Interaction between Polyphenolic Antioxidants and Saccharomyces cerevisiae Cells Defective in Heavy Metal Transport across the Plasma Membrane


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Growth Media
2.2. Yeast Strains and Storage
2.3. Growth Assessment of the Yeast Strains
2.4. Multielemental Analysis of Yeast Cells
2.5. Gene Expression Analysis by Quantitative Real Time-PCR (qRT-PCR)
2.6. Reproducibility of the Results and Statistics
3. Results
3.1. Effect of Polyphenolic Antioxidants on the Growth of Yeast Cells Defective in Heavy Metal Transporters
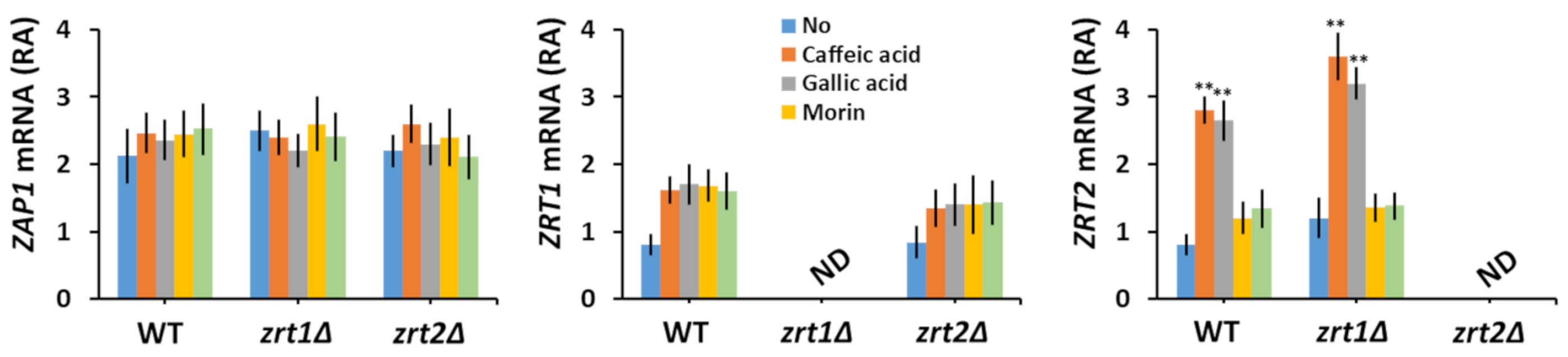
3.2. Polyphenols Alter the Heavy Metal Content of Yeast Cells
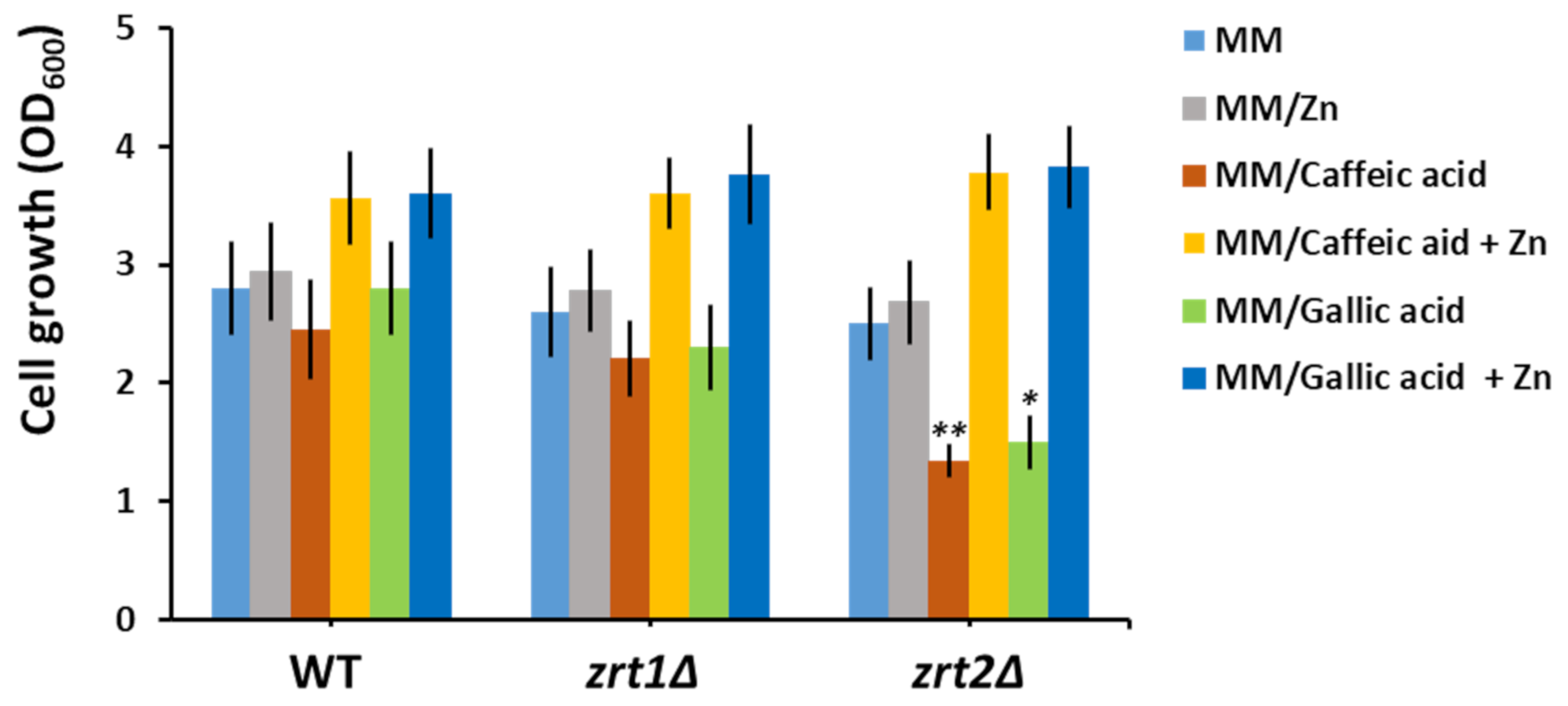
3.3. Supplementary Zn Is Stimulative of Yeast Cell Growth in the Presence of Caffeic and Gallic Acids
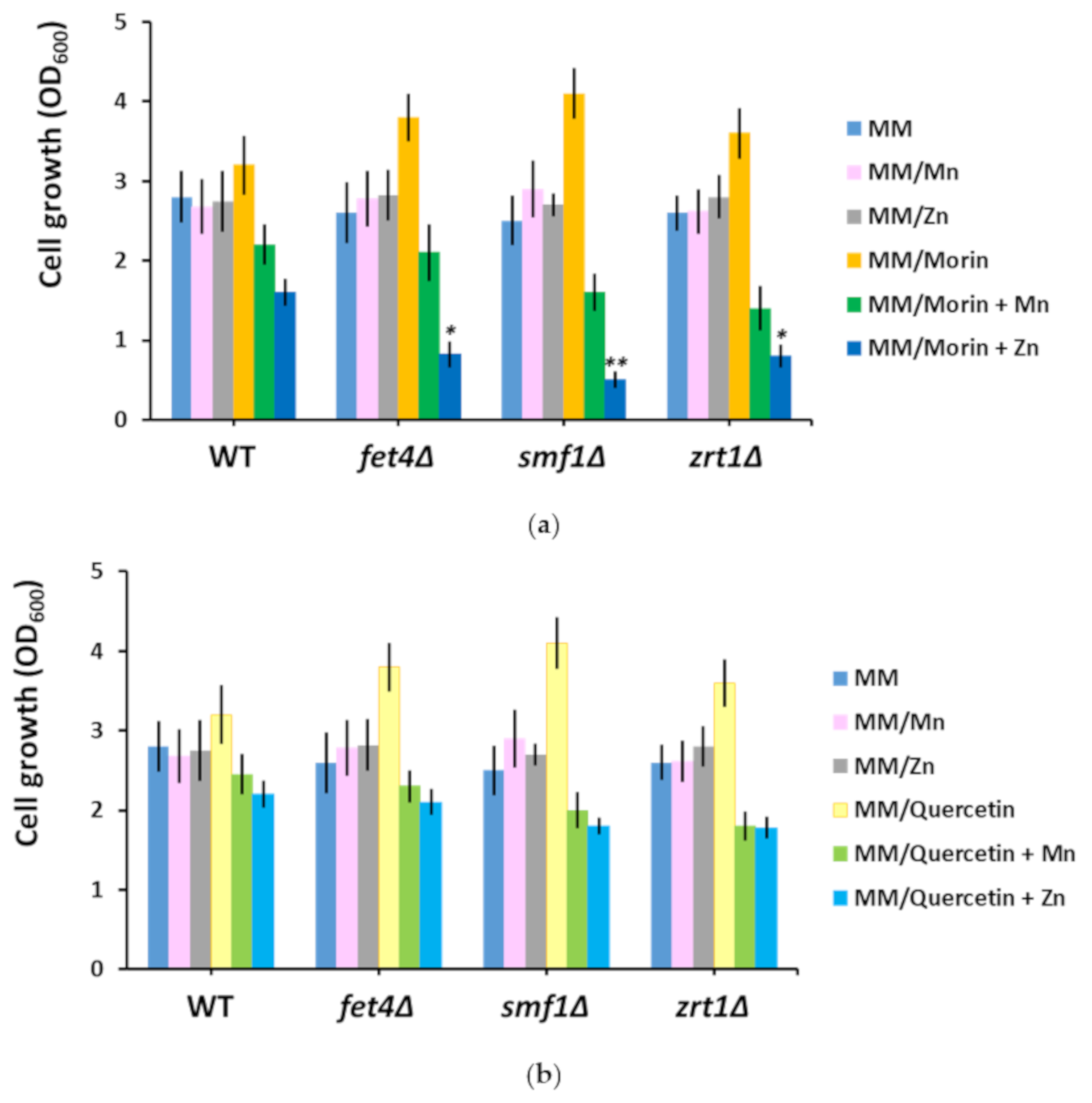
3.4. Supplementary Mn and Zn Abrogate the Chemoprotective Effect of Morin and Quercetin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, R.K.; Agrawal, M. Biological effects of heavy metals: An overview. J. Environ. Biol. 2005, 26 (Suppl. S2), 301–313. [Google Scholar] [PubMed]
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: A brief overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef]
- Fraga, C.G. Relevance, essentiality and toxicity of trace elements in human health. Mol. Aspects Med. 2005, 26, 235–244. [Google Scholar] [CrossRef]
- Kim, J.-J.; Kim, Y.-S.; Kumar, V. Heavy metal toxicity: An update of chelating therapeutic strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef]
- Palacios, O.; Atrian, S.; Capdevila, M. Zn- and Cu-thioneins: A functional classification. J. Biol. Inorg. Chem. 2011, 16, 991–1009. [Google Scholar] [CrossRef] [PubMed]
- Amadi, C.N.; Offor, S.J.; Frazzoli, C.; Orisakwe, O.E. Natural antidotes and management of metal toxicity. Environ. Sci. Pollut. Res. Int. 2019, 26, 18032–18052. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Mila, I.; Expert, D.; Marmolle, F.; Albrecht, A.M.; Hurrell, R.; Huneau, J.F.; Tomé, D. Polyphenols, metal ion complexation and biological consequences. Basic Life Sci. 1999, 66, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Liu, Z.D.; Khodr, H.H. Metal chelation of polyphenols. Methods Enzymol. 2001, 335, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Gomes de Moura, C.F.; Ribeiro, D.A. Are food compounds able to modulate noxious activities induced by cadmium exposure? Crit. Rev. Food Sci. Nutr. 2017, 57, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, X.; Sun, J.; Zhu, C.; Li, X.; Tian, L.; Liu, L.; Bai, W. Cytoprotective effects of dietary flavonoids against cadmium-induced toxicity. Ann. N. Y. Acad. Sci. 2017, 1398, 5–19. [Google Scholar] [CrossRef]
- Oprea, E.; Ruta, L.L.; Nicolau, I.; Popa, C.V.; Neagoe, A.D.; Farcasanu, I.C. Vaccinium corymbosum L. (blueberry) extracts exhibit protective action against cadmium toxicity in Saccharomyces cerevisiae cells. Food Chem. 2014, 152, 516–521. [Google Scholar] [CrossRef]
- Fernandez, M.T.; Mira, M.L.; Florêncio, M.H.; Jennings, K.R. Iron and copper chelation by flavonoids: An electrospray mass spectrometry study. J. Inorg. Biochem. 2002, 92, 105–111. [Google Scholar] [CrossRef]
- Khan, G.N.; Merajver, S.D. Modulation of angiogenesis for cancer prevention: Strategies based on antioxidants and copper deficiency. Curr. Pharm. Des. 2007, 13, 3584–3590. [Google Scholar] [CrossRef]
- Imam, M.U.; Zhang, S.; Ma, J.; Wang, H.; Wang, F. Antioxidants mediate both iron homeostasis and oxidative stress. Nutrients 2017, 9, 671. [Google Scholar] [CrossRef]
- Mandel, S.; Weinreb, O.; Reznichenko, L.; Kalfon, L.; Amit, T. Green tea catechins as brain-permeable, non toxic iron chelators to “iron out iron” from the brain. J. Neural. Transm. Suppl. 2006, 71, 249–257. [Google Scholar] [CrossRef]
- Peres, T.V.; Schettinger, M.R.; Chen, P.; Carvalho, F.; Avila, D.S.; Bowman, A.B.; Aschner, M. Manganese-induced neurotoxicity: A review of its behavioral consequences and neuroprotective strategies. BMC Pharmacol. Toxicol. 2016, 17, 57. [Google Scholar] [CrossRef] [Green Version]
- Maulik, M.; Mitra, S.; Sweeney, M.; Lu, B.; Taylor, B.E.; Bult-Ito, A. Complex interaction of dietary fat and Alaskan bog blueberry supplementation influences manganese mediated neurotoxicity and behavioral impairments. J. Funct. Foods 2019, 53, 306–317. [Google Scholar] [CrossRef]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Fruit polyphenols: A review of anti-inflammatory effects in humans. Crit. Rev. Food Sci. Nutr. 2016, 56, 419–444. [Google Scholar] [CrossRef]
- Oliviero, F.; Scanu, A.; Zamudio-Cuevas, Y.; Punzi, L.; Spinella, P. Anti-inflammatory effects of polyphenols in arthritis. J. Sci. Food Agric. 2018, 98, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Masek, E.; Ebersole, J.L. Dietary polyphenols and periodontitis-a mini-review of literature. Molecules 2018, 23, 1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J. Dietary polyphenols, mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxid. Med. Cell Longev. 2017, 2017, 6723931. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and glycemic control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Tressera-Rimbau, A.; Arranz, S.; Eder, M.; Vallverdú-Queralt, A. Dietary polyphenols in the prevention of stroke. Oxid. Med. Cell Longev. 2017, 2017, 7467962. [Google Scholar] [CrossRef] [Green Version]
- Manolescu, B.N.; Oprea, E.; Mititelu, M.; Ruta, L.L.; Farcasanu, I.C. Dietary anthocyanins and stroke: A review of pharmacokinetic and pharmacodynamic studies. Nutrients 2019, 11, 1479. [Google Scholar] [CrossRef] [Green Version]
- Casas, R.; Castro-Barquero, S.; Estruch, R.; Sacanella, E. Nutrition and Cardiovascular Health. Int. J. Mol. Sci. 2018, 19, 3988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braakhuis, A.J.; Campion, P.; Bishop, K.S. Reducing breast cancer recurrence: The role of dietary polyphenolics. Nutrients 2016, 8, 547. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Knaze, V.; Zamora-Ros, R. Polyphenols: Dietary assessment and role in the prevention of cancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 512–521. [Google Scholar] [CrossRef]
- Daglia, M.; Di Lorenzo, A.; Nabavi, S.F.; Talas, Z.S.; Nabavi, S.M. Polyphenols: Well beyond the antioxidant capacity: Gallic acid and related compounds as neuroprotective agents: You are what you eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef]
- Pawlowska, E.; Szczepanska, J.; Koskela, A.; Kaarniranta, K.; Blasiak, J. Dietary polyphenols in age-related macular degeneration: Protection against oxidative stress and beyond. Oxid. Med. Cell Longev. 2019, 2019, 9682318. [Google Scholar] [CrossRef]
- Braakhuis, A.J.; Hopkins, W.G. Impact of dietary antioxidants on sport performance: A review. Sports Med. 2015, 45, 939–955. [Google Scholar] [CrossRef]
- Bowtell, J.; Kelly, V. Fruit-derived polyphenol supplementation for athlete recovery and performance. Sports Med. 2019, 49 (Suppl. S1), 3–23. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.S.; Ho, C.T.; Zhang, J.; Wan, X.; Zhang, K.; Lim, J. Antioxidants: Differing meanings in food science and health science. J. Agric. Food Chem. 2018, 66, 3063–3068. [Google Scholar] [CrossRef]
- Ames, P.R.J.; Bucci, T.; Merashli, M.; Amaral, M.; Arcaro, A.; Gentile, F.; Nourooz-Zadeh, J.; Delgado Alves, J. Oxidative/nitrative stress in the pathogenesis of systemic sclerosis: Are antioxidants beneficial? Free Radic. Res. 2018, 52, 1063–1082. [Google Scholar] [CrossRef]
- Poljsak, B.; Milisav, I. The neglected significance of “antioxidative stress”. Oxid. Med. Cell. Longev. 2012, 2012, 480895. [Google Scholar] [CrossRef] [Green Version]
- Braconi, D.; Bernardini, G.; Santucci, A. Saccharomyces cerevisiae as a model in ecotoxicological studies: A post-genomics perspective. J. Proteom. 2016, 137, 19–34. [Google Scholar] [CrossRef]
- Nielsen, J. Yeast systems biology: Model organism and cell factory. Biotechnol. J. 2019, 14, e1800421. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Liu, X.; Zhan, J.; Li, J.; Huang, W. The effect of proanthocyanidins on growth and alcoholic fermentation of wine yeast under copper stress. J. Food Sci. 2015, 80, M1319–M1324. [Google Scholar] [CrossRef] [PubMed]
- Farcasanu, I.C.; Mizunuma, M.; Nishiyama, F.; Miyakawa, T. Role of L-histidine in conferring tolerance to Ni2+ in Sacchromyces cerevisiae cells. Biosci. Biotechnol. Biochem. 2005, 69, 2343–2348. [Google Scholar] [CrossRef] [PubMed]
- Minear, S.; O’Donnell, A.F.; Ballew, A.; Giaever, G.; Nislow, C.; Stearns, T.; Cyert, M.S. Curcumin inhibits growth of Saccharomyces cerevisiae through iron chelation. Eukaryot. Cell 2011, 10, 1574–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ho, A.; Ward, D.M.; Kaplan, J. Transition metal transport in yeast. Annu. Rev. Microbiol. 2002, 56, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Dancis, A.; Haile, D.; Yuan, D.S.; Klausner, R.D. The Saccharomyces cerevisiae copper transport protein (Ctr1p). Biochemical characterization, regulation by copper, and physiologic role in copper uptake. J. Biol. Chem. 1994, 269, 25660–25667. [Google Scholar]
- Eide, D.J. The molecular biology of metal ion transport in Saccharomyces cerevisiae. Annu. Rev. Nutr. 1998, 18, 441–469. [Google Scholar] [CrossRef]
- Hassett, R.; Dix, D.R.; Eide, D.J.; Kosman, D.J. The Fe(II) permease Fet4p functions as a low affinity copper transporter and supports normal copper trafficking in Saccharomyces cerevisiae. Biochem. J. 2000, 351 Pt 2, 477–484. [Google Scholar] [CrossRef]
- Jensen, L.T.; Ajua-Alemanji, M.; Culotta, V.C. The Saccharomyces cerevisiae high affinity phosphate transporter encoded by PHO84 also functions in manganese homeostasis. J. Biol. Chem. 2003, 278, 42036–42040. [Google Scholar] [CrossRef] [Green Version]
- Ofiteru, A.M.; Ruta, L.L.; Rotaru, C.; Dumitru, I.; Ene, C.D.; Neagoe, A.; Farcasanu, I.C. Overexpression of the PHO84 gene causes heavy metal accumulation and induces Ire1p-dependent unfolded protein response in Saccharomyces cerevisiae cells. Appl. Microbiol. Biotechnol. 2012, 94, 425–435. [Google Scholar] [CrossRef]
- Supek, F.; Supekova, L.; Nelson, H.; Nelson, N. A yeast manganese transporter related to the macrophage protein involved in conferring resistance to mycobacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 5105–5110. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Eide, D. The yeast ZRT1 gene encodes the zinc transporter protein of a high-affinity uptake system induced by zinc limitation. Proc. Natl. Acad. Sci. USA 1996, 93, 2454–2458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Eide, D. The ZRT2 gene encodes the low affinity zinc transporter in Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 23203–23210. [Google Scholar] [CrossRef] [Green Version]
- Sherman, F. Getting started with yeast. Methods Enzymol. 2002, 350, 3–41. [Google Scholar] [CrossRef] [PubMed]
- Available online: www.euroscarf.de (accessed on 1 July 2018).
- Ruta, L.L.; Kissen, R.; Nicolau, I.; Neagoe, A.D.; Petrescu, A.J.; Bones, A.M.; Farcasanu, I.C. Heavy metal accumulation by Saccharomyces cerevisiae cells armed with metal binding hexapeptides targeted to the inner face of the plasma membrane. Appl. Microbiol. Biotechnol. 2017, 101, 5749–5763. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef]
- Eide, D.J. Multiple regulatory mechanisms maintain zinc homeostasis in Saccharomyces cerevisiae. J. Nutr. 2003, 133 (Suppl. 1), 1532S–1535S. [Google Scholar] [CrossRef]
- Wu, C.Y.; Bird, A.J.; Chung, L.M.; Newton, M.A.; Winge, D.R.; Eide, D.J. Differential control of Zap1-regulated genes in response to zinc deficiency in Saccharomyces cerevisiae. BMC Genom. 2008, 9, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, A.G.; Eide, D.J. Roles of two activation domains in Zap1 in the response to zinc deficiency in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 6844–6854. [Google Scholar] [CrossRef] [Green Version]
- Lapinskas, P.J.; Cunningham, K.W.; Liu, X.F.; Fink, G.R.; Culotta, V.C. Mutations in PMR1 suppress oxidative damage in yeast cells lacking superoxide dismutase. Mol. Cell. Biol. 1995, 15, 1382–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farcasanu, I.C.; Hirata, D.; Tsuchiya, E.; Mizuta, K.; Miyakawa, T. Involvement of thioredoxin peroxidase type II (Ahp1p) of Saccharomyces cerevisiae in Mn2+ homeostasis. Biosci. Biotechnol. Biochem. 1999, 63, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, C.; Liba, A.; Imlay, J.A.; Valentine, J.S.; Gralla, E.B. Yeast lacking superoxide dismutase(s) show elevated levels of “free iron” as measured by whole cell electron paramagnetic resonance. J. Biol. Chem. 2000, 275, 29187–29192. [Google Scholar] [CrossRef] [Green Version]
- Ruta, L.L.; Popa, C.V.; Nicolau, I.; Farcasanu, I.C. Epigallocatechin-3-O-gallate, the main green tea component, is toxic to Saccharomyces cerevisiae cells lacking the Fet3/Ftr1. Food Chem. 2018, 266, 292–298. [Google Scholar] [CrossRef]
- Borowska, S.; Brzoska, M.M.; Tomczyk, M. Complexation of bioelements and toxic metals by polyphenolic compounds—Implications for health. Curr. Drug Targets 2018, 19, 1612–1638. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.C.; Chao, P.D.; Ho, H.J.; Wen, C.C.; Hsiu, S.L. Profound difference in pharmacokinetics between morin and its isomer quercetin in rats. J. Pharm. Pharmacol. 2003, 55, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Menon, L.G.; Kuttan, R.; Kuttan, G. Inhibition of lung metastasis in mice induced by B16F10 melanoma cells by polyphenolic compounds. Cancer Lett. 1995, 95, 221–225. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shen, S.C.; Chow, J.M.; Ko, C.H.; Tseng, S.W. Flavone inhibition of tumor growth via apoptosis in vitro and in vivo. Int. J. Oncol. 2004, 25, 661–670. [Google Scholar] [CrossRef]
- Yao, D.; Cui, H.; Zhou, S.; Guo, L. Morin inhibited lung cancer cells viability, growth, and migration by suppressing miR-135b and inducing its target CCNG2. Tumour Biol. 2017, 39, 1010428317712443. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zhang, Y. Morin inhibits ovarian cancer growth through the inhibition of NF-κB signaling pathway. Anticancer Agents Med. Chem. 2019, 19, 2243–2250. [Google Scholar] [CrossRef]
- Tang, S.M.; Deng, X.T.; Zhou, J.; Li, Q.P.; Ge, X.X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef]
- Gibson, R.S.; King, J.C.; Lowe, N. A review of dietary zinc recommendations. Food Nutr. Bull. 2016, 37, 443–460. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strain | Gene Deleted | Gene Function | Growth in the Presence of Antioxidant (%) | Antioxidant Tested |
|---|---|---|---|---|
| Wild Type 1 | None | Control strain | 0.94 ± 0.22 | Caffeic acid |
| 0.88 ± 0.14 | Chlorogenic acid | |||
| 0.96 ± 0.22 | Cyanidin | |||
| 0.89 ± 0.25 | (−)-Epicatechin | |||
| 1.12 ± 0.21 | Epigallocatechin-3-O-gallate | |||
| 0.91 ± 0.24 | Gallic acid | |||
| 1.24 ± 0.14 | Morin | |||
| 1.28 ± 0.26 | Quercetin | |||
| 0.95 ± 0.14 | Resveratrol | |||
| 0.96 ± 0.23 | Rutin | |||
| Genes Involved in Heavy Metal Transport | ||||
| ctr1Δ | CTR1 | High-affinity copper transporter of plasma membrane | 0.92 ± 0.24 | Caffeic acid |
| 0.89 ± 0.12 | Chlorogenic acid | |||
| 0.93 ± 0.21 | Cyanidin | |||
| 0.91 ± 0.18 | (−)-Epicatechin | |||
| 0.88 ± 0.17 | Epigallocatechin-3-O-gallate | |||
| 0.93 ± 0.22 | Gallic acid | |||
| 0.96 ± 0.14 | Morin | |||
| 1.08 ± 0.16 | Quercetin | |||
| 0.91 ± 0.14 | Resveratrol | |||
| 0.90 ± 0.17 | Rutin | |||
| ftr1Δ | FTR1 | High-affinity iron transporter of the plasma membrane | 0.90 ± 0.17 | Caffeic acid |
| 0.91 ± 0.14 | Chlorogenic acid | |||
| 0.93 ± 0.16 | Cyanidin | |||
| 0.92 ± 0.15 | (−)-Epicatechin | |||
| 0.78 ± 0.34 | Epigallocatechin-3-O-gallate | |||
| 0.84 ± 0.26 | Gallic acid | |||
| 0.84 ± 0.25 | Morin | |||
| 0.88 ± 0.28 | Quercetin | |||
| 0.91 ± 0.16 | Resveratrol | |||
| 0.93 ± 0.24 | Rutin | |||
| fet4Δ | FET41 | Low-affinity iron transporter of the plasma membrane | 0.92 ± 0.23 | Caffeic acid |
| 0.98 ± 0.17 | Chlorogenic acid | |||
| 0.94 ± 0.16 | Cyanidin | |||
| 0.92 ± 0.15 | (−)-Epicatechin | |||
| 0.91 ± 0.21 | Epigallocatechin-3-O-gallate | |||
| 0.90 ± 0.28 | Gallic acid | |||
| 1.43 ± 0.16 * | Morin * | |||
| 1.45 ± 0.25 * | Quercetin * | |||
| 0.98 ± 0.24 | Resveratrol | |||
| 0.94 ± 0.22 | Rutin | |||
| pho84Δ | PHO84 | High-affinity inorganic phosphate (Pi) transporter and low-affinity divalent metal ion transporter | 0.80 ± 0.23 | Caffeic acid |
| 0.78 ± 0.14 | Chlorogenic acid | |||
| 0.76 ± 0.22 | Cyanidin | |||
| 0.79 ± 0.25 | (−)-Epicatechin | |||
| 0.82 ± 0.21 | Epigallocatechin-3-O-gallate | |||
| 0.85 ± 0.24 | Gallic acid | |||
| 0.78 ± 0.21 | Morin | |||
| 0.79 ± 0.25 | Quercetin | |||
| 0.85 ± 0.14 | Resveratrol | |||
| 0.84 ± 0.21 | Rutin | |||
| smf1Δ | SMF1 | Divalent metal ion transporter | 0.92 ± 0.25 | Caffeic acid |
| 0.94 ± 0.14 | Chlorogenic acid | |||
| 1.11 ± 0.21 | Cyanidin | |||
| 0.93 ± 0.15 | (−)-Epicatechin | |||
| 1.02 ± 0.20 | Epigallocatechin-3-O-gallate | |||
| 0.91 ± 0.24 | Gallic acid | |||
| 1.56 ± 0.34 * | Morin * | |||
| 1.47 ± 0.23 * | Quercetin * | |||
| 0.98 ± 0.24 | Resveratrol | |||
| 0.95 ± 0.21 | Rutin | |||
| ztr1Δ | ZTR1 | High-affinity Zn2+ transporter of the plasma membrane | 0.94 ± 0.22 | Caffeic acid |
| 0.88 ± 0.14 | Chlorogenic acid | |||
| 0.96 ± 0.22 | Cyanidin | |||
| 0.89 ± 0.25 | (−)-Epicatechin | |||
| 1.12 ±0.21 | Epigallocatechin-3-O-gallate | |||
| 0.91 ± 0.24 | Gallic acid | |||
| 1.48 ± 0.22 * | Morin * | |||
| 1.46 ± 0.16 * | Quercetin * | |||
| 0.95 ± 0.14 | Resveratrol | |||
| 0.96 ± 0.23 | Rutin | |||
| ztr2Δ | ZTR2 | Low-affinity Zn2+ transporter of the plasma membrane | 0.61 ± 0.14# | Caffeic acid# |
| 0.88 ± 0.14 | Chlorogenic acid | |||
| 0.96 ± 0.22 | Cyanidin | |||
| 0.91 ± 0.15 | (−)-Epicatechin | |||
| 0.92 ± 0.21 | Epigallocatechin-3-O-gallate | |||
| 0.65 ± 0.24# | Gallic acid# | |||
| 1.24 ± 0.14 | Morin | |||
| 1.15 ± 0.26 | Quercetin | |||
| 0.95 ± 0.14 | Resveratrol | |||
| 0.96 ± 0.23 | Rutin | |||
| Strain | Antioxidant | Cellular Metal Content (nmol/mg Cell Total Protein) | |||
|---|---|---|---|---|---|
| Cu | Fe | Mn | Zn | ||
| WT | None | 5.84 ± 0.42 | 52.82 ± 3.24 | 6.25 ± 0.62 | 12.42 ± 1.14 |
| Caffeic acid | 5.21 ± 0.31 | 50.78 ± 5.82 | 5.96 ± 0.38 | 8.48 ± 1.04 | |
| Gallic acid | 4.98 ± 0.52 | 49.34 ± 4.21 | 5.28 ± 0.51 | 9.25 ± 0.98 | |
| Morin | 4.82 ± 0.42 | 48.38 ± 4.54 | 4.21 ± 0.28 | 11.48 ± 1.16 | |
| Quercetin | 4.65 ± 0.63 | 47.22 ± 3.98 | 4.10 ± 0.31 | 11.22 ± 1.28 | |
| fet4Δ | None | 5.24 ± 0.35 | 51.34 ± 2.81 | 4.25 ± 0.57 # | 11.48 ± 1.23 |
| Caffeic acid | 5.04 ± 0.24 | 49.33 ± 3.62 | 4.15 ± 0.27 | 10.32 ± 1.11 | |
| Gallic acid | 4.52 ± 0.38 | 48.68 ± 3.29 | 4.28 ± 0.42 | 9.47 ± 0.87 | |
| Morin | 4.32 ± 0.33 | 48.02 ± 3.65 | 2.21 ± 0.36 #,* | 9.84 ± 0.92 | |
| Quercetin | 4.14 ± 0.41 | 47.82 ± 3.62 | 2.11 ± 0.31 #,* | 9.68 ± 1.08 | |
| smf1Δ | None | 4.34 ± 0.42 | 50.82 ± 4.15 | 3.16 ± 0.51 # | 9.42 ± 1.21 |
| Caffeic acid | 4.02 ± 0.34 | 50.31 ± 3.87 | 3,37 ± 0.39 # | 8.72 ± 1.05 | |
| Gallic acid | 4.23 ± 0.38 | 48.34 ± 4.49 | 3.23 ± 0.47 # | 8.25 ± 0.91 | |
| Morin | 4.56 ± 0.45 | 46.65 ± 4.39 | 1.21 ± 0.18 ##,* | 8.43 ± 1.11 | |
| Quercetin | 4.37 ± 0.45 | 45.92 ± 3.77 | 1.10 ± 0.31 ##,* | 8.22 ± 1.07 | |
| zrt1Δ | None | 5.92 ± 0.53 | 53.92 ± 4.94 | 5.32 ± 0.42 | 8.68 ± 1.04 # |
| Caffeic acid | 4.29 ± 0.38 | 52.75 ± 4.83 | 4.91 ± 0.37 | 7.37 ± 0.95 # | |
| Gallic acid | 4.56 ± 0.45 | 50.68 ± 5.21 | 4.78 ± 0.51 | 7.25 ± 0.92 # | |
| Morin | 4.82 ± 0.42 | 50.08 ± 4.59 | 4.57 ± 0.45 | 6.38 ± 0.56 #,* | |
| Quercetin | 4.68 ± 0.63 | 49.97 ± 4.98 | 4.71 ± 0.43 | 6.22 ± 0.73 #,* | |
| zrt2Δ | None | 4.62 ± 0.51 | 54.31 ± 4.14 | 5.72 ± 0.84 | 10.61 ± 0.89 |
| Caffeic acid | 4.22 ± 0.44 | 55.25 ± 4.32 | 5.38 ± 0.65 | 3.47 ± 1.02 ##,** | |
| Gallic acid | 4.17 ± 0.42 | 51.44 ± 6.12 | 5.09 ± 0.61 | 4.25 ± 0.93 ##,** | |
| Morin | 4.22 ± 0.40 | 50.25 ± 5.24 | 2.98 ± 0.28 * | 9.26 ± 1.05 | |
| Quercetin | 4.12 ± 0.51 | 50.86 ± 4.63 | 2.34 ± 0.22 * | 9.12 ± 1.28 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruta, L.L.; Farcasanu, I.C. Interaction between Polyphenolic Antioxidants and Saccharomyces cerevisiae Cells Defective in Heavy Metal Transport across the Plasma Membrane. Biomolecules 2020, 10, 1512. https://doi.org/10.3390/biom10111512
Ruta LL, Farcasanu IC. Interaction between Polyphenolic Antioxidants and Saccharomyces cerevisiae Cells Defective in Heavy Metal Transport across the Plasma Membrane. Biomolecules. 2020; 10(11):1512. https://doi.org/10.3390/biom10111512
Chicago/Turabian StyleRuta, Lavinia Liliana, and Ileana Cornelia Farcasanu. 2020. "Interaction between Polyphenolic Antioxidants and Saccharomyces cerevisiae Cells Defective in Heavy Metal Transport across the Plasma Membrane" Biomolecules 10, no. 11: 1512. https://doi.org/10.3390/biom10111512
APA StyleRuta, L. L., & Farcasanu, I. C. (2020). Interaction between Polyphenolic Antioxidants and Saccharomyces cerevisiae Cells Defective in Heavy Metal Transport across the Plasma Membrane. Biomolecules, 10(11), 1512. https://doi.org/10.3390/biom10111512