T.; Olson, S.L.; et al. Exoplanet Biosignatures: A Review of Remotely Detectable Signs of Life. Astrobiology 2018, 18, 663–708. [Google Scholar] [CrossRef]
- Seager, S.; Schrenk, M.; Bains, W. An Astrophysical View of Earth-Based Metabolic Biosignature Gases. Astrobiology 2012, 12, 61–82. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.R. The potential feasibility of chlorinic photosynthesis on exoplanets. Astrobiology 2010, 10, 953–963. [Google Scholar] [CrossRef]
- Seager, S.; Bains, W.; Hu, R. Biosignature Gases in H2-dominated Atmospheres on Rocky Exoplanets. Astrophys. J. 2013, 777, 95. [Google Scholar] [CrossRef] [Green Version]
- Bains, W.; Seager, S.; Zsom, A. Photosynthesis in hydrogen-dominated atmospheres. Life 2014, 4, 716–744. [Google Scholar] [CrossRef] [Green Version]
- Valmalette, J.C.; Dombrovsky, A.; Brat, P.; Mertz, C.; Capovilla, M.; Robichon, A. Light-induced electron transfer and ATP synthesis in a carotene synthesizing insect. Sci. Rep. 2012, 2, 579. [Google Scholar] [CrossRef] [Green Version]
- Moses, J.I.; Line, M.R.; Visscher, C.; Richardson, M.R.; Nettelmann, N.; Fortney, J.J.; Barman, T.S.; Stevenson, K.B.; Madhusudhan, N. Compositional Diversity in the Atmospheres of Hot Neptunes, with Application to GJ 436B. Astrophys. J. 2013, 777, 34. [Google Scholar] [CrossRef] [PubMed]
- Hoehler, T.M.; Alperin, M.J.; Albert, D.B.; Martens, C.S. Apparent minimum free energy requirements for methanogenic Archaea and sulfate-reducing bacteria in an anoxic marine sediment. FEMS Microbiol. Ecol. 2001, 38, 33–41. [Google Scholar] [CrossRef]
- Hu, R.; Seager, S.; Bains, W. Photochemistry in terrestrial exoplanet atmospheres. I. Photochemistry model and benchmark cases. Astrophys. J. 2012, 761, 166. [Google Scholar] [CrossRef]
- Mollière, P.; Snellen, I.A.G. Detecting isotopologues in exoplanet atmospheres using ground-based high-dispersion spectroscopy. Astron. Astrophys. 2019, 622, A139. [Google Scholar] [CrossRef] [Green Version]
- Arnold, L.; Gillet, S.; Lardière, O.; Riaud, P.; Schneider, J. A test for the search for life on extrasolar planets. Astron. Astrophys. 2002, 392, 231–237. [Google Scholar] [CrossRef]
- Woolf, N.J.; Smith, P.S.; Traub, W.A.; Jucks, K.W. The Spectrum of Earthshine: A Pale Blue Dot Observed from the Ground. Astrophys. J. 2002, 574, 430–433. [Google Scholar] [CrossRef] [Green Version]
- Seager, S.; Turner, E.L.; Schafer, J.; Ford, E.B. Vegetation’s red edge: A possible spectroscopic biosignature of extraterrestrial plants. Astrobiology 2005, 5. [Google Scholar] [CrossRef] [PubMed]
- Montanes-Rodriguez, P.; Palle, E.; Goode, P.R.; Martin-Torres, F.J. Vegetation Signature in the Observed Globally Integrated Spectrum of Earth Considering Simultaneous Cloud Data: Applications for Extrasolar Planets. Astrophys. J. 2006, 651, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, M.C.; Traub, W.A.; Jucks, K.W.; Woolf, N.J.; Meyer, M.R.; Gorlova, N.; Skrutskie, M.F.; Wilson, J.C. Spectrum of a Habitable World: Earthshine in the Near-Infrared. Astrophys. J. 2006, 644, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Kiang, N.Y.; Segura, A.; Tinetti, G.; Govindjee; Blankenship, R.E.; Cohen, M.; Siefert, J.; Crisp, D.; Meadows, V.S. Spectral Signatures of Photosynthesis. II. Coevolution with Other Stars And The Atmosphere on Extrasolar Worlds. Astrobiology 2007, 7, 252–274. [Google Scholar] [CrossRef] [Green Version]
- Hegde, S.; Paulino-Lima, I.G.; Kent, R.; Kaltenegger, L.; Rothschild, L. Surface biosignatures of exo-Earths: Remote detection of extraterrestrial life. Proc. Natl. Acad. Sci. USA 2015, 112, 3886–3891. [Google Scholar] [CrossRef] [Green Version]
- Kempton, E.M.; Bean, J.L.; Louie, D.R.; Deming, D.; Koll, D.D.B.; Mansfield, M.; Christiansen, J.L.; López-Morales, M.; Swain, M.R.; Zellem, R.T.; et al. A framework for prioritizing the TESS planetary candidates most amenable to atmospheric characterization. Publ. Astron. Soc. Pac. 2018, 130. [Google Scholar] [CrossRef] [Green Version]
- Seager, S. Exoplanet Atmospheres: Physical Processes; Princeton University Press: Princeton, NJ, USA, 2010; ISBN 9780691130262. [Google Scholar]
- Weiss, L.M.; Marcy, G.W. The Mass-Radius Relation for 65 Exoplanets Smaller than 4 Earth Radii. Astrophys. J. 2014, 783, L6. [Google Scholar] [CrossRef]
- Chen, J.; Kipping, D. Probabilistic Forecasting of the Masses and Radii of Other Worlds. Astrophys. J. 2017, 834, 17. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seager, S.; Petkowski, J.J.; Günther, M.N.; Bains, W.; Mikal-Evans, T.; Deming, D. Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres. Universe 2021, 7, 172. https://doi.org/10.3390/universe7060172
Seager S, Petkowski JJ, Günther MN, Bains W, Mikal-Evans T, Deming D. Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres. Universe. 2021; 7(6):172. https://doi.org/10.3390/universe7060172
Chicago/Turabian StyleSeager, Sara, Janusz J. Petkowski, Maximilian N. Günther, William Bains, Thomas Mikal-Evans, and Drake Deming. 2021. "Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres" Universe 7, no. 6: 172. https://doi.org/10.3390/universe7060172
APA StyleSeager, S., Petkowski, J. J., Günther, M. N., Bains, W., Mikal-Evans, T., & Deming, D. (2021). Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres. Universe, 7(6), 172. https://doi.org/10.3390/universe7060172