Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
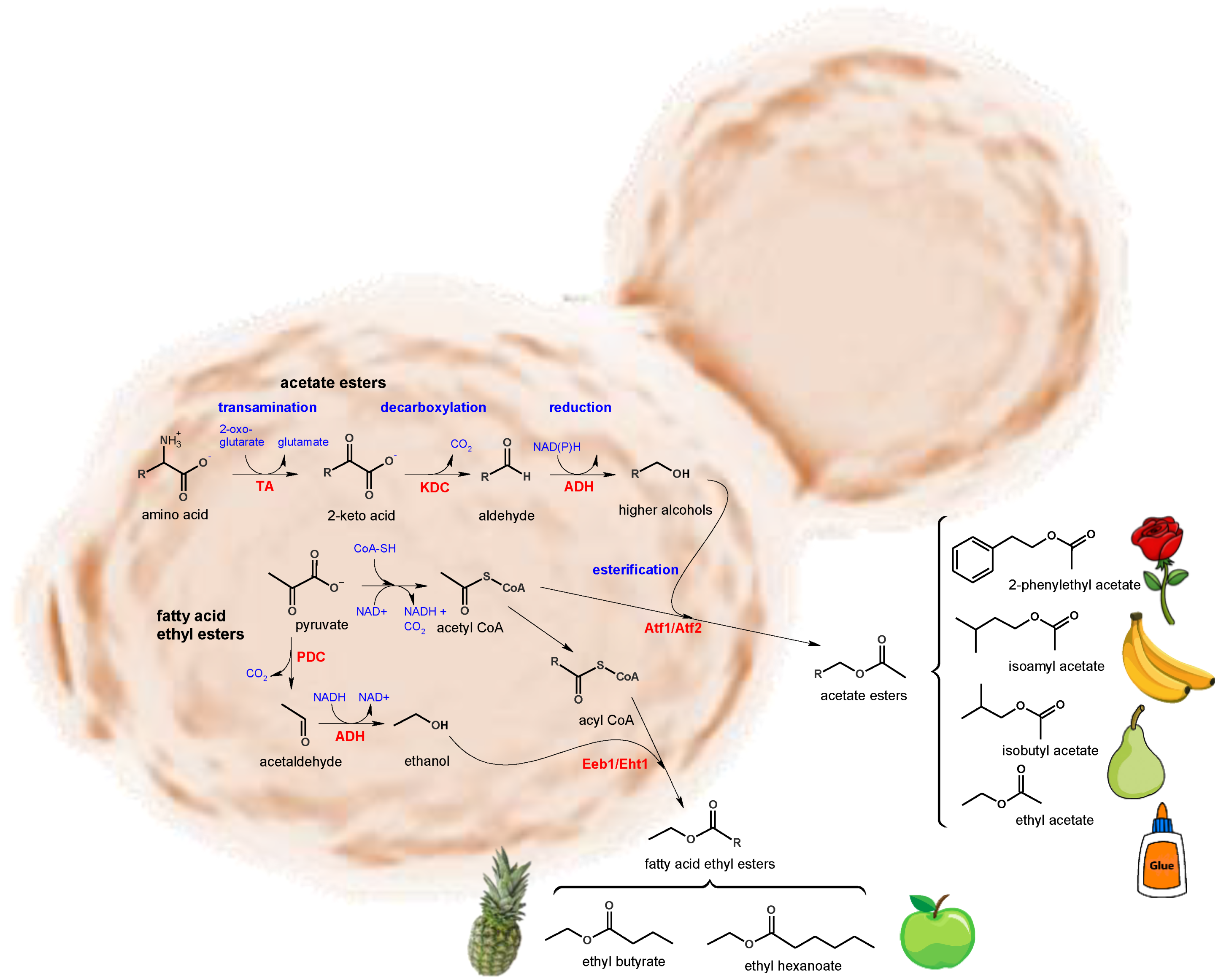
2. Influence of Non-Saccharomyces Yeasts on Wine Aroma
3. Acidity and pH Control
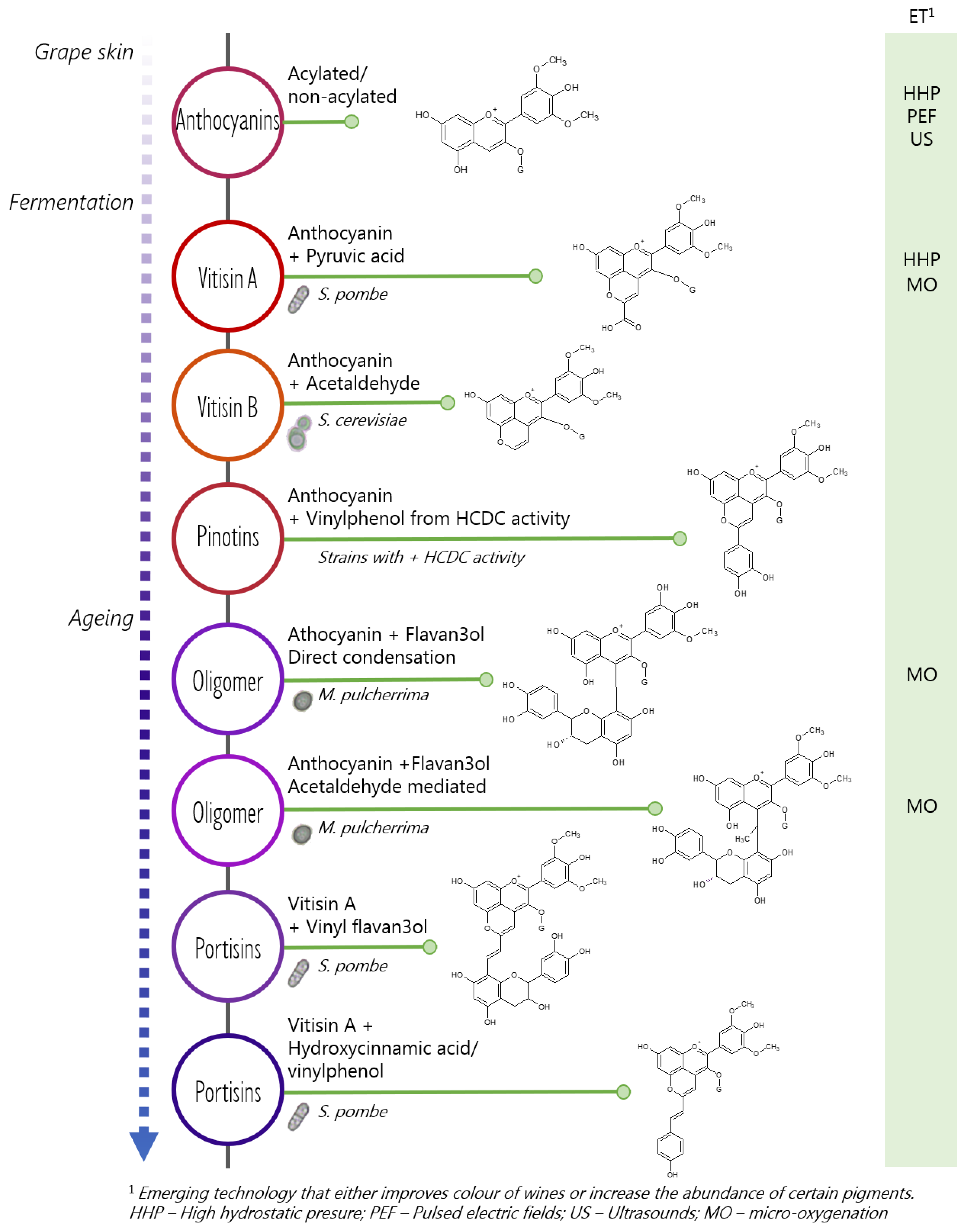
4. Non-Saccharomyces-Mediated Formation of Stable Pyranoanthocyanin and Polymeric Pigments
5. Commercially Available Non-Saccharomyces Yeasts
6. Non-Saccharomyces and Off-Flavour Production
7. Biological Control of Indigenous Yeasts Producing Defective Off-Flavours
8. Emerging Technologies to Improve the Implantation of Non-Saccharomyces
9. Cell Wall Polysaccharides from Non-Saccharomyces Yeasts
10. Conclusions
Funding
Conflicts of Interest
References
- Morata, A.; Loira, I.; Del Fresno, J.M.; Escott, C.; Bañuelos, M.A.; Tesfaye, W.; González, C.; Palomero, F.; Suárez-Lepe, J.A. Strategies to Improve the Freshness in Wines from Warm Areas. In Advances in Grape and Wine Biotechnology; Morata, A., Iris, L., Eds.; InTech: London, UK, 2019. [Google Scholar]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Delfini, C.; Cocito, C.; Bonino, M.; Schellino, R.; Gaia, P.; Baiocchi, C. Definitive evidence for the actual contribution of yeast in the transformation of neutral precursors of grape aromas. J. Agric. Food Chem. 2001, 49, 5397–5408. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R. Wine Science: Principles and Applications, 3rd ed.; Academic Press: London, UK, 2008; p. 751. [Google Scholar]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.; Velázquez, R. The Yeast Torulaspora delbrueckii: An Interesting But Difficult-To-Use Tool for Winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Gil, J.; Manzanares, P. Challenges of the Non-Conventional Yeast Wickerhamomyces anomalus in Winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.; Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.; Arroyo, T. Advances in the Study of Candida stellata. Fermentation 2018, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Rosi, I.; Vinella, M.; Domizio, P. Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 1994, 77, 519–527. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Zelle, R.M.; De Hulster, E.; Van Winden, W.A.; De Waard, P.; Dijkema, C.; Winkler, A.A.; Geertman, J.M.A.; Van Dijken, J.P.; Pronk, J.T.; Van Maris, A.J.A. Malic acid production by Saccharomyces cerevisiae: Engineering of pyruvate carboxylation, oxaloacetate reduction, and malate export. Appl. Environ. Microbiol. 2008, 74, 2766–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata, A.; Bañuelos, M.A.; Vaquero, C.; Loira, I.; Cuerda, R.; Palomero, F.; González, C.; Suárez-Lepe, J.A.; Wang, J.; Han, S.; et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur. Food Res. Technol. 2019, 245, 885–894. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Suárez Lepe, J.A. Influence of Yeasts in Wine Colour. In Grape and Wine Biotechnology; Morata, A., Ed.; InTech: London, UK, 2016; pp. 285–305. [Google Scholar]
- Morata, A.; Gómez-Cordovés, M.C.; Colomo, B.; Suárez, J.A. Pyruvic Acid and Acetaldehyde Production by Different Strains of Sacchromyces cerevisiae: Relationship with Vitisin A and B Formation in Red Wines. J. Agric. Food Chem. 2003, 51, 7402–7409. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Gómez-Cordovés, M.C.; Calderón, F.; Suárez-Lepe, J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006, 106, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; Morata, A.; Loira, I.; Tesfaye, W.; Suarez-Lepe, J.A. Characterization of polymeric pigments and pyranoanthocyanins formed in microfermentations of non-Saccharomyces yeasts. J. Appl. Microbiol. 2016, 121, 1346–1356. [Google Scholar] [CrossRef]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Tesfaye, W.; González, C.; Suárez-Lepe, J.A. Formation of polymeric pigments in red wines through sequential fermentation of flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018, 239, 975–983. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Suberviola, J.; Bartolomé, B.; Colomo, B.; Suárez, J.A. Adsorption of Anthocyanins by Yeast Cell Walls during the Fermentation of Red Wines. J. Agric. Food Chem. 2003, 51, 4084–4088. [Google Scholar] [CrossRef]
- Caridi, A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007, 120, 167–172. [Google Scholar] [CrossRef]
- Echeverrigaray, S.; Scariot, F.J.; Menegotto, M.; Delamare, A.P.L. Anthocyanin adsorption by Saccharomyces cerevisiae during wine fermentation is associated to the loss of yeast cell wall/membrane integrity. Int. J. Food Microbiol. 2020, 314, 108383. [Google Scholar] [CrossRef]
- Loira, I.; Vejarano, R.; Morata, A.; Ricardo-Da-Silva, J.M.; Laureano, O.; González, M.C.; Suárez-Lepe, J.A. Effect of Saccharomyces strains on the quality of red wines aged on lees. Food Chem. 2013, 139, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar] [CrossRef]
- Kulkarni, P.; Loira, I.; Morata, A.; Tesfaye, W.; González, M.C.; Suárez-Lepe, J.A. Use of Non Saccharomyces yeast strains coupled with ultrasound treatments as a novel technique to accelerate aging over lees of red wines and its repercussion in sensorial parameters. LWT - Food Sci. Technol. 2015, 64, 1255–1262. [Google Scholar] [CrossRef]
- García Martín, J.F.; Guillemet, L.; Feng, C.; Sun, D.W. Cell viability and proteins release during ultrasound-assisted yeast lysis of light lees in model wine. Food Chem. 2013, 141, 934–939. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Loira, I.; Morata, A.; González, C.; Suárez-Lepe, J.A.; Cuerda, R. Application of ultrasound to improve lees ageing processes in red wines. Food Chem. 2018, 261, 157–163. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Morata, A.; Escott, C.; Loira, I.; Cuerda, R.; Suárez-Lepe, J.A. Sonication of yeast biomasses to improve the ageing on lees technique in red wines. Molecules 2019, 24, 635. [Google Scholar] [CrossRef] [Green Version]
- Comuzzo, P.; Calligaris, S. Potential Applications of High Pressure Homogenization in Winemaking: A Review. Beverages 2019, 5, 56. [Google Scholar] [CrossRef] [Green Version]
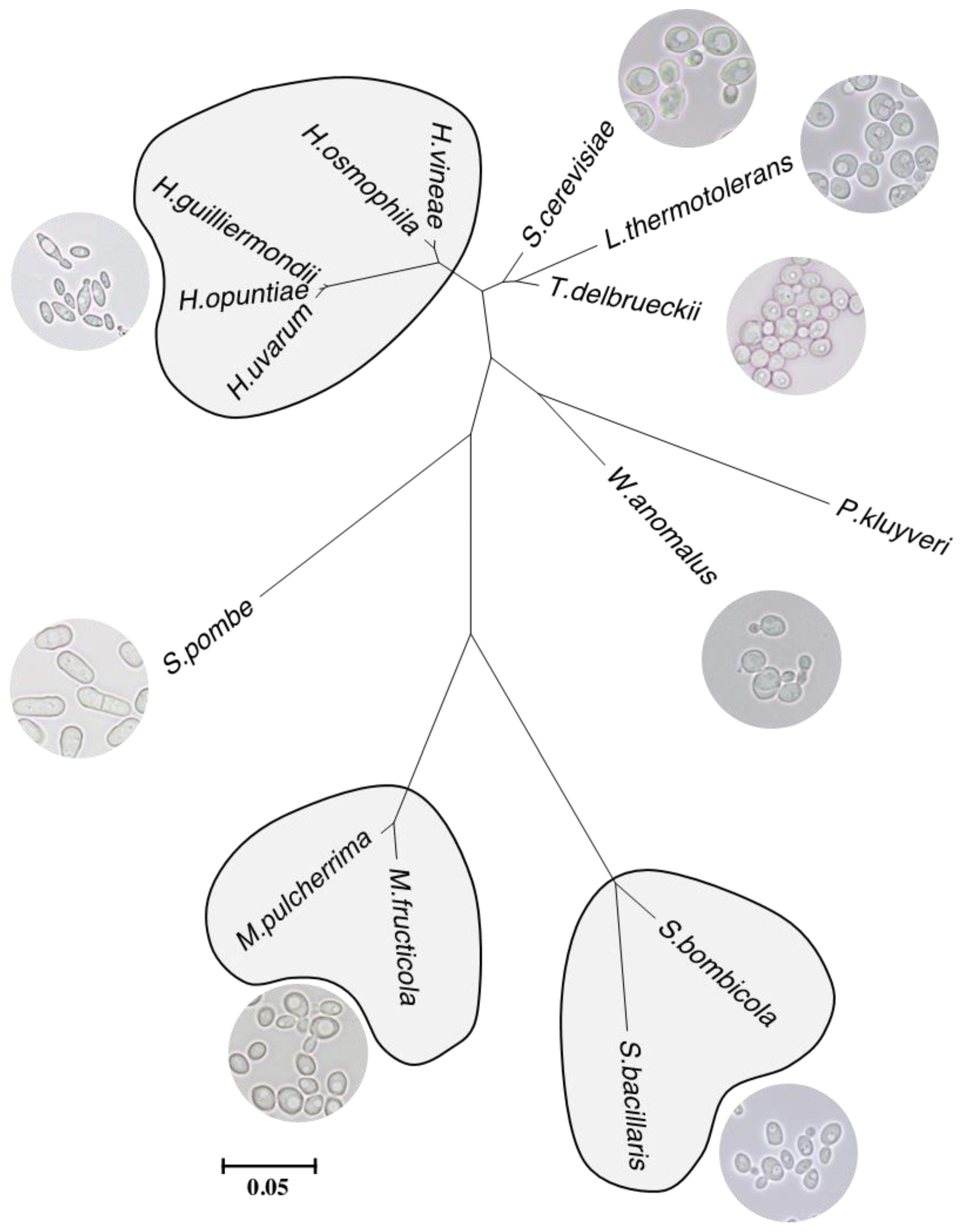
- Kurtzman, C.P.; Robnett, C.J. Phylogenetic relationships among yeasts of the “Saccharomyces complex” determined from multigene sequence analyses. FEMS Yeast Res. 2003, 3, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Vejarano, R.; González, C.; Callejo, M.J.; Suárez-Lepe, J.A. Emerging preservation technologies in grapes for winemaking. Trends Food Sci. Technol. 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
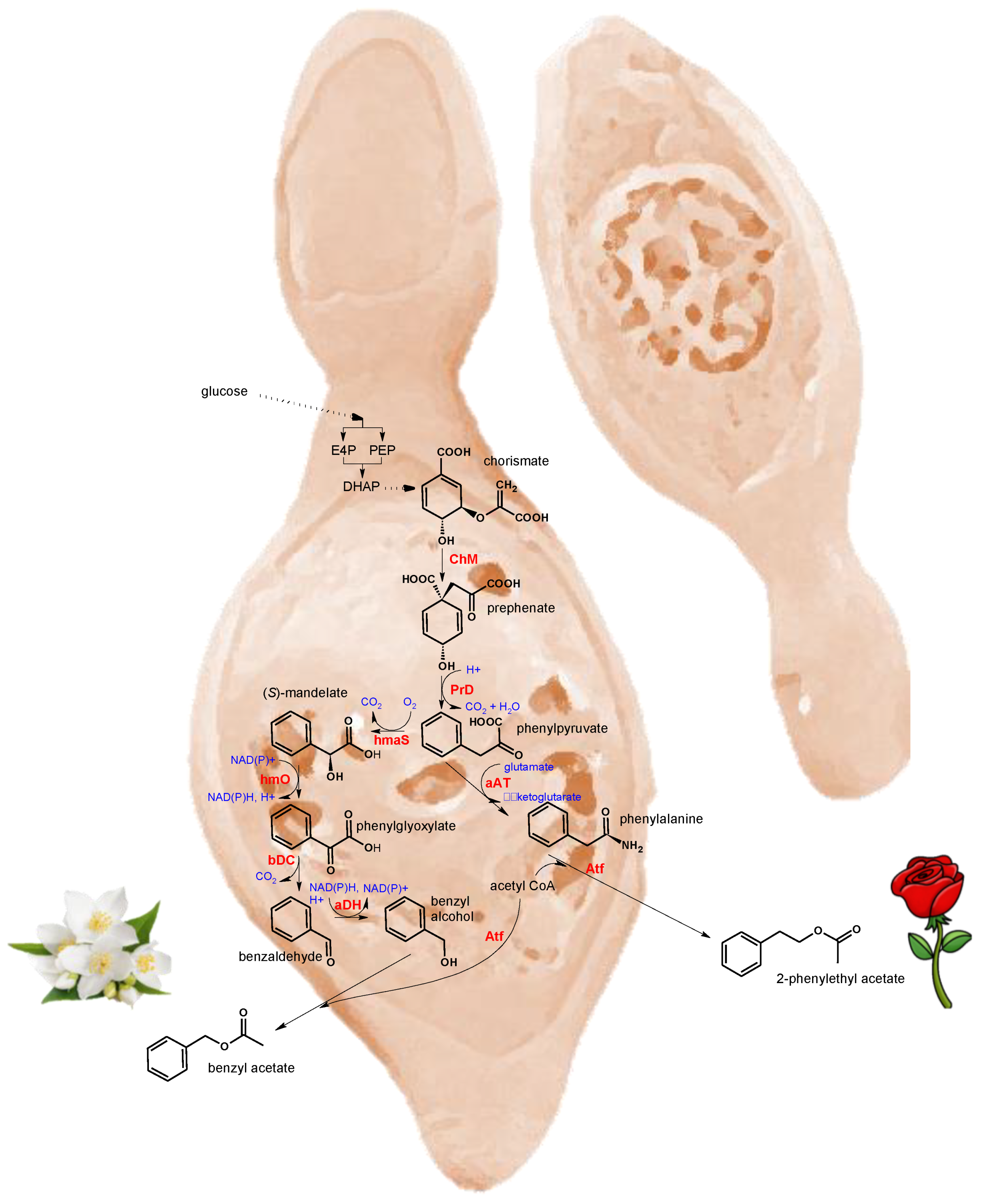
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Fariña, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and Transcriptomic Basis of Hanseniaspora vineae’s Impact on Flavor Diversity and Wine Quality. Appl. E 2019, 85, e01959-18. [Google Scholar]
- Martin, V.; Giorello, F.; Fariña, L.; Minteguiaga, M.; Salzman, V.; Boido, E.; Aguilar, P.S.; Gaggero, C.; Dellacassa, E.; Mas, A.; et al. De novo synthesis of benzenoid compounds by the yeast Hanseniaspora vineae increases the flavor diversity of wines. J. Agric. Food Chem. 2016, 64, 4574–4583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, V.; Boido, E.; Giorello, F.; Mas, A.; Dellacassa, E.; Carrau, F. Effect of yeast assimilable nitrogen on the synthesis of phenolic aroma compounds by Hanseniaspora vineae strains. Yeast 2016, 33, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotton, R.G.H.; Gibson, F. The biosynthesis of phenylalanine and tyrosine; enzymes converting chorismic acid into prephenic acid and their relationships to prephenate dehydratase and prephenate dehydrogenase. Biochim. Biophys. Acta Gen. Subj. 1965, 100, 76–88. [Google Scholar] [CrossRef]
- Pugh, S.; McKenna, R.; Halloum, I.; Nielsen, D.R. Engineering Escherichia coli for renewable benzyl alcohol production. Metab. Eng. Commun. 2015, 2, 39–45. [Google Scholar] [CrossRef]
- Marcon, A.R.; Schwarz, L.V.; Dutra, S.V.; Moura, S.; Agostini, F.; Delamare, A.P.L.; Echeverrigaray, S. Contribution of a Brazilian Torulaspora delbrueckii isolate and a commercial Saccharomyces cerevisiae to the aroma profile and sensory characteristics of Moscato Branco wines. Aust. J. Grape Wine Res. 2018, 24, 461–468. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; Moine, V.; Thibon, C.; Bely, M. Enhanced 3-Sulfanylhexan-1-ol Production in Sequential Mixed Fermentation with Torulaspora delbrueckii/Saccharomyces cerevisiae Reveals a Situation of Synergistic Interaction between Two Industrial Strains. Front. Microbiol. 2016, 7, 293. [Google Scholar] [CrossRef] [Green Version]
- Culleré, L.; López, R.; Ferreira, V. The Instrumental Analysis of Aroma-Active Compounds for Explaining the Flavor of Red Wines. In Red Wine Technology; Morata, A., Ed.; Academic Press: London, UK, 2019; pp. 283–307. ISBN 978-0-12-814399-5. [Google Scholar]
- Engel, K.H.; Tressl, R. Identification of new sulfur-containing volatiles in yellow passionfruit (Passiflora edulis f. flavicarpa). J. Agric. Food Chem. 1991, 39, 2249–2252. [Google Scholar] [CrossRef]
- Loira, I.; Vejarano, R.; Bañuelos, M.A.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suárez-Lepe, J.A. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [Green Version]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.; Valles, S.; Giraudo, M.R.; Caballero, A. Selection and preliminary characterization of β-glycosidases producer Patagonian wild yeasts. Enzyme Microb. Technol. 2007, 41, 812–820. [Google Scholar] [CrossRef]
- Manzanares, P.; Ramón, D.; Querol, A. Screening of non-Saccharomyces wine yeasts for the production of β-D- xylosidase activity. Int. J. Food Microbiol. 1999, 46, 105–112. [Google Scholar] [CrossRef]
- Spencer, J.F.T.; Gorin, P.A.J. Mannose-containing polysaccharides of the apiculate yeasts Nadsonia, Hanseniaspora, Kloeckera, and Sacharomycodes, and their use as an aid in classification. J. Bacteriol. 1968, 96, 180–183. [Google Scholar] [PubMed]
- Zhang, S.; Petersen, M.A.; Liu, J.; Toldam-Andersen, T.B. Influence of pre-fermentation treatments on wine volatile and sensory profile of the new disease tolerant cultivar solaris. Molecules 2015, 20, 21609–21625. [Google Scholar] [CrossRef] [Green Version]
- Banilas, G.; Sgouros, G.; Nisiotou, A. Development of microsatellite markers for Lachancea thermotolerans typing and population structure of wine-associated isolates. Microbiol. Res. 2016, 193, 1–10. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Hranilovic, A.; Bely, M.; Masneuf-Pomarede, I.; Jiranek, V.; Albertin, W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE 2017, 12, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812–14825. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
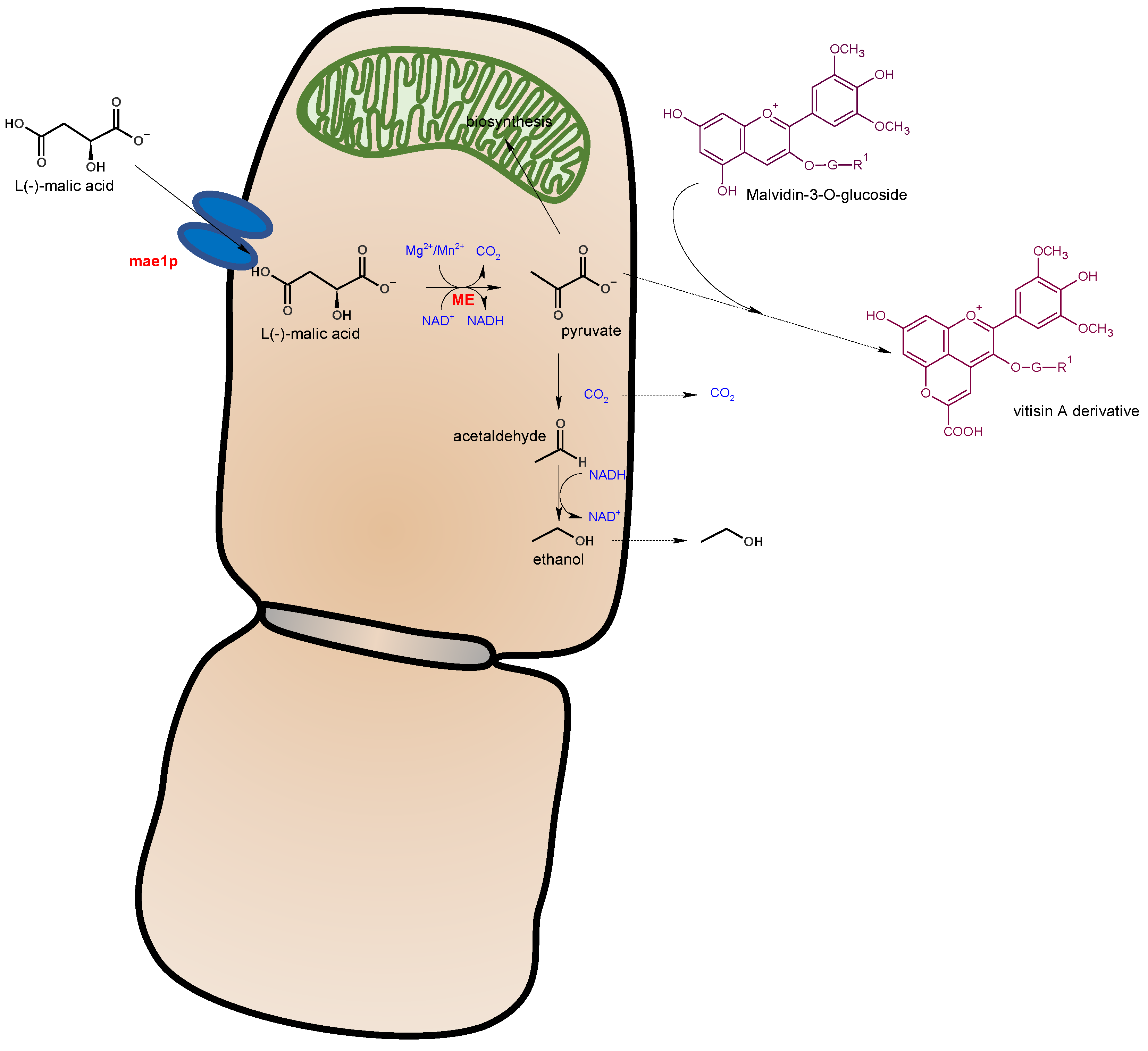
- Suárez-Lepe, J.A.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Volschenk, H.; van Vuuren, H.J.J.; Viljoen-Bloom, M. Malo-ethanolic fermentation in Saccharomyces and Schizosaccharomyces. Curr. Genet. 2003, 43, 379–391. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Palomero, F.; González, C.; Suárez-Lepe, J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation 2018, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Calderón, F.; González, M.C.; Gómez-Cordovés, M.C.; Suárez, J.A. Formation of the highly stable pyranoanthocyanins (vitisins A and B) in red wines by the addition of pyruvic acid and acetaldehyde. Food Chem. 2007, 100, 1144–1152. [Google Scholar] [CrossRef]
- Rentzsch, M.; Schwarz, M.; Winterhalter, P. Pyranoanthocyanins—An overview on structures, occurrence, and pathways of formation. Trends Food Sci. Technol. 2007, 18, 526–534. [Google Scholar] [CrossRef]
- Eglinton, J.; Griesser, M.; Henschke, P.; Kwiatkowski, M.; Parker, M.; Herderich, M. Yeast-Mediated Formation of Pigmented Polymers in Red Wine. Red Wine Color 2004, 886, 7–21. [Google Scholar]
- Es-Safi, N.E.; Fulcrand, H.; Cheynier, V.; Moutounet, M. Competition between (+)-catechin and (-)-epicatechin in acetaldehyde- induced polymerization of flavanols. J. Agric. Food Chem. 1999, 47, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Es-Safi, N.E.; Fulcrand, H.; Cheynier, V.; Moutounet, M. Studies on the acetaldehyde-induced condensation of (-)-epicatechin and malvidin 3-O-glucoside in a model solution system. J. Agric. Food Chem. 1999, 47, 2096–2102. [Google Scholar] [CrossRef] [PubMed]
- Dallas, C.; Ricardo-da-Silva, J.M.; Laureano, O. Products Formed in Model Wine Solutions Involving Anthocyanins, Procyanidin B2, and Acetaldehyde. J. Agric. Food Chem. 1996, 44, 2402–2407. [Google Scholar] [CrossRef]
- Quaglieri, C.; Jourdes, M.; Waffo-Teguo, P.; Teissedre, P.L. Updated knowledge about pyranoanthocyanins: Impact of oxygen on their contents, and contribution in the winemaking process to overall wine color. Trends Food Sci. Technol. 2017, 67, 139–149. [Google Scholar] [CrossRef]
- Mateus, N.; Oliveira, J.; Pissarra, J.; González-Paramás, A.M.; Rivas-Gonzalo, J.C.; Santos-Buelga, C.; Silva, A.M.S.; De Freitas, V. A new vinylpyranoanthocyanin pigment occurring in aged red wine. Food Chem. 2006, 97, 689–695. [Google Scholar] [CrossRef]
- Mateus, N.; Oliveira, J.; Haettich-Motta, M.; de Freitas, V. New family of bluish pyranoanthocyanins. J. Biomed. Biotechnol. 2004, 5, 299–305. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Liang, N.N.; Mu, L.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Anthocyanins and their variation in red wines II. Anthocyanin derived pigments and their color evolution. Molecules 2012, 17, 1483–1519. [Google Scholar] [CrossRef] [Green Version]
- Bañuelos, M.A.; Loira, I.; Escott, C.; Del Fresno, J.M.; Morata, A.; Sanz, P.D.; Otero, L.; Suárez-Lepe, J.A. Grape Processing by High Hydrostatic Pressure: Effect on Use of Non-Saccharomyces in Must Fermentation. Food Bioprocess Technol. 2016, 9, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Fava, J.; Hodara, K.; Nieto, A.; Guerrero, S.; Alzamora, S.M.; Castro, M.A. Structure (micro, ultra, nano), color and mechanical properties of Vitis labrusca L. (grape berry) fruits treated by hydrogen peroxide, UV-C irradiation and ultrasound. Food Res. Int. 2011, 44, 2938–2948. [Google Scholar] [CrossRef]
- Puértolas, E.; López, N.; Saldaña, G.; Álvarez, I.; Raso, J. Evaluation of phenolic extraction during fermentation of red grapes treated by a continuous pulsed electric fields process at pilot-plant scale. J. Food Eng. 2010, 98, 120–125. [Google Scholar] [CrossRef]
- Cano-López, M.; López-Roca, J.M.; Pardo-Minguez, F.; Gómez Plaza, E. Oak barrel maturation vs. micro-oxygenation: Effect on the formation of anthocyanin-derived pigments and wine colour. Food Chem. 2010, 119, 191–195. [Google Scholar] [CrossRef]
- Morata, A.; López, C.; Tesfaye, W.; González, C.; Escott, C. Anthocyanins as natural pigments in beverages. In Value-added ingredients and enrichements of beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: London, UK, 2019; pp. 383–428. ISBN 9780128166871. [Google Scholar]
- Rodriguez-Amaya, D.B. Update on natural food pigments—A mini-review on carotenoids, anthocyanins, and betalains. Food Res. Int. 2018, 124, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.R.C.; Murador, D.C.; de Souza Mesquita, L.M.; de Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Influence of Saccharomyces and non-Saccharomyces Yeasts in the Formation of Pyranoanthocyanins and Polymeric Pigments during Red Wine Making. Molecules 2019, 24, 4490. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Suárez-Lepe, J.A. New Biotechnologies for Wine Fermentation and Ageing. In Advances in Food Biotechnology; V, R.R., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2016; pp. 287–302. ISBN 9781118864555. [Google Scholar]
- Siebert, T.E.; Solomon, M.R.; Pollnitz, A.P.; Jeffery, D.W. Selective determination of volatile sulfur compounds in wine by gas chromatography with sulfur chemiluminescence detection. J. Agric. Food Chem. 2010, 58, 9454–9462. [Google Scholar] [CrossRef]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from Douro Wine Region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Sabel, A.; Martens, S.; Petri, A.; König, H.; Claus, H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2014, 64, 483–491. [Google Scholar] [CrossRef]
- Fan, G.; Teng, C.; Xu, D.; Fu, Z.; Minhazul, K.A.H.M.; Wu, Q.; Liu, P.; Yang, R.; Li, X. Enhanced production of ethyl acetate using co-culture of Wickerhamomyces anomalus and Saccharomyces cerevisiae. J. Biosci. Bioeng. 2019, 128, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Orentaite, I.; Poranen, M.M.; Oksanen, H.M.; Daugelavicius, R.; Bamford, D.H. K2 killer toxin-induced physiological changes in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Xiao, H.; Cheng, X.; Zhou, H.; Si, L. Hanseniaspora uvarum prolongs shelf life of strawberry via volatile production. Food Microbiol. 2017, 63, 205–212. [Google Scholar] [CrossRef]
- Guo, H.; Qin, X.; Wu, Y.; Yu, W.; Liu, J.; Xi, Y.; Dou, G.; Wang, L.; Xiao, H. Biocontrol of Gray Mold of Cherry Tomatoes with the Volatile Organic Monomer from Hanseniaspora uvarum, Trans-Cinnamaldehyde. Food Bioprocess Technol. 2019, 12, 1809–1820. [Google Scholar] [CrossRef]
- Passoth, V.; Fredlund, E.; Druvefors, U.Ä.; Schnürer, J. Biotechnology, physiology and genetics of the yeast Pichia anomala. FEMS Yeast Res. 2006, 6, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Csutak, O.; Vassu, T.; Sarbu, I.; Stoica, I.; Cornea, P. Antagonistic activity of three newly isolated yeast strains from the surface of fruits. Food Technol. Biotechnol. 2013, 51, 70–77. [Google Scholar]
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol potential of Metchnikowia pulcherrima strains against blue mold of apple. Phytopathology 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Kántor, A.; Hutková, J.; Petrová, J.; Hleba, L.; Kačániová, M. Antimicrobial activity of pulcherrimin pigment produced by Metschnikowia pulcherrima against various yeast species. J. Microbiol. Biotechnol. Food Sci. 2015, 5, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Ingeniis De, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata, A.; Loira, I.; Vejarano, R.; Bañuelos, M.A.; Sanz, P.D.; Otero, L.; Suárez-Lepe, J.A. Grape Processing by High Hydrostatic Pressure: Effect on Microbial Populations, Phenol Extraction and Wine Quality. Food Bioprocess Technol. 2015, 8, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Corrales, M.; Toepfl, S.; Butz, P.; Knorr, D.; Tauscher, B. Extraction of anthocyanins from grape by-products assisted by ultrasonics, high hydrostatic pressure or pulsed electric fields: A comparison. Innov. Food Sci. Emerg. Technol. 2008, 9, 85–91. [Google Scholar] [CrossRef]
- Corrales, M.; García, A.F.; Butz, P.; Tauscher, B. Extraction of anthocyanins from grape skins assisted by high hydrostatic pressure. J. Food Eng. 2009, 90, 415–421. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Bañuelos, M.A.; Puig-Pujol, A.; Guamis, B.; González, C.; Suárez-Lepe, J.A. Use of Ultra-High Pressure Homogenization processing in winemaking: Control of microbial populations in grape musts and effects in sensory quality. Innov. Food Sci. Emerg. Technol. 2018, 50, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Saldaña, G.; Álvarez, I.; Condón, S.; Raso, J. Microbiological aspects related to the feasibility of PEF technology for food pasteurization. Crit. Rev. Food Sci. Nutr. 2014, 54, 1415–1426. [Google Scholar] [CrossRef]
- López, N.; Puértolas, E.; Condón, S.; Álvarez, I.; Raso, J. Effects of pulsed electric fields on the extraction of phenolic compounds during the fermentation of must of Tempranillo grapes. Innov. Food Sci. Emerg. Technol. 2008, 9, 477–482. [Google Scholar] [CrossRef]
- Puértolas, E.; López, N.; Condón, S.; Álvarez, I.; Raso, J. Potential applications of PEF to improve red wine quality. Trends Food Sci. Technol. 2010, 21, 247–255. [Google Scholar] [CrossRef]
- Puértolas, E.; Saldaña, G.; Álvarez, I.; Raso, J. Experimental design approach for the evaluation of anthocyanin content of rosé wines obtained by pulsed electric fields. Influence of temperature and time of maceration. Food Chem. 2011, 126, 1482–1487. [Google Scholar] [CrossRef]
- Wu, Y.; Mittal, G.S.; Griffiths, M.W. Effect of pulsed electric field on the inactivation of microorganisms in grape juices with and without antimicrobials. Biosyst. Eng. 2005, 90, 1–7. [Google Scholar] [CrossRef]
- Escott, C.; Vaquero, C.; del Fresno, J.M.; Bañuelos, M.A.; Loira, I.; Han, S.Y.; Bi, Y.; Morata, A.; Suárez-Lepe, J.A. Pulsed Light Effect in Red Grape Quality and Fermentation. Food Bioprocess Technol. 2017, 10, 1540–1547. [Google Scholar] [CrossRef]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine—A review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; González, M.C.; Suárez-Lepe, J.A. Conventional and enzyme-assisted autolysis during ageing over lees in red wines: Influence on the release of polysaccharides from yeast cell walls and on wine monomeric anthocyanin content. Food Chem. 2007, 105, 838–846. [Google Scholar] [CrossRef]
- Liu, L.; Loira, I.; Morata, A.; Suárez-Lepe, J.A.; González, M.C.; Rauhut, D. Shortening the ageing on lees process in wines by using ultrasound and microwave treatments both combined with stirring and abrasion techniques. Eur. Food Res. Technol. 2016, 242, 559–569. [Google Scholar] [CrossRef]
- Maza, M.A.; Delso, C.; Álvarez, I.; Raso, J.; Martínez, J.M. Effect of pulsed electric fields on mannoproteins release from Saccharomyces cerevisiae during the aging on lees of Caladoc red wine. LWT 2020, 118, 108788. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Maza, M.A.; Álvarez, I.; Raso, J. Pulsed electric fields accelerate release of mannoproteins from Saccharomyces cerevisiae during aging on the lees of Chardonnay wine. Food Res. Int. 2019, 116, 795–801. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Aguilar, D.; Cebrián, G.; Álvarez, I.; Raso, J. Factors influencing autolysis of Saccharomyces cerevisiae cells induced by pulsed electric fields. Food Microbiol. 2018, 73, 67–72. [Google Scholar] [CrossRef]
- Morata, A.; Calderón, F.; González, M.C.; Colomo, B.; Suárez, J.A. Crianza sobre lías, chips y microoxigenación, utilización conjunta en el envejecimiento de vinos tintos. Ventajas del uso de levaduras seleccionadas. Enólogos. 2005, 34, 1–8. [Google Scholar]
- Gallart, M.; López-Tamames, E.; Suberbiola, G.; Buxaderas, S. Influence of Fatty Acids on Wine Foaming. J. Agric. Food Chem. 2002, 50, 7042–7045. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, S.; Charpentier, C.; Feuillat, M.; Voilley, A. Influence of yeast walls on the behavior of aroma compounds in a model wine. Am. J. Enol. Vitic. 1994, 45, 29–33. [Google Scholar]
- Del Barrio-Galán, R.; Pérez-Magariño, S.; Ortega-Heras, M. Effect of the aging on lees and other alternative techniques on the low molecular weight phenols of Tempranillo red wine aged in oak barrels. Anal. Chim. Acta 2012, 732, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Saucier, C.; Little, D.; Glories, Y. First evidence of acetaldehyde-flavanol condensation products in red wine. Am. J. Enol. Vitic. 1997, 3, 370–373. [Google Scholar]
- Sartor, S.; Toaldo, I.M.; Panceri, C.P.; Caliari, V.; Luna, A.S.; de Gois, J.S.; Bordignon-Luiz, M.T. Changes in organic acids, polyphenolic and elemental composition of rosé sparkling wines treated with mannoproteins during over-lees aging. Food Res. Int. 2019, 124, 34–42. [Google Scholar] [CrossRef]
- González-Marco, A.; Ancín-Azpilicueta, C. Influence of Lees Contact on Evolution of Amines in Chardonnay Wine. J. Food Sci. 2006, 71, C544–C548. [Google Scholar] [CrossRef]
- Masino, F.; Montevecchi, G.; Arfelli, G.; Antonelli, A. Evaluation of the Combined Effects of Enzymatic Treatment and Aging on Lees on the Aroma of Wine from Bombino bianco Grapes. J. Agric. Food Chem. 2008, 56, 9495–9501. [Google Scholar] [CrossRef]
- Bueno, J.E.; Peinado, R.A.; Medina, M.; Moreno, J. Effect of a short contact time with lees on volatile composition of Airen and Macabeo wines. Biotechnol. Lett. 2006, 28, 1007–1011. [Google Scholar] [CrossRef]
- Dubourdieu, D.; Lavigne-Cruege, V. The Role of Glutathione on the Aromatic Evolution of Dry White Wine. Available online: https://www.infowine.com/en/technical_articles/the_role_of_glutathione_on_the_aromatic_evolution_of_dry_white_wine._sc_993.htm# (accessed on 7 December 2019).
- Liberatore, M.T.; Pati, S.; Del Nobile, M.A.; Notte, E. La Aroma quality improvement of Chardonnay white wine by fermentation and ageing in barrique on lees. Food Res. Int. 2010, 43, 996–1002. [Google Scholar] [CrossRef]
- Chalier, P.; Angot, B.; Delteil, D.; Doco, T.; Gunata, Z. Interactions between aroma compounds and whole mannoprotein isolated from Saccharomyces cerevisiae strains. Food Chem. 2007, 100, 22–30. [Google Scholar] [CrossRef]
- Giovani, G.; Rosi, I.; Bertuccioli, M. Quantification and characterization of cell wall polysaccharides released by non-Saccharomyces yeast strains during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 113–118. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Saccharomyces Species | Metabolite/ Biopolymer | Structure | Sensory Repercussion | Technical Impact | Sensory Olfactive Threshold (µg/L)1 * If usually above This Value | Reference |
|---|---|---|---|---|---|---|
| Hanseniaspora/Kloeckera | 2-Phenylethyl acetate |  | Floral, rose petals hints | Enhance floral notes x2-10 compared to S. cerevisiae | 250 * | [35,37] |
| Mannans | Cell wall polysaccharides, mannoproteins | Increased mouthfeel, even perceptible after fermentation | [50] | |||
| Hanseniaspora vineae | Benzyl acetate |  | Floral jasmine aroma | Floral | 2 * | [35,51] |
| Lachancea thermotolerans | 2-Phenylethyl acetate |  | Floral, rose petals hints | 10–50 mg/L | 250 * | [15] |
| Ethyl lactate |  | Strawberry, toffee | >40 mg/L High sensory threshold | 150,000 | [15] | |
| Lactic acid |  | Citric acidity | 0.3–16 g/L Up to 0.5 pH reductions in oenological conditions Slight sugar depletion with some alcohol reduction | [15,52] | ||
| Metschnikowia pulcherrima | 2-Phenylethanol |  | Rose-like odour | >30 mg/L | 14,000 * | [48] |
| Monoterpens (e.g., linalool) |  | Floral | Increase varietal aromas by hydrolysing glucoside terpenes | 25 * | [48] | |
| Pichia kluyveri | Mercaptohexanol (3-MH) |  | Grapefruit, passion fruit | Fruity smell: > 625 ng/L single fermentation to 3000 ng/L co-inoculation | 0.060 * | [46] |
| Mercaptohexyl acetate (3-MHA) |  | Grapefruit, passion fruit | Fruity smell: > 500 ng/L single fermentation to 1700 ng/L co-inoculation | 0.004 * | [46] | |
| Schizosaccharomyces pombe | Pyruvate |  | Stable pigments, colour stability Precursor for vitisin A type compounds:  | Enhance the formation of vitisin A derivatives Some strains also vinylphenolic pyranoanthocyanins | [53] | |
| Cell wall polysaccharides, mannoproteins | Better wine structure, softening of the astringency | Increased mouthfeel | ||||
| Torulaspora delbrueckii | 2-Phenylethyl acetate |  | Flower, honey | 1.2-2x compared to S. cerevisiae & S. uvarum | 250 * | [42,44] |
| Ethyl hexanoate |  | Apple | Fruity smell | 62 * | [44] | |
| 3-Ethoxy-1-propanol |  | Black currant, solvent | Black fruity smell | [44] | ||
| Wickerhamomyces anomalus | 2-phenylethyl acetate |  | Flower, honey | 250 * | [42] | |
| Isoamyl acetate |  | Banana | Enhance fruitiness | 30 * | [42] | |
| Ethyl acetate |  | Fruity at low concentration | Fruity smell at low concentration Enhance complexity | 12,300 * | [54] |
| Non-Saccharomyces Species Commercially Available | Brand, Producer, Year, Format | Sensory Repercussion | Fermentative Performance (% vol. Ethanol) Recommended Inoculation Dose | Application. Requirements. | Reference |
|---|---|---|---|---|---|
| Hanseniaspora vineae | OENOBRANDS Launch 2021 Dry yeast | Enhance production of fruity and floral esters De novo formation of floral esters from sugars Increased body, softness and roundness | 10% | Low SO2 Nutrition: thiamine and yeast extract | http://www.oenobrands.com/en/our-innovation |
| Lachancea thermotolerans | CONCERTOTM CHr HANSEN 2012 Dry yeast | Red and black fruit integration Round mouthfeel Soft acidification Low volatile acidity, H2S and SO2 | 10% 25 g/HL | Red/White wines from warm areas | https://www.chr-hansen.com/ |
| LEVEL2 LAKTIA™ LALLEMAND 2018 Dry yeast | Enhancement of aromatic complexity, freshness and acidity Lactic acid production Low volatile acidity High glycerol production | <10% 25 g/HL | High nitrogen requirements Free SO2 must be <15 mg/L | https://www.lallemandwine.com/ | |
| Metschnikowia fructicola | Gaïa™ LALLEMAND 2016 Dry yeast | Improvement of the sensory expression Preservation of varietal character | Low to no fermentative power 7–25 g/HL | Reduce the use of SO2 and volatile acidity Active K2. Biological control Resistant to low pH and 50 mg/L SO2 | https://www.lallemandwine.com/ |
| Metschnikowia pulcherrima | LEVEL2 FLAVIA® MP346 LALLEMAND 2013 Dry yeast | Release of varietal aromas | 9% 25 g/HL | Specific enzymatic activity helping in releasing varietal aromas (terpenes and thiols) Free SO2 must be <15 mg/L | https://www.lallemandwine.com/ |
| LEVULIA® PULCHERRIMA AEB Dry yeast | High production of 2-phenyl and isoamyl acetates and terpenes Low volatile acidity | 11.5% 20–50 g/HL | Release of varietal aromas (terpenes) | https://www.aeb-group.com/ | |
| Pichia kluyveri | FROOTZEN® CHr HANSEN 2010 Frozen yeast | Enhancement of volatile thiols Blackcurrant, liquorice, black pepper, menthol aromas | 4–5% 1 bag/10kL | Enhance fruitiness | https://www.chr-hansen.com/ |
| Torulaspora delbrueckii | PRELUDETM CHr HANSEN 2009 Frozen yeast | Enhance production of medium chain fatty acid esters Increased body, softness and roundness | 9% 25 g/HL | Enhance fruitiness Promote malolactic fermentation by depletion of medium chain fatty acids | https://www.chr-hansen.com/ |
| LEVEL2 BIODIVA™ TD291 LALLEMAND 2009 Dry yeast | Aromatic complexity, ester production Low volatile acidity | 10% 25 g/HL | Tolerance to osmotic pressure. Adapted for fermenting late harvest and ice wines Free SO2 must be <15 mg/L | https://www.lallemandwine.com/ | |
| ZYMAFLORE® Alpha LAFFORT 2010 Dry yeast | Aromatic complexity and good mouthfeel Suitable for making expressive and full-bodied wines Revelation of thiol-type varietal aromas (3MH, 3MHA) | 10% 25 g/HL | Medium nitrogen requirements Low volatile acidity, volatile phenols and H2S | https://laffort.com/en/ | |
| Torulaspora delbrueckii + Saccharomyces spp. | Oenoferm® wild & pure F3 Erbslöh Dry yeast | Enhanced mono terpenes and formation of fruity esters Support the ripe and exotic fruit aroma Full-bodied wines | - 20–40 g/HL | Moderate to high nitrogen requirement High alcohol tolerance | https://erbsloeh.com/en/ |
| Torulaspora delbrueckii + Metschnikowia pulcherrima | ZYMAFLORE® ÉGIDE LAFFORT 2017 Dry yeast | Organoleptic neutrality and control the microflora | 10% 2–5 g/HL | Bioprotection of grapes and juices Restriction of the growth of indigenous flora SO2 reduction | https://laffort.com/en/ |
| Schizosaccharomyces pombe | ProMalic® PROENOL Dry beads | Wine deacidification | 100g/HL | Maloalcoholic fermentation Free SO2 must be <14 mg/L | https://www.proenol.com/ |
| Non-Saccharomyces species | Metabolite | Structure | Off-Smell/Off-Flavour | Technical Impact. Sensory Threshold (µg/L) 1 | Reference |
|---|---|---|---|---|---|
| Hanseniaspora/Kloeckera | Acetic acid |  | Vinegar taste | Many species/strains >0.6 g/L of volatile acidity 300,000 | [85] |
| Ethyl acetate |  | Solvent smell, nail varnish | Many species/strains >100 mg/L 12,300 | [85] | |
| Lachancea thermotolerans | Lactic acid |  | Some strains can produce excessive sour taste | Some strains >7 g/L; maximum described 16 g/L | [15,52] |
| Metschnikowia pulcherrima | Acetic acid |  | Vinegar taste | >0.7 g/L 300,000 | [86] |
| Ethyl acetate |  | Solvent smell, nail varnish | >200 mg/L 12,300 | [84] | |
| Hexanoic acid |  | Fatty, cheese | >1.1 mg/L 420 | [84] | |
| Octanoic acid |  | Rancid, harsh | >1.2 mg/L 500 | [84] | |
| Schizosaccharomyces pombe | Acetic acid |  | Vinegar taste | Many strains >1 g/L of volatile acidity 300,000 | [62,86] |
| Torulaspora delbrueckii | Sulphydric acid |  | Reductive smell, egg rot smell | Impact depending on the intensity 1.1 | [57,82] |
| Wickerhamomyces anomalus | Ethyl acetate |  | Solvent smell, nail varnish | ≈100 mg/L 12,300 | [85,87] |
| Acetic acid |  | Vinegar taste | 0.02 g/L 300,000 | [86,87,88] |
| Non-Saccharomyces Species | Metabolite | Antimicrobial Effect | Technical Impact | Reference |
|---|---|---|---|---|
| Hanseniaspora uvarum | trans-Cinnamaldehyde | Inhibition of mycelium growth and conidia germination | Biocontrol of Botrytis cinerea | [90,91] |
| Metschnikowia pulcherrima | Pulcherrimin | Iron depletion | Biological control Effective inhibitory activity against several yeasts: Candida tropicalis, Candida albicans, Brettanomyces/Dekkera, Hanseniaspora and Pichia genera; and some fungi: Botrytis cinerea, Penicillium spp., Alternaria spp. and Monilia spp. | [8,93,94,95,96,97,98] |
| Killer factor | Membrane permeabilization | Biological control | [57] | |
| Metschnikowia fructicola | Killer factor: active K2 | Increase of membrane permeability | Biological control of apiculate yeasts: K. apiculate/H. uvarum Reduction of volatile acidity | www.lallemandwine.com/ |
| Wickerhamomyces anomalus | Exo-β-1,3 glucanase | Wall-lytic enzymes | Inhibition of Botrytis cinerea | [92] |
| Pikt killer toxin | Mycocins that control apiculate wine yeasts | Biological control of Dekkera/Brettanomyces spp. | [92,99] | |
| Torulaspora delbrueckii | Kbarr-1 killer toxin | Toxicity against sensitive strains of S. cerevisiae | Better implantation and domination of the fermentation on the native S. cerevisiae of the must | [45] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules 2020, 10, 34. https://doi.org/10.3390/biom10010034
Morata A, Escott C, Bañuelos MA, Loira I, del Fresno JM, González C, Suárez-Lepe JA. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules. 2020; 10(1):34. https://doi.org/10.3390/biom10010034
Chicago/Turabian StyleMorata, Antonio, Carlos Escott, María Antonia Bañuelos, Iris Loira, Juan Manuel del Fresno, Carmen González, and José Antonio Suárez-Lepe. 2020. "Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review" Biomolecules 10, no. 1: 34. https://doi.org/10.3390/biom10010034
APA StyleMorata, A., Escott, C., Bañuelos, M. A., Loira, I., del Fresno, J. M., González, C., & Suárez-Lepe, J. A. (2020). Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules, 10(1), 34. https://doi.org/10.3390/biom10010034