A Comparative View on the Oviductal Environment during the Periconception Period

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Impact of Embryo and Oviduct Communication
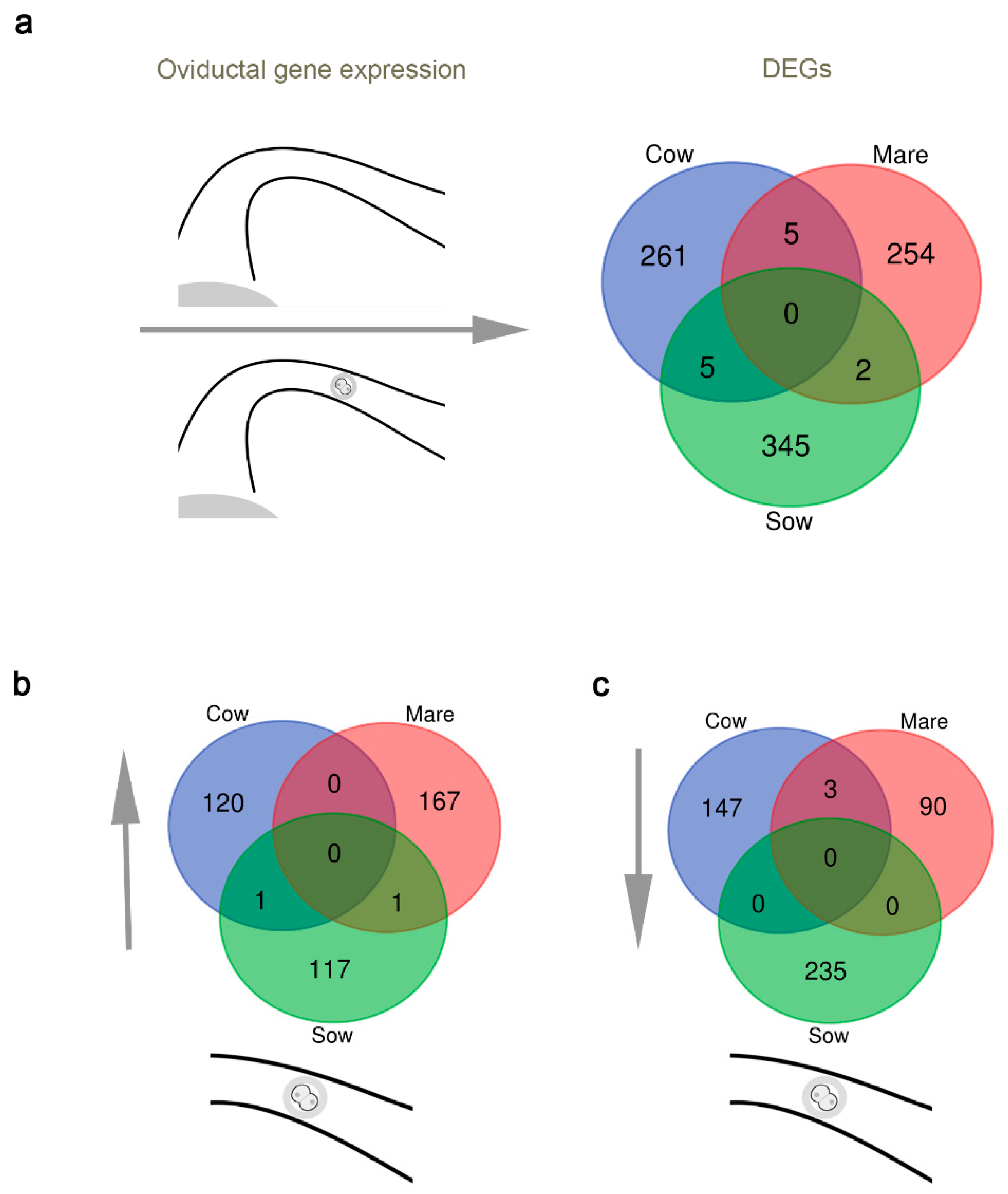
3.1. A Comparative View of the Impact of the Embryo on Oviductal Gene Expression
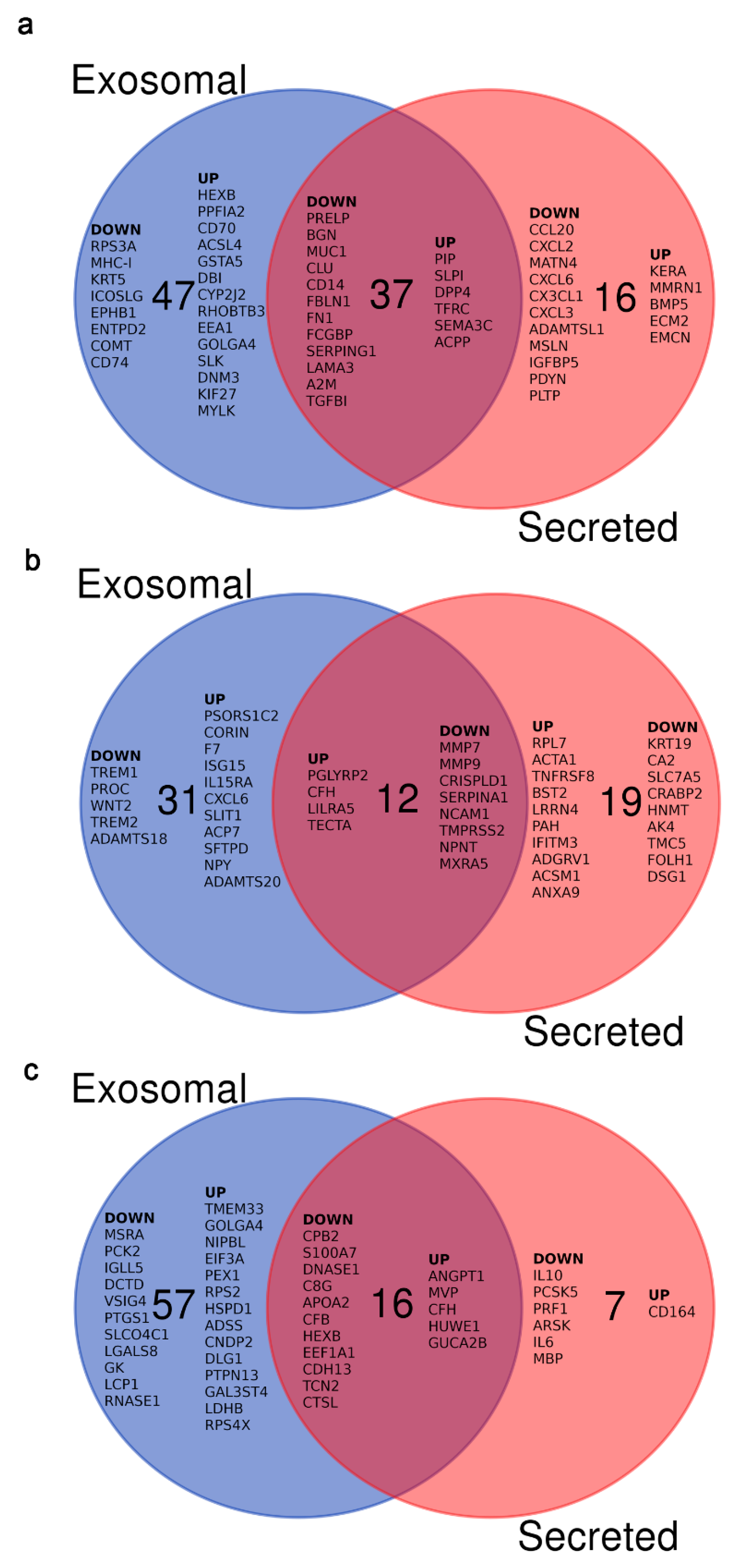
3.2. Impact of the Oviduct on the Embryo
4. OVGP1
4.1. Origin and Localization of OVGP1
4.2. Characterization of OVGP1
4.3. Effect of OVGP1 Localized in Eggs and Embryos
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Date Availability
References
- Woodruff, J.D.; Pauerstein, C.J. The Fallopian Tube: Structure, Function, Pathology, and Management; Williams & Wilkins Co.: Washington, DC, USA, 1969. [Google Scholar]
- Hunter, R.H.F. Have the Falopian Tubes a Vital Role in Promoting Fertility? Acta Obs. Gynecol. Scand. 1998, 77, 475–486. [Google Scholar]
- Avilés, M.; Gutiérrez-Adán, A.; Coy, P. Oviductal Secretions: Will They Be Key Factors for the Future ARTs? Mol. Hum. Reprod. 2010, 16, 896–906. [Google Scholar] [CrossRef]
- Holt, W.V.; Fazeli, A. The Oviduct as a Complex Mediator of Mammalian Sperm Function and Selection. Mol. Reprod. Dev. 2010, 77, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; Garcia-Vasquez, F.A.; Visconti, P.E.; Aviles, M. Roles of the Oviduct in Mammalian Fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillo, V.; De Frutos, C.; O’Gaora, P.; Forde, N.; Burns, G.W.; Spencer, T.E.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Spatial Differences in Gene Expression in the Bovine Oviduct. Reproduction 2016, 152, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alminana-Brines, C.; Corbin, E.; Tsikis, G.; Neto, A.S.D.A.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct Extracellular Vesicles Protein Content and Their Role During oviduct–embryo Cross-Talk. Reproduction 2017, 154, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Almiñana, C.; Tsikis, G.; Labas, V.; Uzbekov, R.E.; Da Silveira, J.C.; Bauersachs, S.; Mermillod, P. Deciphering the Oviductal Extracellular Vesicles Content across the Estrous Cycle: Implications for the Gametes-Oviduct Interactions and the Environment of the Potential Embryo. BMC Genom. 2018, 19, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Cerezales, S.; Ramos-Ibeas, P.; Acuña, O.S.; Avilés, M.; Coy, P.; Rizos, D.; Gutiérrez-Adán, A. The Oviduct: From Sperm Selection to the Epigenetic Landscape of the embryo†. Biol. Reprod. 2018, 98, 262–276. [Google Scholar] [CrossRef] [Green Version]
- Duranthon, V.; Watson, A.J.; Lonergan, P. Preimplantation Embryo Programming: Transcription, Epigenetics, and Culture Environment. Reproduction 2008, 135, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.T.; Bonneau, A.R.; Takacs, C.M.; Bazzini, A.A.; DiVito, K.R.; Fleming, E.S.; Giraldez, A.J. Nanog, Pou5f1 and SoxB1 Activate Zygotic Gene Expression During the Maternal-to-Zygotic Transition. Nat. Cell Biol. 2013, 503, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Ko, M.S.H. Zygotic Genome Activation Revisited: Looking through the Expression and Function of Zscan4. In Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2016; Volume 120, pp. 103–124. [Google Scholar]
- Wu, E.; Vastenhouw, N.L. From Mother to Embryo: A Molecular Perspective on Zygotic Genome Activation. Curr. Top. Dev. Biol. 2020, 209–254. [Google Scholar] [CrossRef]
- Salilew-Wondim, D.; Saeed-Zidane, M.; Hoelker, M.; Gebremedhn, S.; Poirier, M.; Pandey, H.O.; Tholen, E.; Neuhoff, C.; Held, E.; Besenfelder, U.; et al. Genome-Wide DNA Methylation Patterns of Bovine Blastocysts Derived from in Vivo Embryos Subjected to in Vitro Culture Before, During or After Embryonic Genome Activation. BMC Genom. 2018, 19, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugentobler, S.A.; Diskin, M.G.; Leese, H.J.; Humpherson, P.G.; Watson, T.; Sreenan, J.M.; Morris, D.G. Amino Acids in Oviduct and Uterine Fluid and Blood Plasma During the Estrous Cycle in the Bovine. Mol. Reprod. Dev. 2007, 74, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Simintiras, C.A.; Fröhlich, T.; Sathyapalan, T.; Arnold, G.J.; Ulbrich, S.; Leese, H.J.; Sturmey, R.G. Modelling Aspects of Oviduct Fluid Formation in Vitro. Reproduction 2017, 153, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. Int. J. Mol. Sci. 2019, 21, 223. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J.; Tay, J.I.; Reischl, J.; Downing, S.J. Formation of Fallopian Tubal Fluid: Role of a Neglected Epithelium. Reproduction 2001, 121, 339–346. [Google Scholar] [CrossRef]
- Leese, H.J.; Hugentobler, S.A.; Gray, S.M.; Morris, D.G.; Sturmey, R.G.; Whitear, S.-L.; Sreenan, J.M. Female Reproductive Tract Fluids: Composition, Mechanism of Formation and Potential Role in the Developmental Origins of Health and Disease. Reprod. Fertil. Dev. 2008, 20, 1–8. [Google Scholar] [CrossRef]
- Almiñana, C.; Bauersachs, S. Extracellular Vesicles in the Oviduct: Progress, Challenges and Implications for the Reproductive Success. Bioengineering 2019, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, K.; Kashiwazaki, N.; Noguchi, J.; Shimada, A.; Takahashi, R.; Hirabayashi, M.; Shino, M.; Ueda, M.; Kaneko, H. Developmental Competence, After Transfer to Recipients, of Porcine Oocytes Matured, Fertilized, and Cultured in Vitro. Biol. Reprod. 1999, 60, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Lonergan, P. State-of-the-Art Embryo Technologies in Cattle. Biosci. Proc. 2019, 64. [Google Scholar] [CrossRef]
- Urrego, R.; Rodríguez-Osorio, N.; Niemann, H. Epigenetic Disorders and Altered Gene Expression After Use of Assisted Reproductive Technologies in Domestic Cattle. Epigenetics 2014, 9, 803–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, S.; Ivanova, E.; Romar, R.; García-Martínez, S.; Soriano-Úbeda, C.; García-Vázquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. Author Response: DNA Methylation and Gene Expression Changes Derived from Assisted Reproductive Technologies Can Be Decreased by Reproductive Fluids. eLife 2017, 6, e23670. [Google Scholar] [CrossRef] [PubMed]
- Romar, R.; Funahashi, H.; Coy, P. In Vitro Fertilization in Pigs: New Molecules and Protocols to Consider in the Forthcoming Years. Theriogenology 2016, 85, 125–134. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T.; Forde, N.; Rizos, D. Embryo Development in Dairy Cattle. Theriogenology 2016, 86, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.M.; Brom-De-Luna, J.G.; Resende, H.L.; Canesin, H.S.; Hinrichs, K. Lower Blastocyst Quality After Conventional Vs. Piezo ICSI in the Horse Reflects Delayed Sperm Component Remodeling and Oocyte Activation. J. Assist. Reprod. Genet. 2018, 35, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Harada, Y.; Maeda, T.; Fukunaga, E.; Shiba, R.; Okano, S.; Kinutani, M.; Horiuchi, T. Selection of High-Quality and Viable Blastocysts Based on Timing of Morula Compaction and Blastocyst Formation. Reprod. Med. Biol. 2019, 19, 58–64. [Google Scholar] [CrossRef]
- Lewis, W. On the Early Development of the Mouse Egg. Carnegie Inst. Contrib. Embryol. 1935, 25, 113–143. [Google Scholar]
- Hunter, R.H.F. Chronological and Cytological Details of Fertilization and Early Embryonic Development in the Domestic pig, Sus Scrofa. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1974, 178, 169–185. [Google Scholar] [CrossRef]
- Rizos, D.; Maillo, V.; Sánchez-Calabuig, M.J.; Lonergan, P. The Consequences of Maternal-Embryonic Cross Talk During the Periconception Period on Subsequent Embryonic Development. Adv. Exp. Med. Biol. 2017, 1014, 69–86. [Google Scholar]
- Clark, R.T. Studies on the Physiology of Reproduction in the Sheep II. The Cleavage Stages of the Ovum. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1934, 60, 135–159. [Google Scholar] [CrossRef]
- Webel, S.K.; Franklin, V.; Harland, B.; Dziuk, P.J. Fertility, Ovulation and Maturation of Eggs in Mares Injected With HCG. Reproduction 1977, 51, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, W.F.; Roth, T.L.; Wildt, D.E. In Vivo Embryogenesis, Embryo Migration, and Embryonic Mortality in the Domestic Cat1. Biol. Reprod. 1994, 51, 452–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynaud, K.; Fontbonne, A.; Marseloo, N.; Thoumire, S.; Chebrout, M.; De Lesegno, C.V.; Chastant-Maillard, S. In Vivo Meiotic Resumption, Fertilization and Early Embryonic Development in the Bitch. Reproduction 2005, 130, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, O.; Ogura, A.; Asano, T.; Noguchi, Y.; Yamamoto, Y.; Oike, M. Development of Preimplantation Guinea-Pig Embryos in Serum-Free Media. Reprod. Fertil. Dev. 1993, 5, 425–432. [Google Scholar] [CrossRef]
- Sultana, F.; Hatori, M.; Shimozawa, N.; Ebisawa, T.; Sankai, T. Continuous Observation of Rabbit Preimplantation Embryos In Vitro by Using a Culture Device Connected to a Microscope. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 52–56. [Google Scholar]
- Hlinka, D.; Kaľatová, B.; Uhrinová, I.; Dolinská, S.; Rutarová, J.; Řezáčová, J.; Lazarovská, S.; Dudáš, M. Time-Lapse Cleavage Rating Predicts Human Embryo Viability. Physiol. Res. 2012, 61, 513–525. [Google Scholar] [CrossRef]
- Hamilton, W.; Laing, J. Development of the Egg of the Cow up to the Stage of Blastocyst Formation. J. Anat. 1946, 80, 194. [Google Scholar]
- Betteridge, K.J.; Eaglesome, M.D.; Mitchell, D.; Flood, P.F.; Beriault, R. Development of Horse Embryos up to Twenty Two Days After Ovulation: Observations on Fresh Specimens. J. Anat. 1982, 135, 191–209. [Google Scholar]
- Renton, J.P.; Boyd, J.S.; Eckersall, P.D.; Ferguson, J.M.; Harvey, M.J.A.; Mullaney, J.; Perry, B. Ovulation, Fertilization and Early Embryonic Development in the Bitch (Canis Familiaris). Reproduction 1991, 93, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Denker, H.-W.; Gerdes, H.-J. The Dynamic Structure of Rabbit Blastocyst Coverings. Anat. Embryol. 1979, 157, 15–34. [Google Scholar] [CrossRef]
- Fischer, B.; Chavatte-Palmer, P.; Viebahn, C.; Santos, A.N.; Duranthon, V. Rabbit as a Reproductive Model for Human Health. Reproduction 2012, 144, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Implantation Mechanisms: Insights from the Sheep. Reproduction 2004, 128, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Sathananthan, H.; Gunasheela, S.; Menezes, J. Mechanics of Human Blastocyst Hatching in Vitro. Reprod. Biomed. Online 2003, 7, 228–234. [Google Scholar] [CrossRef]
- Pratt, H.P. Marking Time and Making Space: Chronology and Topography in the Early Mouse Embryo. Adv. Clin. Chem. 1989, 117, 99–130. [Google Scholar]
- Pope, C. Aspects of in Vivo Oocyte Production, Blastocyst Development, and Embryo Transfer in the Cat. Theriogenology 2014, 81, 126–137. [Google Scholar] [CrossRef]
- Concannon, P.; Tsutsui, T.; Shille, V. Embryo Development, Hormonal Requirements and Maternal Responses During Canine Pregnancy. J. Reprod. Fertil. 2001, 57, 169–179. [Google Scholar]
- Blandau, R.J. Observations on Implantation of the Guinea Pig Ovum. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1949, 103, 19–47. [Google Scholar] [CrossRef]
- Adams, C.E. Egg Transfer in The Rabbit. Mamm. Egg Transf. 2018, 1982, 29–48. [Google Scholar]
- Croxatto, H.B. Physiology of Gamete and Embryo Transport through the Fallopian Tube. Reprod. Biomed. Online 2002, 4, 160–169. [Google Scholar] [CrossRef]
- Battut, I.; Colchen, S.; Fiéni, F.; Tainturier, D.; Bruyas, J. Success Rates When Attempting to Nonsurgically Collect Equine Embryos at 144, 156 or 168 Hours After Ovulation. Equine Veter J. 2010, 29, 60–62. [Google Scholar] [CrossRef]
- Tsutsui, T. Studies on the Reproduction in the Dog. V. On Cleavage and Transport of Fertilized Ova in the Oviduct. Jpn. J. Anim. Reprod. 1975, 21, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Harper, M. Gamete and Zygote Transport. In The Physiology of Reproduction; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1988; pp. 103–134. [Google Scholar]
- Wang, H.; Dey, S.K. Roadmap to Embryo Implantation: Clues from Mouse Models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Valtonen, M.; Jalkanen, L. Species-Specific Features of Oestrous Development and Blastogenesis in Domestic Canine Species. J. Reprod. Fertil. 1993, 47, 133–137. [Google Scholar]
- Hunter, R.H.F.; Hunt, D.M.; Chang, M.C. Temporal and Cytological Aspects of Fertilization and Early Development in the Guinea pig, Cavia Porcellus. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1969, 165, 411–429. [Google Scholar] [CrossRef]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of Implantation: Strategies for Successful Pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef]
- Senger, P.L. Pathways to Pregnancy and Parturition, 2nd ed.; Current Conceptions, Inc.: Redmond, OR, USA, 2003. [Google Scholar]
- Denker, H.-W.; Eng, L.A.; Hamner, C.E. Studies on the Early Development and Implantation in the Cat. Brain Struct. Funct. 1978, 154, 39–54. [Google Scholar] [CrossRef]
- Holst, P.A.; Phemister, R.D. The Prenatal Development of the Dog: Preimplantation Events1. Biol. Reprod. 1971, 5, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Cordova, A.; Perreau, C.; Schmaltz-Panneau, B.; Locatelli, Y.; Ponsart, C.; Mermillod, P. Use of an in Vitro Model in Bovine to Evidence a Functional and Molecular Dialogue Between Preimplantation Embryo and Oviduct Epithelial Cells. Gynecol. Obs. Fertil. 2013, 41, 537–539. [Google Scholar] [CrossRef]
- Hamdi, M.; Lopera, R.; Maillo, V.; Núñez, C.; Gutierrez-Adan, A.; Lonergan, P.; Bermejo-Alvarez, P.; Rizos, D. Bovine Oviduct Epithelial Cells: An in Vitro Model to Study Early Embryo-Maternal Communication. Anim. Reprod. 2015, 12, 798. [Google Scholar]
- Romar, R.; Cánovas, S.; Matás, C.; Gadea, J.; Coy, P. Pig in Vitro Fertilization: Where Are We and Where Do We Go? Theriogenology 2019, 137, 113–121. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange Between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Almiñana, C.; Bauersachs, S. Extracellular Vesicles: Multi-Signal Messengers in the gametes/Embryo-Oviduct Cross-Talk. Theriogenology 2020, 150, 59–69. [Google Scholar] [CrossRef]
- Alcântara-Neto, A.S.; Fernandez-Rufete, M.; Corbin, E.; Tsikis, G.; Uzbekov, R.; Garanina, A.S.; Coy, P.; Almiñana, C.; Mermillod, P. Oviduct Fluid Extracellular Vesicles Regulate Polyspermy During Porcine in Vitro Fertilisation. Reprod. Fertil. Dev. 2020, 32, 409–418. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Fernandez-Fuertes, B.; Maillo, V.; Beltrán-Breña, P.; Calle, A.; Redruello, A.; López-Martín, S.; Gutiérrez-Adán, A.; Yáñez-Mó, M.; et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS ONE 2016, 11, e0148083. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of Bovine Oviductal Extracellular Vesicles on Embryo Development and Quality in Vitro. Reproduction 2017, 153, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Lloreda, V.; Coy, P.; Gutiérrez-Adán, A.; Bermejo-Álvarez, P.; Rizos, D. Effect of Bovine Oviductal Fluid on Development and Quality of Bovine Embryos Produced in Vitro. Reprod. Fertil. Dev. 2017, 29, 621. [Google Scholar] [CrossRef]
- Hamdi, M.; Lopera-Vasquez, R.; Maillo, V.; Sánchez-Calabuig, M.J.; Núnez, C.; Gutiérrez-Adán, A.; Rizos, D. Bovine Oviductal and Uterine Fluid Support In Vitro Embryo Development. Reprod. Fertil. Dev. 2018, 30, 935. [Google Scholar] [CrossRef]
- Fazeli, A.; Holt, W.V. Cross Talk During the Periconception Period. Theriogenology 2016, 86, 438–442. [Google Scholar] [CrossRef]
- Maillo, V.; Gaora, P.Ó.; Forde, N.; Besenfelder, U.; Havlicek, V.; Burns, G.W.; Spencer, T.E.; Gutiérrez-Adán, A.; Lonergan, P.; Rizos, D. Oviduct-Embryo Interactions in Cattle: Two-Way Traffic or a One-Way Street? Biol. Reprod. 2015, 92, 144. [Google Scholar] [CrossRef]
- Smits, K.; De Coninck, D.I.; Van Nieuwerburgh, F.; Govaere, J.; Van Poucke, M.; Peelman, L.; Deforce, D.; Van Soom, A. The Equine Embryo Influences Immune-Related Gene Expression in the Oviduct1. Biol. Reprod. 2016, 94, 36. [Google Scholar] [CrossRef] [Green Version]
- Martyniak, M.; Zglejc-Waszak, K.; Franczak, A.; Kotwica, G. Transcriptomic Analysis of the Oviduct of Pigs During the Peri-Conceptional Period. Anim. Reprod. Sci. 2018, 197, 278–289. [Google Scholar] [CrossRef]
- Graf, A.; Krebs, S.; Zakhartchenko, V.; Schwalb, B.; Blum, H.; Wolf, E. Fine Mapping of Genome Activation in Bovine Embryos by RNA Sequencing. Proc. Natl. Acad. Sci. USA 2014, 111, 4139–4144. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Yang, X.; Zhang, H.; Liu, S.; Zhao, J.; Zhang, J.; Weng, X.; Jin, J.; Liu, Z. Lineage Specification and Pluripotency Revealed by Transcriptome Analysis from Oocyte to Blastocyst in Pig. FASEB J. 2019, 34, 691–705. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Huang, K.; Cai, C.; Cai, L.; Jiang, C.-Y.; Feng, Y.; Liu, Z.; Zeng, Q.; Cheng, L.; Sun, Y.E.; et al. Genetic Programs in Human and Mouse Early Embryos Revealed by Single-Cell RNA Sequencing. Nat. Cell Biol. 2013, 500, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Durinck, S.; Spellman, P.T.; Birney, E.; Huber, W. Mapping Identifiers for the Integration of Genomic Datasets With the R/Bioconductor Package BiomaRt. Nat. Protoc. 2009, 4, 1184–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamy, J.; Labas, V.; Harichaux, G.; Tsikis, G.; Mermillod, P.; Saint-Dizier, M. Regulation of the Bovine Oviductal Fluid Proteome. Reproduction 2016, 152, 629–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, V.V.; Weber, D.M.; Phinney, B.S.; Selvaraj, V. Profiling of Proteins Secreted in the Bovine Oviduct Reveals Diverse Functions of This Luminal Microenvironment. PLoS ONE 2017, 12, e0188105. [Google Scholar] [CrossRef] [Green Version]
- Smits, K.; Nelis, H.; Van Steendam, K.; Govaere, J.; Roels, K.; Ververs, C.; Leemans, B.; Wydooghe, E.; Deforce, D.; Van Soom, A. Proteome of Equine Oviducal Fluid: Effects of Ovulation and Pregnancy. Reprod. Fertil. Dev. 2017, 29, 1085. [Google Scholar] [CrossRef]
- Canha-Gouveia, A.; Paradela, A.; Ramos-Fernández, A.; Prieto-Sánchez, M.T.; Sánchez-Ferrer, M.L.; Corrales, F.; Coy, P. Which Low-Abundance Proteins are Present in the Human Milieu of Gamete/Embryo Maternal Interaction? Int. J. Mol. Sci. 2019, 20, 5305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luongo, C.; González-Brusi, L.; Cots-Rodríguez, P.; Izquierdo-Rico, M.J.; Avilés, M.; García-Vázquez, F.A. Sperm Proteome After Interaction with Reproductive Fluids in Porcine: From the Ejaculation to the Fertilization Site. Int. J. Mol. Sci. 2020, 21, 6060. [Google Scholar] [CrossRef] [PubMed]
- Ramilowski, J.A.; Goldberg, T.; Harshbarger, J.; Kloppmann, E.; Lizio, M.; Satagopam, V.P.; Itoh, M.; Kawaji, H.; Carninci, P.; Rost, B.; et al. A Draft Network of ligand–receptor-Mediated Multicellular Signalling in Human. Nat. Commun. 2015, 6, 7866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-F.; Yao, Y.; Kwok, K.-L.; Xu, J.-S.; Yeung, W.S.B. Early Developing Embryos Affect the Gene Expression Patterns in the Mouse Oviduct. Biochem. Biophys. Res. Commun. 2002, 292, 564–570. [Google Scholar] [CrossRef]
- Almiñana, C.; Heath, P.R.; Wilkinson, S.; Sanchez-Osorio, J.; Cuello, C.; Parrilla, I.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. Early Developing Pig Embryos Mediate Their Own Environment in the Maternal Tract. PLoS ONE 2012, 7, e33625. [Google Scholar] [CrossRef] [Green Version]
- Schmaltzpanneau, B.; Cordova, A.; Dhorne-Pollet, S.; Hennequet-Antier, C.; Uzbekova, S.; Martinot, E.; Doret, S.; Martin, P.; Mermillod, P.; Locatelli, Y. Early Bovine Embryos Regulate Oviduct Epithelial Cell Gene Expression During In Vitro Co-Culture. Anim. Reprod. Sci. 2014, 149, 103–116. [Google Scholar] [CrossRef]
- Rizos, D.; Ramírez, M.; Pintado, B.; Lonergan, P.; Gutiérrez-Adán, A. Culture of Bovine Embryos in Intermediate Host Oviducts with Emphasis on the Isolated Mouse Oviduct. Theriogenology 2010, 73, 777–785. [Google Scholar] [CrossRef]
- Talukder, A.K.; Rashid, M.B.; Yousef, M.S.; Kusama, K.; Shimizu, T.; Shimada, M.; Suarez, S.S.; Imakawa, K.; Miyamoto, A. Oviduct Epithelium Induces Interferon-Tau in Bovine Day-4 Embryos, Which Generates an Anti-Inflammatory Response in Immune Cells. Sci. Rep. 2018, 8, 7850. [Google Scholar] [CrossRef]
- Dimou, E.; Nickel, W. Unconventional Mechanisms of Eukaryotic Protein Secretion. Curr. Biol. 2018, 28, R406–R410. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stahl, P.D. Extracellular Vesicles: A New Communication Paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Gwatkin, R. Effect of Viruses on Early Mammalian Development: III. Further Studies Concerning the Interaction of Mengo Encephalitis Virus with Mouse Ova. Fertil. Steril. 1966, 17, 411–420. [Google Scholar] [CrossRef]
- Kan, F.W.; Roux, E.; Bleau, G. Immunolocalization of Oviductin in Endocytic Compartments in the Blastomeres of Developing Embryos in the Golden Hamster1. Biol. Reprod. 1993, 48, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44. [Google Scholar]
- Banliat, C.; Tsikis, G.; Labas, V.; Teixeira-Gomes, A.-P.; Com, E.; Lavigne, R.; Pineau, C.; Guyonnet, B.; Mermillod, P.; Saint-Dizier, M. Identification of 56 Proteins Involved in Embryo–Maternal Interactions in the Bovine Oviduct. Int. J. Mol. Sci. 2020, 21, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-L.; Lee, K.-F.; Xu, J.-S.; He, Q.-Y.; Chiu, J.-F.; Lee, W.M.; Luk, J.M.; Yeung, W.S.B. The Embryotrophic Activity of Oviductal Cell-Derived Complement C3b and iC3b, a Novel Function of Complement Protein in Reproduction. J. Biol. Chem. 2004, 279, 12763–12768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiou, A.S.; Snijders, A.P.L.; Sostaric, E.; Aflatoonian, R.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Wright, P.C.; Fazeli, A. Modulation of The Oviductal Environment by Gametes. J. Proteome Res. 2007, 6, 4656–4666. [Google Scholar] [CrossRef] [PubMed]
- Klein, C. Novel Equine conceptus–endometrial Interactions on Day 16 of Pregnancy Based on RNA Sequencing. Reprod. Fertil. Dev. 2016, 28, 1712–1720. [Google Scholar] [CrossRef]
- Ward, P.P.; Mendoza-Meneses, M.; Mulac-Jericevic, B.; Cunningham, G.A.; Saucedo-Cardenas, O.; Teng, C.T.; Conneely, O.M. Restricted Spatiotemporal Expression of Lactoferrin During Murine Embryonic Development. Endocrinology 1999, 140, 1852–1860. [Google Scholar] [CrossRef]
- Østrup, O.; Olbricht, G.; Østrup, E.; Hyttel, P.; Collas, P.; Cabot, R. RNA Profiles of Porcine Embryos During Genome Activation Reveal Complex Metabolic Switch Sensitive to In Vitro Conditions. PLoS ONE 2013, 8, e61547. [Google Scholar] [CrossRef] [Green Version]
- Soleilhavoup, C.; Riou, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.L.; Reynaud, K.; De Graaf, S.P.; Gerard, N.; Druart, X. Proteomes of the Female Genital Tract During the Oestrous Cycle. Mol. Cell. Proteom. 2016, 15, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Larson, R.C.; Ignotz, G.G.; Currie, W.B. Effect of Fibronectin on Early Embryo Development in Cows. Reproduction 1992, 96, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafrenie, R.M.; Yamada, K.M. Integrin-Dependent Signal Transduction. J. Cell. Biochem. 1996, 61, 543–553. [Google Scholar] [CrossRef]
- Gabler, C.; Chapman, D.A.; Killian, G.J. Expression and Presence of Osteopontin and Integrins in the Bovine Oviduct During the Oestrous Cycle. Reproduction 2003, 126, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R.; Teng, T.; Beckman, W.C., Jr.; Jefferson, W.N.; Hanson, R.B.; Miller, J.V.; Mclachlan, J.A. Fluctuations of Lactoferrin Protein and Messenger Ribonucleic Acid in the Reproductive Tract of the Mouse During the Estrous Cycle. Biol. Reprod. 1992, 47, 903–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumoffen, C.M.; Gil, R.; Caille, A.M.; Morente, C.; Munuce, M.J.; Ghersevich, S.A. A Protein Isolated from Human Oviductal Tissue in Vitro Secretion, Identified as Human Lactoferrin, Interacts with Spermatozoa and Oocytes and Modulates Gamete Interaction. Hum. Reprod. 2013, 28, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, P.P.; Mendoza-Meneses, M.; Cunningham, G.A.; Conneely, O.M. Iron Status in Mice Carrying a Targeted Disruption of Lactoferrin. Mol. Cell. Biol. 2003, 23, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Buhi, W.; Alvarez, I. Identification, Characterization and Localization of Three Proteins Expressed by the Porcine Oviduct. Theriogenology 2003, 60, 225–238. [Google Scholar] [CrossRef]
- Georgiou, A.; Gil, M.; Almiñana, C.; Cuello, C.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Fazeli, A. Effects of Complement Component 3 Derivatives on Pig Oocyte Maturation, Fertilization and Early Embryo Development In Vitro. Reprod. Domest. Anim. 2011, 46, 1017–1021. [Google Scholar] [CrossRef]
- Algarra, B.; Han, L.; Soriano-Úbeda, C.; Avilés, M.; Coy, P.; Jovine, L.; Jiménez-Movilla, M. The C-Terminal Region of OVGP1 Remodels the Zona Pellucida and Modifies Fertility Parameters. Sci. Rep. 2016, 6, 32556. [Google Scholar] [CrossRef] [Green Version]
- Oliphant, G.; Bowling, A.; Eng, L.A.; Keen, S.; Randall, P.A. The Permeability of Rabbit Oviduct to Proteins Present in the Serum. Biol. Reprod. 1978, 18, 516–520. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Alonso, B.; Maillo, V.; Acuña, O.S.; López-Úbeda, R.; Torrecillas, A.; Simintiras, C.A.; Sturmey, R.; Avilés, M.; Lonergan, P.; Rizos, D. Spatial and Pregnancy-Related Changes in the Protein, Amino Acid, and Carbohydrate Composition of Bovine Oviduct Fluid. Int. J. Mol. Sci. 2020, 21, 1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliphant, G.; Reynolds, A.B.; Smith, P.F.; Ross, P.R.; Marta, J.S. Immunocytochemical Localization and Determination of Hormone-Induced Synthesis of the Sulfated Oviductal Glycoproteins. Biol. Reprod. 1984, 31, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.R.; Cheng, W.K.T. Changes in Composition of the Porcine Zona-Pellucida During Development of the Oocyte to the 2-Cell to 4-Cell Embryo. J. Embryol. Exp. Morphol. 1986, 92, 183–191. [Google Scholar]
- Oikawa, T.; Sendai, Y.; Kurata, S.-I.; Yanagimachi, R. A Glycoprotein of Oviductal Origin Alters Biochemical Properties of the Zona Pellucida of Hamster Egg. Gamete Res. 1988, 19, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Buhi, W.C. Characterization and Biological Roles of Oviduct-Specific, Oestrogen-Dependent Glycoprotein. Reproduction 2002, 123, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Nohara, M.; Yoshida-Komiya, H.; Kuramochi, T.; Ito, M.; Hoshi, H.; Shinkai, Y.; Sendai, Y. Effect of a Null Mutation of the Oviduct-Specific Glycoprotein Gene on Mouse Fertilization. Biochem. J. 2003, 374, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.K.; DeSouza, M.M. Messenger RNA Encoding an Estrogen-Dependent Oviduct Secretory Protein in the Sheep Is Localized in the Apical Tips and Basal Compartments of Fimbria and Ampulla Epithelial Cells Implying Translation at Unique Cytoplasmic Foci. Mol. Reprod. Dev. 1995, 42, 268–283. [Google Scholar] [CrossRef]
- Kan, F.W.; St-Jacques, S.; Bleau, G. Immunoelectron Microscopic Localization of an Oviductal Antigen in Hamster Zona Pellucida by Use of a Monoclonal Antibody. J. Histochem. Cytochem. 1988, 36, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Abé, H.; Oikawa, T. Immunocytochemical Localization of an Oviductal Zona Pellucida Glycoprotein in the Oviductal Epithelium of the Golden Hamster. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1991, 229, 305–314. [Google Scholar]
- McBride, D.S.; Boisvert, C.; Bleau, G.; Kan, F.W. Detection of Nascent and/Or Mature Forms of Oviductin in the Female Reproductive Tract and Post-Ovulatory Oocytes by Use of a Polyclonal Antibody Against Recombinant Hamster Oviductin. J. Histochem. Cytochem. 2004, 52, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Boice, M.L.; Geisert, R.D.; Blair, R.M.; Verhage, H.G. Identification and Characterization of Bovine Oviductal Glycoproteins Synthesized at Estrus. Biol. Reprod. 1990, 43, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Abé, H.; Numazawa, C.; Abe, M.; Onodera, M.; Katsumi, A. Immunocytochemical Localization of Oviduct-Specific Glycoproteins in the Oviductal Epithelium from Cows at Follicular and Luteal Phases. Cell Tissue Res. 1993, 274, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Buhi, W.C.; O’Brien, B.; Alvarez, I.M.; Erdos, G.; Dubois, D. Immunogold Localization of Porcine Oviductal Secretory Proteins Within the Zona Pellucida, Perivitelline Space, and Plasma Membrane of Oviductal and Uterine Oocytes and Early Embryos1. Biol. Reprod. 1993, 48, 1274–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhage, H.G.; Mavrogianis, P.A.; Boice, M.L.; Li, W.; Fazleabas, A.T. Oviductal Epithelium of the Baboon: Hormonal Control and the Immuno-Gold Localization of Oviduct-Specific Glycoproteins. Am. J. Anat. 1990, 187, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Rapisarda, J.J.; Mavrogianis, P.A.; O’Day-Bowman, M.B.; Fazleabas, A.T.; Verhage, H.G. Immunological Characterization and Immunocytochemical Localization of an Oviduct-Specific Glycoprotein in the Human. J. Clin. Endocrinol. Metab. 1993, 76, 1483–1488. [Google Scholar] [PubMed]
- Kapur, R.P.; Johnson, L.V. Ultrastructural Evidence That Specialized Regions of the Murine Oviduct Contribute a Glycoprotein to the Extracellular Matrix of Mouse Oocytes. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1988, 221, 720–729. [Google Scholar] [CrossRef]
- Arias, E.B.; Verhage, H.G.; Jaffe, R.C. Complementary Deoxyribonucleic Acid Cloning and Molecular Characterization of an Estrogen-Dependent Human Oviductal Glycoprotein1. Biol. Reprod. 1994, 51, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Pascal, G.; Fouchécourt, S.; Pontarotti, P.; Monget, P. Gene Birth, Death, and Divergence: The Different Scenarios of Reproduction-Related Gene Evolution1. Biol. Reprod. 2009, 80, 616–621. [Google Scholar] [CrossRef] [Green Version]
- Moros-Nicolás, C.; Fouchécourt, S.; Goudet, G.; Monget, P. Genes Encoding Mammalian oviductal proteins involved in fertilization are subjected to gene death and positive selection. J. Mol. Evol. 2018, 86, 655–667. [Google Scholar] [CrossRef]
- Fazleabas, A.T.; Verhage, H.G. The Detection of Oviduct-Specific Proteins in the Baboon (Papio anubis). Biol. Reprod. 1986, 35, 455–462. [Google Scholar] [CrossRef]
- Verhage, H.G.; Fazleabas, A.T.; Donnelly, K. The in vitro synthesis and release of proteins by the human oviduct. Endocrinology 1988, 122, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.M.; Fazleabas, A.T.; Verhage, H.G.; Mavrogianis, P.A.; Jaffe, R.C. Cloning of a Recombinant Complementary DNA to a Baboon (Papio Anubis) Estradiol-Dependent Oviduct-Specific Glycoprotein. Mol. Endocrinol. 1991, 5, 356–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Day-Bowman, M.B.; Mavrogianis, P.A.; Reuter, L.M.; Johnson, D.E.; Fazleabas, A.T.; Verhage, H.G. Association of oviduct-specific glycoproteins with human and baboon (Papio anubis) ovarian oocytes and enhancement of human sperm binding to human hemizonae following in vitro incubation. Biol. Reprod. 1996, 54, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Verhage, H.G.; Fazleabas, A.T.; Mavrogianis, P.A.; O’Day-Bowman, M.B.; Donnelly, K.M.; Jaffe, R.C. The baboon oviduct: Characteristics of an oestradiol-dependent oviduct-specific glycoprotein. Hum. Reprod. Update 1997, 3, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachen, A.; Jewgenow, K.; Braun, B.C. Sequence analysis of feline oviductin and its expression during the estrous cycle in the domestic cat (Felis catus). Theriogenology 2012, 77, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Hribal, R.; Hachen, A.; Zahmel, J.; Gonzalez, L.F.; Jewgenow, K.; Braun, B.C. The effect of recombinant feline oviductin used in the cat IVF system. Reprod. Domest. Anim. 2014, 49, 24–25. [Google Scholar]
- Hribal, R.; Hachen, A.; Jewgenow, K.; Zahmel, J.; Fernandez-Gonzalez, L.; Braun, B.C. The influence of recombinant feline oviductin on different aspects of domestic cat (Felis catus) IVF and embryo quality. Theriogenology 2014, 82, 742–749. [Google Scholar] [CrossRef]
- Kim, R.N.; Kim, D.W.; Choi, S.H.; Chae, S.H.; Nam, S.H.; Kim, A.; Kang, A.; Park, K.H.; Lee, Y.S.; Hirai, M.; et al. Major chimpanzee-specific structural changes in sperm development-associated genes. Funct. Integr. Genom. 2011, 11, 507–517. [Google Scholar] [CrossRef]
- Malayer, J.; Hansen, P.; Buhi, W. Secretion of proteins by cultured bovine oviducts collected from estrus through early diestrus. J. Exp. Zool. 1988, 248, 345–353. [Google Scholar] [CrossRef]
- King, R.S.; Anderson, S.H.; Killian, G.J. Effect of bovine oviductal estrus-associated protein on the ability of sperm to capacitate and fertilize oocytes. J. Androl. 1994, 15, 468–478. [Google Scholar]
- Abe, H.; Sendai, Y.; Satoh, T.; Hoshi, H. Bovine oviduct-specific glycoprotein: A potent factor for maintenance of viability and motility of bovine spermatozoa in vitro. Mol. Reprod. Dev. 1995, 42, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Martus, N.S.; Verhage, H.G.; Mavrogianis, P.A.; Thibodeaux, J.K. Enhancement of bovine oocyte fertilization in vitro with a bovine oviductal specific glycoprotein. J. Reprod. Fertil. 1998, 113, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Dizier, M.; Marnier, C.; Tahir, M.Z.; Grimard, B.; Thoumire, S.; Chastant-Maillard, S.; Reynaud, K. OVGP1 Is Expressed in the Canine Oviduct at the Time and Place of Oocyte Maturation and Fertilization. Mol. Reprod. Dev. 2014, 81, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Onodera, M.; Sugawara, S. Immunological detection and characterization of an estrus-associated antigen in the goat oviduct. J. Exp. Zool. 1995, 272, 134–141. [Google Scholar] [CrossRef]
- Pradeep, M.A.; Jagadeesh, J.; De, A.K.; Kaushik, J.K.; Malakar, D.; Kumar, S.; Dang, A.K.; Das, S.K.; Mohanty, A.K. Purification, sequence characterization and effect of goat oviduct-specific glycoprotein on in vitro embryo development. Theriogenology 2011, 75, 1005–1015. [Google Scholar] [CrossRef]
- Léveillé, M.-C.; Roberts, K.D.; Chevalier, S.; Chapdelaine, A.; Bleau, G. Uptake of an oviductal antigen by the hamster zona pellucida. Biol. Reprod. 1987, 36, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Robitaille, G.; St-Jacques, S.; Potier, M.; Bleau, G. Characterization of an oviductal glycoprotein associated with the ovulated hamster oocyte. Biol. Reprod. 1988, 38, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Kan, F.W.; Roux, E.; St.-Jacquesz, S.; Bleau, G. Demonstration by lectin-gold cytochemistry of transfer of glycoconjugates of oviductal origin to the zona pellucida of oocytes after ovulation in hamsters. Anat. Rec. 1990, 226, 37–47. [Google Scholar] [CrossRef]
- Suzuki, K.; Sendai, Y.; Onuma, T.; Hoshi, H.; Hiroi, M.; Araki, Y. Molecular characterization of a hamster oviduct-specific glycoprotein. Biol. Reprod. 1995, 53, 345–354. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Mavrogianis, P.A.; O’dayBowman, M.B.; Jaffe, R.C.; Verhage, H.G. Characterization of antibodies generated against a conserved portion of oviductal glycoprotein (OGP) and endogenous hamster OGP and their ability to decrease sperm binding to the zona pellucida in vitro. Am. J. Reprod. Immunol. 1997, 38, 377–383. [Google Scholar] [CrossRef]
- Abe, H.; Satoh, T.; Hoshi, H. Primary modulation by oestradiol of the production of an oviduct-specific glycoprotein by the epithelial cells in the oviduct of newborn golden hamsters. J. Reprod. Fertil. 1998, 112, 157–163. [Google Scholar] [CrossRef] [PubMed]
- O’Day-Bowman, M.B.; Mavrogianis, P.A.; Minshall, R.D.; Verhage, H.G. In vivo versus in vitro oviductal glycoprotein (OGP) association with the zona pellucida (ZP) in the hamster and baboon. Mol. Reprod. Dev. 2002, 62, 248–256. [Google Scholar] [CrossRef] [PubMed]
- O’day-Bowman, M.B.; Mavrogianis, P.A.; Fazleabas, A.T.; Verhage, H.G. A human oviduct-specific glycoprotein: Synthesis, secretion, and localization during the menstrual cycle. Microsc. Res. Tech. 1995, 32, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Briton-Jones, C.; Lok, I.H.; Yuen, P.M.; Chiu, T.T.Y.; Cheung, L.P.; Haines, C. Regulation of human oviductin mRNA expression in vivo. Fertil. Steril. 2001, 75, 942–946. [Google Scholar] [CrossRef]
- Briton-Jones, C.; Lok, I.H.; Cheung, C.K.; Chiu, T.T.Y.; Cheung, L.P.; Haines, C. Estradiol regulation of oviductin/oviduct-specific glycoprotein messenger ribonucleic acid expression in human oviduct mucosal cells in vitro. Fertil. Steril. 2004, 81, 749–756. [Google Scholar] [CrossRef]
- Verhage, H.G.; Mavrogianis, P.A.; Boomsma, R.A.; Schmidt, A.; Brenner, R.M.; Slayden, O.V.; Jaffe, R.C. Immunologic and molecular characterization of an estrogen-dependent glycoprotein in the rhesus (Macaca mulatta) oviduct. Biol. Reprod. 1997, 57, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.M.; Zhang, G.J.; Fang, X.D.; Zhang, Y.F.; Li, C.; Ling, F.; Cooper, D.N.; Li, Q.Y.; Li, Y.; van Gool, A.J.; et al. Genome sequencing and comparison of two nonhuman primate animal models, the cynomolgus and Chinese rhesus macaques. Nat. Biotechnol. 2011, 29, 1019–U1089. [Google Scholar] [CrossRef] [Green Version]
- Kapur, R.P.; Johnson, L.V. An oviductal fluid glycoprotein associated with ovulated mouse ova and early embryos. Dev. Biol. 1985, 112, 89–93. [Google Scholar] [CrossRef]
- Sendai, Y.; Komiya, H.; Suzuki, K.; Onuma, T.; Kikuchi, M.; Hoshi, H.; Araki, Y. Molecular cloning and characterization of a mouse oviduct-specific glycoprotein. Biol. Reprod. 1995, 53, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Buhi, W.; Vallet, J.; Bazer, F. De novo synthesis and release of polypeptides from cyclic and early pregnant porcine oviductal tissue in explant culture. J. Exp. Zool. 1989, 252, 79–88. [Google Scholar] [CrossRef]
- Buhi, W.; Alvarez, I.; Sudhipong, V.; Dones-Smith, M. Identification and characterization of de novo-synthesized porcine oviductal secretory proteins. Biol. Reprod. 1990, 43, 929–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhi, W.C.; Alvarez, I.M.; Choi, I.; Cleaver, B.D.; Simmen, F.A. Molecular cloning and characterization of an estrogen-dependent porcine oviductal secretory glycoprotein. Biol. Reprod. 1996, 55, 1305–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouba, A.J.; Abeydeera, L.R.; Alvarez, I.M.; Day, B.N.; Buhi, W.C. Effects of the porcine oviduct-specific glycoprotein on fertilization, polyspermy, and embryonic development in vitro. Biol. Reprod. 2000, 63, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merchan, M.; Rendon, M.; Folch, J.M. Assignment of the oviductal glycoprotein 1 gene (OVGP1) to porcine chromosome 4q22 -> q23 by radiation hybrid panel mapping. Cytogenet. Genome Res. 2006, 114, 93C. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, G.; Ross, P.R. Demonstration of production and isolation of three sulfated glycoproteins from the rabbit oviduct. Biol. Reprod. 1982, 26, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Merchan, M.; Peiro, R.; Argente, M.J.; Santacreu, M.A.; Garcia, M.L.; Blasco, A.; Folch, J.M. Analysis of the oviductal glycoprotein 1 polymorphisms and their effects on components of litter size in rabbits. Anim. Genet. 2009, 40, 756–758. [Google Scholar] [CrossRef]
- Garcia, M.L.; Peiro, R.; Argente, M.J.; Merchan, M.; Folch, J.M.; Blasco, A.; Santacreu, M.A. Investigation of the oviductal glycoprotein 1 (OVGP1) gene associated with embryo survival and development in the rabbit. J. Anim. Sci. 2010, 88, 1597–1602. [Google Scholar] [CrossRef] [Green Version]
- Sutton, R.; Nancarrow, C.; Wallace, A.; Rigby, N. Identification of an oestrus-associated glycoprotein in oviducal fluid of the sheep. J. Reprod. Fertil. 1984, 72, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Gandolfi, F.; Brevini, T.; Richardson, L.; Brown, C.; Moor, R. Characterization of proteins secreted by sheep oviduct epithelial cells and their function in embryonic development. Development 1989, 106, 303–312. [Google Scholar]
- Gandolfi, F.; Modina, S.; Brevini, T.; Galli, C.; Moor, R.; Lauria, A. Oviduct ampullary epithelium contributes a glycoprotein to the zona pellucida, perivitelline space and blastomeres membrane of sheep embryos. Eur. J. Basic Appl. Histochem. 1991, 35, 383–392. [Google Scholar]
- DeSouza, M.M.; Murray, M.K. An estrogen-dependent secretory protein, which shares identity with chitinases, is expressed in a temporally and regionally specific manner in the sheep oviduct at the time of fertilization and embryo development. Endocrinology 1995, 136, 2485–2496. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.L.; Wade, M.G.; Nancarrow, C.D.; Kelleher, D.L.; Boland, M.P. Influence of ovine oviducal amino acid concentrations and an ovine oestrus-associated glycoprotein on development and viability of bovine embryos. Mol. Reprod. Dev. 1997, 47, 164–169. [Google Scholar] [CrossRef]
- Malette, B.; Paquette, Y.; Merlen, Y.; Bleau, G. Oviductins possess chitinase-and mucin-like domains: A lead in the search for the biological function of these oviduct-specific ZP-associating glycoproteins. Mol. Reprod. Dev. 1995, 41, 384–397. [Google Scholar] [CrossRef]
- Laheri, S.; Ashary, N.; Bhatt, P.; Modi, D. Oviductal glycoprotein 1 (OVGP1) is expressed by endometrial epithelium that regulates receptivity and trophoblast adhesion. J. Assist. Reprod. Genet. 2018, 35, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Buhi, W.; Bazer, F.; Alvarez, I.; Mirandot, M. In vitro synthesis of oviductal proteins associated with estrus and 17β-estradiol-treated ovariectomized ewes. Endocrinology 1991, 128, 3086–3095. [Google Scholar] [CrossRef]
- Sendai, Y.; Abe, H.; Kikuchi, M.L.; Satoh, T.; Hoshi, H. Purification and molecular cloning of bovine oviduct-specific glycoprotein. Biol. Reprod. 1994, 50, 927–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nancarrow, C.; Hill, J. Oviduct proteins in fertilization and early embryo development. J. Reprod. Fertil. Suppl. Only 1995, 49, 3–14. [Google Scholar] [CrossRef]
- Jaffe, R.C.; Arias, E.B.; OdayBowman, M.B.; Donnelly, K.M.; Mavrogianis, P.A.; Verhage, H.G. Regional distribution and hormonal control of estrogen-dependent oviduct-specific glycoprotein messenger ribonucleic acid in the baboon (Papio anubis). Biol. Reprod. 1996, 55, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Buhi, W.C.; Alvarez, I.M.; Kouba, A.J. Secreted proteins of the oviduct. Cells Tissues Organs 2000, 166, 165–179. [Google Scholar] [CrossRef]
- Lok, I.H.; Briton-Jones, C.M.; Yuen, P.M.; Haines, C.J. Variable expression of oviductin mRNA at different stages of human reproductive cycle. J. Assist. Reprod. Genet. 2002, 19, 569–576. [Google Scholar] [CrossRef]
- Laheri, S.; Modi, D.; Bhatt, P. Extra-oviductal expression of oviductal glycoprotein 1 in mouse: Detection in testis, epididymis and ovary. J. Biosci. 2017, 42, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Paquette, Y.; Merlen, Y.; Malette, B.; Bleau, G. Allelic polymorphism in the hamster oviductin gene is due to a variable number of mucin-like tandem repeats. Mol. Reprod. Dev. 1995, 42, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Merchan, M.; Peiro, R.; Santacreu, M.A.; Francino, O.; Folch, J.M. Rabbit oviductal glycoprotein 1 gene: Genomic organization polymorphism analysis and mRNA expression. Mol. Reprod. Dev. 2007, 74, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Komiya, H.; Onuma, T.; Hiroi, M.; Araki, Y. In situ localization of messenger ribonucleic acid for an oviduct-specific glycoprotein during various hormonal conditions in the golden hamster. Biol. Reprod. 1996, 55, 1107–1118. [Google Scholar] [CrossRef]
- Roux, E.; Kan, F.W. Stage-specific immunolabeling for oviductin in the secretory granules of the oviductal epithelium of the golden hamster during the estrous cycle. Anat. Rec. 1995, 241, 369–376. [Google Scholar] [CrossRef]
- Buhi, W.; Alvarez, I.; Kouba, A. Oviductal regulation of fertilization and early embryonic development. J. Reprod. Fertil. Suppl. 1997, 52, 285. [Google Scholar]
- Murray, M.K. An estrogen-dependent glycoprotein is synthesized and released from the oviduct in a temporal-and region-specific manner during early pregnancy in the ewe. Biol. Reprod. 1993, 48, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Hyde, B.; Black, D. Synthesis and secretion of sulphated glycoproteins by rabbit oviduct explants in vitro. Reproduction 1986, 78, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Erickson-Lawrence, M.F.; Turner, T.T.; Thomas, T.S.; Oliphant, G. Effect of steroid hormones on sulfated oviductal glycoprotein secretion by oviductal explants in vitro. Biol. Reprod. 1989, 40, 1311–1319. [Google Scholar] [CrossRef]
- Gonella-Diaza, A.M.; da Silva Andrade, S.C.; Sponchiado, M.; Pugliesi, G.; Mesquita, F.S.; Van Hoeck, V.; de Francisco Strefezzi, R.; Gasparin, G.R.; Coutinho, L.L.; Binelli, M. Size of the ovulatory follicle dictates spatial differences in the oviductal transcriptome in cattle. PLoS ONE 2015, 10, e0145321. [Google Scholar] [CrossRef] [Green Version]
- Kadam, K.M.; D’Souza, S.J.; Bandivdekar, A.H.; Natraj, U. Identification and characterization of oviductal glycoprotein-binding protein partner on gametes: Epitopic similarity to non-muscle myosin IIA, MYH 9. Mol. Hum. Reprod. 2006, 12, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSouza, M.M.; Murray, M.K. An estrogen-dependent sheep oviductal glycoprotein has glycan linkages typical of sialomucins and does not contain chitinase activity. Biol. Reprod. 1995, 53, 1517–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, S.; Janjanam, J.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K. Structural and functional characterization of buffalo oviduct-specific glycoprotein (OVGP1) expressed during estrous cycle. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadam, K.M.; D’Souza, S.J.; Natraj, U. Identification of cellular isoform of oviduct-specific glycoprotein: Role in oviduct tissue remodeling? Cell Tissue Res. 2007, 330, 545–556. [Google Scholar] [CrossRef]
- Wegner, C.C.; Killian, G.J. In vitro and in vivo association of an oviduct estrus-associated protein with bovine zona pellucida. Mol. Reprod. Dev. 1991, 29, 77–84. [Google Scholar] [CrossRef]
- Coy, P.; Canovas, S.; Mondejar, I.; Saavedra, M.D.; Romar, R.; Grullon, L.; Matas, C.; Aviles, M. Oviduct-specific glycoprotein and heparin modulate sperm-zona pellucida interaction during fertilization and contribute to the control of polyspermy. Proc. Natl. Acad. Sci. USA 2008, 105, 15809–15814. [Google Scholar] [CrossRef] [Green Version]
- Algarra, B.; Maillo, V.; Avilés, M.; Gutiérrez-Adán, A.; Rizos, D.; Jiménez-Movilla, M. Effects of recombinant OVGP1 protein on in vitro bovine embryo development. J. Reprod Dev. 2018, 64, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Kapur, R.P.; Johnson, L.V. Selective sequestration of an oviductal fluid glycoprotein in the perivitelline space of mouse oocytes and embryos. J. Exp. Zool. 1986, 238, 249–260. [Google Scholar] [CrossRef]
- Lyng, R.; Shur, B.D. Mouse oviduct-specific glycoprotein is an egg-associated ZP3-independent sperm-adhesion ligand. J. Cell Sci. 2009, 122, 3894–3906. [Google Scholar] [CrossRef] [Green Version]
- Boice, M.L.; McCarthy, T.J.; Mavrogianis, P.A.; Fazlebas, A.T.; Verhage, H.G. Localization of Oviductal Glycoproteins within the Zona Pellucida and Perivitelline Space of Ovulated Ova and Early Embryos in Baboons (Paplo Anubis). Biol. Reprod. 1990, 43, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Reuter, L.M.; O’Day-Bowman, M.B.; Mavrogianis, P.A.; Fazleabas, A.T.; Verhage, H.G. In vitro incubation of golden (Syrian) hamster ovarian oocytes and human sperm with a human oviduct specific glycoprotein. Mol. Reprod. Dev. 1994, 38, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Malette, B.; Bleau, G. Biochemical characterization of hamster oviductin as a sulphated zona pellucida-binding glycoprotein. Biochem. J. 1993, 295, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.J.; Zhao, Y.W.; Yang, X.L.; Kan, F.W.K. Recombinant Hamster Oviductin Is Biologically Active and Exerts Positive Effects on Sperm Functions and Sperm-Oocyte Binding. PLoS ONE 2015, 10, e0123003. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, R.E.; Romar, R.; Matas, C.; Gutierrez-Adan, A.; Holt, W.V.; Coy, P. Effects of oviductal fluid on the development, quality, and gene expression of porcine blastocysts produced in vitro. Reproduction 2009, 137, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, R.F.; Staros, A.L.; Killian, G.J. Oviductal Fluid Proteins Associated with the Bovine Zona Pellucida and the Effect on In Vitro Sperm-Egg Binding, Fertilization and Embryo Development. Reprod. Domest. Anim. 2008, 43, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Broermann, D.; Xie, S.; Nephew, K.; Pope, W. Effects of the oviduct and wheat germ agglutinin on enzymatic digestion of porcine zona pellucidae. J. Anim. Sci. 1989, 67, 1324–1329. [Google Scholar] [CrossRef] [PubMed]
- Davachi, N.D.; Shahneh, A.Z.; Kohram, H.; Zhandi, M.; Shamsi, H.; Hajiyavand, A.M.; Saadat, M. Differential influence of ampullary and isthmic derived epithelial cells on zona pellucida hardening and in vitro fertilization in ovine. Reprod. Biol. 2016, 16, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.H.; Funahashi, H.; Abeydeera, L.R.; Moon, S.J.; Prather, R.S.; Day, B.N. Effects of oviductal fluid on sperm penetration and cortical granule exocytosis during fertilization of pig oocytes in vitro. J. Reprod. Fertil. 1996, 107, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolbe, T.; Holtz, W. Differences in proteinase digestibility of the zona pellucida of in vivo and in vitro derived porcine oocytes and embryos. Theriogenology 2005, 63, 1695–1705. [Google Scholar] [CrossRef]
- Mondejar, I.; Aviles, M.; Coy, P. The human is an exception to the evolutionarily-conserved phenomenon of pre-fertilization zona pellucida resistance to proteolysis induced by oviductal fluid. Hum. Reprod. 2013, 28, 718–728. [Google Scholar] [CrossRef] [Green Version]
- Mondejar, I.; Martinez-Martinez, I.; Aviles, M.; Coy, P. Identification of Potential Oviductal Factors Responsible for Zona Pellucida Hardening and Monospermy During Fertilization in Mammals. Biol. Reprod. 2013, 89, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Abeydeera, L.R.; Prather, R.S.; Day, B.N. Morphologic comparison of ovulated and in vitro-matured porcine oocytes, with particular reference to polyspermy after in vitro fertilization. Mol. Reprod. Dev. 1998, 49, 308–316. [Google Scholar] [CrossRef]
- Wright Jr, R.W.; Cupps, P.T.; Gaskins, C.T.; Hillers, J.K. Comparative solubility properties of the zona pellucidae of unfertilized murine, ovine and bovine ova. J. Anim. Sci. 1977, 44, 850–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCauley, T.C.; Buhi, W.C.; Wu, G.M.; Mao, J.; Caamano, J.N.; Didion, B.A.; Day, B.N. Oviduct-specific glycoprotein modulates sperm-zona binding and improves efficiency of porcine fertilization in vitro. Biol. Reprod. 2003, 69, 828–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Mavrogianis, P.A.; OdayBowman, M.B.; Verhage, H.G. Species-specific effect of oviductal glycoproteins on hamster sperm binding to hamster oocytes. Mol. Reprod. Dev. 1997, 46, 201–207. [Google Scholar] [CrossRef]
- Boatman, D.E.; Magnoni, G.E. Identification of a sperm penetration factor in the oviduct of the golden hamster. Biol. Reprod. 1995, 52, 199–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, P.; Gu, Z.; Luo, J.P.; Wang, J.R.; Tso, J.K. Antibodies against the C-terminal peptide of rabbit oviductin inhibit mouse early embryo development to pass 2-cell stage. Cell Res. 2002, 12, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.; Walker, S.; Brown, G.; Nancarrow, C. The effects of an estrus-associated oviductal glycoprotein on the in vitro fertilization and development of ovine oocytes matured in vitro. Theriogenology 1996, 46, 1379–1388. [Google Scholar] [CrossRef]
- Hill, J.; Walker, S.; Brown, G.; Nancarrow, C. The effects of an ovine oviductal estrus-associated glycoprotein on early embryo development. Theriogenology 1996, 46, 1367–1377. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HUMAN | MOUSE | PIG | COW | SHEEP | MARE | CAT | BITCH | GUINEA PIG | RABBIT | |
|---|---|---|---|---|---|---|---|---|---|---|
| 2 cells | 1 (in vitro) [28] | 1 [29] | 14–16 h [30] | 24–30 h [31] | 1.5 [32] | 20 h [33] | 2.5 [34] | 4.5–5.5 [35] | 2 [36] | 16–17 h [37] |
| 8 cells | 3 (in vitro) [38] | 2.5 [29] | 2 [30] | 3.5 [39] | 2 [32] | 3 [40] | 3 [34] | 4.5–5.5 [35] | 4 [36] | 29–30 h [37] |
| Morula | 4 (in vitro) [28] | 3 [29] | 3.5 [30] | 5–6 [31] | 4 [32] | 4 [40] | 5 [34] | 10 [41] | 5 [36] | 2.5 [42,43] |
| Blastocyst | 5 (in vitro) [28] | 3.5 [29] | 5 [30] | 7 [31] | 6 [44] | 6 [40] | 6–9 [34] | 12–13 [41] | 6 [36] | 3 [43] |
| Hatching | 6–7 (in vitro) [45] | 4.5 [46] | 6 [30] | 8 [31] | 8 [44] | 7–8 [40] | 12 [47] | 19–20 [48] | 6 [49] | 4 (in vitro) [37] no hatching until implantation in vivo [50] |
| Conceptus reaches the uterus | 3.5 [51] | 3 [29] | 2 [30] | 4 [31] | 4 [44] | 5.5–6.5 [40,52] | 5–6 [34] | 7–10 [41,53,54] | 4.5 [36] | 3.5 [42] |
| Conceptus reaches the uterus (stage) | Morula [51] | Morula [29,55] | 4–8 cells [30] | 16 cells [31] | Morula (16–32 cells) [44] | Blastocyst [40] | Compact morula or blastocyst [34] | Compact morula or blastocyst [56] | 8–16 cells [36,57] | Blastocyst [42] |
| Implantation | 6–10 [58] | 4.5 [55] | 14–18 [59] | 19 [31] | 16 [44] | ~40 [59] | 13–14 [60] | From 17–18 [61] | 6–7 [49] | 7.5 [42] |
| OF Ligands | Receptor in Blastomeres | Citations | ||
|---|---|---|---|---|
| Annexin A1 | ANXA1 | Epidermal growth factor receptor | EGFR | [98] |
| Apolipoprotein C-III | APOC3 | Syndecan 2 | SDC2 | |
| Complement component 3 | C3 | CD19 molecule | CD19 | [99,100] |
| CD81 molecule | CD81 | |||
| Calreticulin | CALR | Integrin, alpha V | ITGAV | |
| Fibrinogen gamma chain | FGC | Integrin, alpha V | ITGAV | [101] |
| Integrin, beta 1 (fibronectin receptor, beta polypeptide) | ITGB1 | |||
| Heat shock protein 90 kDa alpha (cytosolic), class A member 1 | GPI | Cystic fibrosis transmembrane conductance regulator | CFTR | |
| Epidermal growth factor receptor | EGFR | |||
| Lactotransferrin | LTF | Transferrin receptor | TFRC | [102] |
| Glucose-6-phosphate isomerase | GPI | Autocrine motility factor receptor | AMFR | |
| Species | Reference |
|---|---|
| Baboon (Papio anubis) | [134,135,136,137,138] |
| Cat (Felis catus) | [139,140,141] |
| Chimpanzee (Pan troglodytes) | [142] |
| Cow (Bos taurus) | [7,125,143,144,145,146] |
| Dog (Canis lupus familiaris) | [147] |
| Goat (Capra ibex) | [148,149] |
| Hamster (Mesocricetus auratus) | [118,124,150,151,152,153,154,155,156] |
| Human (Homo sapiens) | [129,131,157,158,159] |
| Macaque (Macaca mulatta) | [160,161] |
| Mouse (Mus musculus) | [120,162,163] |
| Pig (Sus scrofa) | [164,165,166,167,168] |
| Rabbit (Oryctolagus cuniculus) | [169,170,171] |
| Sheep (Ovis aries) | [172,173,174,175,176] |
| OVGP1 Source | In Vivo/In Vitro | Technique | ZP | Oocyte | Embryo | Reference | |
|---|---|---|---|---|---|---|---|
| Baboon (Papio anubis) | Baboon | in vivo/in vitro | OM | + | + (PVS) | 2–4 cells. + (ZP, PVS) | [137,156,204] |
| EM | + | + (PVS, PM, O) | + (ZP, PVS, BO) | ||||
| Human | in vitro | OM | + | / | / | [137] | |
| Human (Homo sapiens) | Human | in vitro | OM | + | / | / | [137] |
| Baboon | in vitro (Hemizonae) | OM | + | / | / | [137] | |
| Hamster (Mesocricetus auratus) | Human | in vitro | OM | + | + (PVS) | / | [205] |
| Hamster | in vivo/in vitro | OM | + | - | / | [96,124,150,151,154,156,205,206,207] | |
| EM | + | + (PVS, PM, OV) | 2–8 cells. + (ZP, E, L, MVB) | ||||
| WB | + | / | / | ||||
| Pig (Sus scrofa) | Pig | in vivo/in vitro | OM | + | + (ZP, PM, O) | / | [113,127,200] |
| EM | + | + (PVS, PM, MVB) | Day 2–7 + (ZP, PVS, PM, BM) | ||||
| MS | + | / | / | ||||
| Cow | in vitro | OM | + | / | / | [208] | |
| Cow (Bos taurus) | Cow | in vivo/in vitro | OM | + | - | Day 7 + (ZP) Day 4–6 and morula + (PVS, BC) | [98,199,209] |
| MS | / | / | + | ||||
| Pig | in vitro | OM | + | + (ZP) | + (ZP, PVS, BM, BC) | [201] | |
| Sheep (Ovis aries) | Sheep | in vivo/in vitro | OM/EM/WB | + | + (PVS) | + (ZP, PVS, BC) | [173,174] |
| Mouse (Mus musculus) | Mouse | in vivo/in vitro | OM | +/− | + (PVS) | 2 cells + (ZP, PVS) | [130,162,202,203] |
| EM | − | + (PVS) | / | ||||
| WB | +/− | +/− | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Brusi, L.; Algarra, B.; Moros-Nicolás, C.; Izquierdo-Rico, M.J.; Avilés, M.; Jiménez-Movilla, M. A Comparative View on the Oviductal Environment during the Periconception Period. Biomolecules 2020, 10, 1690. https://doi.org/10.3390/biom10121690
González-Brusi L, Algarra B, Moros-Nicolás C, Izquierdo-Rico MJ, Avilés M, Jiménez-Movilla M. A Comparative View on the Oviductal Environment during the Periconception Period. Biomolecules. 2020; 10(12):1690. https://doi.org/10.3390/biom10121690
Chicago/Turabian StyleGonzález-Brusi, Leopoldo, Blanca Algarra, Carla Moros-Nicolás, Mª José Izquierdo-Rico, Manuel Avilés, and Maria Jiménez-Movilla. 2020. "A Comparative View on the Oviductal Environment during the Periconception Period" Biomolecules 10, no. 12: 1690. https://doi.org/10.3390/biom10121690
APA StyleGonzález-Brusi, L., Algarra, B., Moros-Nicolás, C., Izquierdo-Rico, M. J., Avilés, M., & Jiménez-Movilla, M. (2020). A Comparative View on the Oviductal Environment during the Periconception Period. Biomolecules, 10(12), 1690. https://doi.org/10.3390/biom10121690