A Genetic Polymorphism in the Pannexin1 Gene Predisposes for The Development of Endothelial Dysfunction with Increasing BMI

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Blood Measurements
2.3. Assessment of Myocardial Perfusion with PET/CT
2.4. Phenotype–Genotype Association Studies
2.5. Statistical Analysis
3. Results
3.1. BMI Correlates with Cardiovascular Risk Factors
3.2. BMI, Hemodynamic Parameters, and Panx1-400A>C SNP
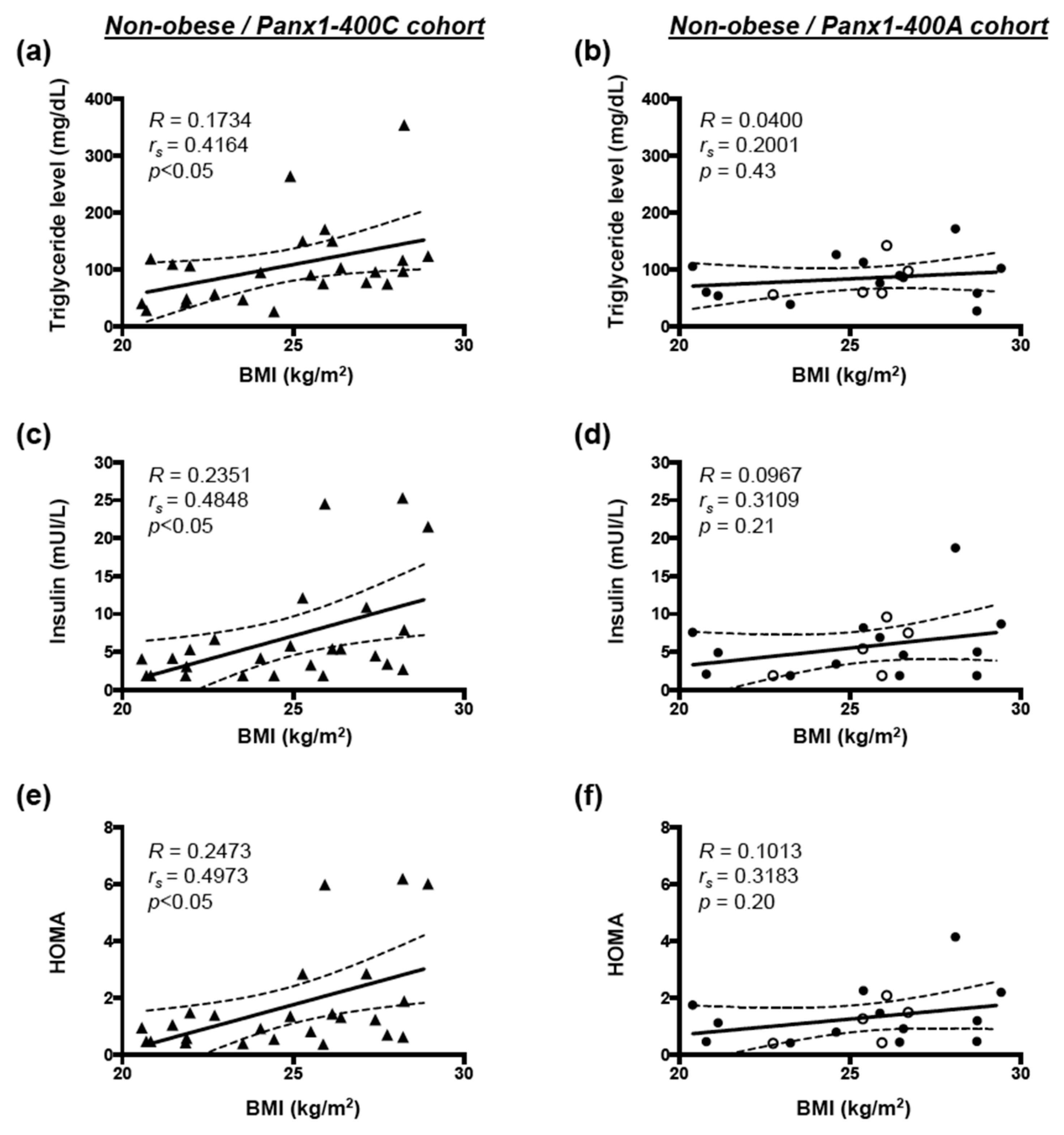
3.3. Correlation between BMI and Cardiovascular Risk Factors in Non-obese and Obese Subjects
3.4. Correlation Between Hemodynamic Parameters and Cardiovascular Risk Factors in Non-Obese Subjects
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silver, A.E.; Beske, S.D.; Christou, D.D.; Donato, A.J.; Moreau, K.L.; Eskurza, I.; Gates, P.E.; Seals, D.R. Overweight and obese humans demonstrate increased vascular endothelial NAD(P)H oxidase-p47(phox) expression and evidence of endothelial oxidative stress. Circulation 2007, 115, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Guns, P.J.; Van Assche, T.; Fransen, P.; Robaye, B.; Boeynaems, J.M.; Bult, H. Endothelium-dependent relaxation evoked by ATP and UTP in the aorta of P2Y2-deficient mice. Br. J. Pharmacol. 2006, 147, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, C.G.; Specht, A.; Wegiel, B.; Ferran, C.; Kaczmarek, E. Mechanism of purinergic activation of endothelial nitric oxide synthase in endothelial cells. Circulation 2009, 119, 871–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathanoori, R.; Sward, K.; Olde, B.; Erlinge, D. The ATP receptors P2X7 and P2X4 modulate high glucose and palmitate-induced inflammatory responses in endothelial cells. PLoS ONE 2015, 10, e0125111. [Google Scholar]
- Stachon, P.; Geis, S.; Peikert, A.; Heidenreich, A.; Michel, N.A.; Unal, F.; Hoppe, N.; Dufner, B.; Schulte, L.; Marchini, T.; et al. Extracellular ATP induces vascular inflammation and atherosclerosis via purinergic receptor Y2 in mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1577–1586. [Google Scholar] [CrossRef] [Green Version]
- Molica, F.; Figueroa, X.F.; Kwak, B.R.; Isakson, B.E.; Gibbins, J.M. Connexins and pannexins in vascular function and disease. Int. J. Mol. Sci. 2018, 19, 1663. [Google Scholar] [CrossRef] [Green Version]
- Penuela, S.; Harland, L.; Simek, J.; Laird, D.W. Pannexin channels and their links to human disease. Biochem. J. 2014, 461, 371–381. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Schappe, M.S.; Desai, B.N.; Bayliss, D.A. Revisiting multimodal activation and channel properties of pannexin 1. J. Gen. Physiol. 2018, 150, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Gaynullina, D.; Shestopalov, V.I.; Panchin, Y.; Tarasova, O.S. Pannexin 1 facilitates arterial relaxation via an endothelium-derived hyperpolarization mechanism. FEBS Lett. 2015, 589, 1164–1170. [Google Scholar] [CrossRef] [Green Version]
- Lohman, A.W.; Leskov, I.L.; Butcher, J.T.; Johnstone, S.R.; Stokes, T.A.; Begandt, D.; DeLalio, L.J.; Best, A.K.; Penuela, S.; Leitinger, N.; et al. Pannexin 1 channels regulate leukocyte emigration through the venous endothelium during acute inflammation. Nat. Commun. 2015, 6, 7965. [Google Scholar] [CrossRef]
- Sharma, A.K.; Charles, E.J.; Zhao, Y.; Narahari, A.K.; Baderdinni, P.K.; Good, M.E.; Lorenz, U.M.; Kron, I.L.; Bayliss, D.A.; Ravichandran, K.S.; et al. Pannexin-1 channels on endothelial cells mediate vascular inflammation during lung ischemia-reperfusion injury. Am. J. Physiol. Lung. Cell Mol Physiol. 2018, 315, L301–L312. [Google Scholar] [CrossRef] [PubMed]
- Molica, F.; Meens, M.J.; Dubrot, J.; Ehrlich, A.; Roth, C.L.; Morel, S.; Pelli, G.; Vinet, L.; Braunersreuther, V.; Ratib, O.; et al. Pannexin1 links lymphatic function to lipid metabolism and atherosclerosis. Sci. Rep. 2017, 7, 13706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, V.R.; Barr, K.J.; Kelly, J.J.; Johnston, D.; Brown, C.F.C.; Robb, K.P.; Sayedyahossein, S.; Huang, K.; Gros, R.; Flynn, L.E.; et al. Pannexin 1 regulates adipose stromal cell differentiation and fat accumulation. Sci. Rep. 2018, 8, 16166. [Google Scholar] [CrossRef] [PubMed]
- Molica, F.; Morel, S.; Meens, M.J.; Denis, J.F.; Bradfield, P.F.; Penuela, S.; Zufferey, A.; Monyer, H.; Imhof, B.A.; Chanson, M.; et al. Functional role of a polymorphism in the pannexin1 gene in collagen-induced platelet aggregation. Thromb. Haemost. 2015, 114, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quercioli, A.; Pataky, Z.; Vincenti, G.; Makoundou, V.; Di Marzo, V.; Montecucco, F.; Carballo, S.; Thomas, A.; Staub, C.; Steffens, S.; et al. Elevated endocannabinoid plasma levels are associated with coronary circulatory dysfunction in obesity. Eur. Heart J. 2011, 32, 1369–1378. [Google Scholar] [CrossRef] [Green Version]
- Quercioli, A.; Pataky, Z.; Montecucco, F.; Carballo, S.; Thomas, A.; Staub, C.; Di Marzo, V.; Vincenti, G.; Ambrosio, G.; Ratib, O.; et al. Coronary vasomotor control in obesity and morbid obesity: Contrasting flow responses with endocannabinoids, leptin, and inflammation. JACC Cardiovasc. Imaging 2012, 5, 805–815. [Google Scholar] [CrossRef]
- Schindler, T.H.; Cardenas, J.; Prior, J.O.; Facta, A.D.; Kreissl, M.C.; Zhang, X.L.; Sayre, J.; Dahlbom, M.; Licinio, J.; Schelbert, H.R. Relationship between increasing body weight, insulin resistance, inflammation, adipocytokine leptin, and coronary circulatory function. J. Am. Coll. Cardiol. 2006, 47, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Stierlin, F.B.; Molica, F.; Reny, J.L.; Kwak, B.R.; Fontana, P. Pannexin1 single nucleotide polymorphism and platelet reactivity in a cohort of cardiovascular patients. Cell Commun. Adhes. 2017, 23, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic syndrome: A comprehensive perspective based on interactions between obesity, diabetes, and inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Shamai, L.; Lurix, E.; Shen, M.; Novaro, G.M.; Szomstein, S.; Rosenthal, R.; Hernandez, A.V.; Asher, C.R. Association of body mass index and lipid profiles: Evaluation of a broad spectrum of body mass index patients including the morbidly obese. Obes. Surg. 2011, 21, 42–47. [Google Scholar] [CrossRef]
- Sinaiko, A.R.; Steinberger, J.; Moran, A.; Prineas, R.J.; Vessby, B.; Basu, S.; Tracy, R.; Jacobs, D.R., Jr. Relation of body mass index and insulin resistance to cardiovascular risk factors, inflammatory factors, and oxidative stress during adolescence. Circulation 2005, 111, 1985–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargiotas, P.; Krenz, A.; Hormuzdi, S.G.; Ridder, D.A.; Herb, A.; Barakat, W.; Penuela, S.; von Engelhardt, J.; Monyer, H.; Schwaninger, M. Pannexins in ischemia-induced neurodegeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 20772–20777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamson, S.E.; Meher, A.K.; Chiu, Y.H.; Sandilos, J.K.; Oberholtzer, N.P.; Walker, N.N.; Hargett, S.R.; Seaman, S.A.; Peirce-Cottler, S.M.; Isakson, B.E.; et al. Pannexin 1 is required for full activation of insulin-stimulated glucose uptake in adipocytes. Mol. Metab. 2015, 4, 610–618. [Google Scholar] [CrossRef]
- Billaud, M.; Chiu, Y.H.; Lohman, A.W.; Parpaite, T.; Butcher, J.T.; Mutchler, S.M.; DeLalio, L.J.; Artamonov, M.V.; Sandilos, J.K.; Best, A.K.; et al. A molecular signature in the pannexin1 intracellular loop confers channel activation by the alpha1 adrenoreceptor in smooth muscle cells. Sci. Signal 2015, 8, ra17. [Google Scholar] [CrossRef] [Green Version]
- Weilinger, N.L.; Lohman, A.W.; Rakai, B.D.; Ma, E.M.; Bialecki, J.; Maslieieva, V.; Rilea, T.; Bandet, M.V.; Ikuta, N.T.; Scott, L.; et al. Metabotropic nmda receptor signaling couples src family kinases to pannexin-1 during excitotoxicity. Nat. Neurosci. 2016, 19, 432–442. [Google Scholar] [CrossRef]
- Burma, N.E.; Bonin, R.P.; Leduc-Pessah, H.; Baimel, C.; Cairncross, Z.F.; Mousseau, M.; Shankara, J.V.; Stemkowski, P.L.; Baimoukhametova, D.; Bains, J.S.; et al. Blocking microglial pannexin-1 channels alleviates morphine withdrawal in rodents. Nat. Med. 2017, 23, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.L.; Arandjelovic, S.; Brown, G.S.K.M.M.S.S.; Buckley, M.W.; Chiu, Y.H.; Shu, S.; Kim, J.K.; Chung, J.; et al. Hematopoietic pannexin 1 function is critical for neuropathic pain. Sci. Rep. 2017, 7, 42550. [Google Scholar] [CrossRef] [Green Version]
- Bartley, C.; Brun, T.; Oberhauser, L.; Grimaldi, M.; Molica, F.; Kwak, B.R.; Bosco, D.; Chanson, M.; Maechler, P. Chronic fructose renders pancreatic beta-cells hyper-responsive to glucose-stimulated insulin secretion through extracellular atp signaling. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E25–E41. [Google Scholar] [CrossRef] [Green Version]
- Michalski, K.; Henze, E.; Nguyen, P.; Lynch, P.; Kawate, T. The weak voltage dependence of pannexin 1 channels can be tuned by n-terminal modifications. J. Gen. Physiol. 2018, 150, 1758–1768. [Google Scholar] [CrossRef] [Green Version]
- Prieto, D.; Contreras, C.; Sanchez, A. Endothelial dysfunction, obesity and insulin resistance. Curr. Vasc. Pharmacol. 2014, 12, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, M.; Higashi, Y. Triglycerides and endothelial function: Molecular biology to clinical perspective. Curr. Opin. Lipidol. 2019, 30, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lin, Y.; Wang, S.; Zhang, L.; Guo, L. Glp-1 inhibits high-glucose-induced oxidative injury of vascular endothelial cells. Sci. Rep. 2017, 7, 8008. [Google Scholar] [CrossRef] [PubMed]
- Symons, J.D. Opportunity “nox”: A novel approach to preventing endothelial dysfunction in the context of insulin resistance. Diabetes 2013, 62, 1818–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vita, J.A.; Treasure, C.B.; Nabel, E.G.; McLenachan, J.M.; Fish, R.D.; Yeung, A.C.; Vekshtein, V.I.; Selwyn, A.P.; Ganz, P. Coronary vasomotor response to acetylcholine relates to risk factors for coronary artery disease. Circulation 1990, 81, 491–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celermajer, D.S.; Sorensen, K.E.; Bull, C.; Robinson, J.; Deanfield, J.E. Endothelium-dependent dilation in the systemic arteries of asymptomatic subjects relates to coronary risk factors and their interaction. J. Am. Coll. Cardiol. 1994, 24, 1468–1474. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Cohort (n = 72) | Panx1-400C Cohort (n = 40) | Panx1-400A Cohort (n = 32) | p-Value | |
|---|---|---|---|---|
| Age, years, mean ± SEM | 43 ± 1 | 45 ± 2 | 42 ± 2 | 0.63 |
| Female/male, n (%) | 19 (26)/53 (74) | 10 (25)/30 (75) | 9 (28)/23 (72) | 0.89 |
| Weight (kg), mean ± SEM | 90 ± 3 | 88 ± 3 | 93 ± 5 | 0.58 |
| Height (cm), mean ± SEM | 173 ± 1 | 173 ± 1 | 173 ± 2 | 0.98 |
| BMI (kg/m2), mean ± SEM | 30 ± 1 | 45 ± 2 | 31 ± 1 | 0.58 |
| Heart rate at rest (b.p.m.),median (IQR) | 63 (56–70) | 63 (55–70) | 64 (58–70) | 0.97 |
| SBP at rest (b.p.m.),median (IQR) | 119 (110–129) | 118 (106–126) | 120 (112–132) | 0.28 |
| DBP at rest (b.p.m.),median (IQR) | 74 (68–80) | 73 (67–80) | 76 (68–83) | 0.75 |
| Full Cohort (n = 72) | Panx1-400C Cohort (n = 40) | Panx1-400A Cohort (n = 32) | p-Value | |
|---|---|---|---|---|
| MBF at rest (mL/g/min), median (IQR) | 0.72 (0.62–0.78) | 0.72 (0.63–0.76) | 0.72 (0.62–0.79) | 0.99 |
| MBF during CPT (mL/g/min), median (IQR) | 0.85 (0.76–1.04) | 0.90 (0.75–1.04) | 0.84 (0.76–1.05) | 0.99 |
| MBF during hyperemia (mL/g/min), median (IQR) | 2.11 (1.79–2.42) | 2.15 (1.84–2.44) | 2.08 (1.71–2.32) | 0.59 |
| MFR, median (IQR) | 3.03 (2.41–3.43) | 3.28 (2.52–3.50) | 2.76 (2.38–3.31) | 0.40 |
| ΔMBF to CPT from rest (mL/g/min), mean ± SEM | 0.186 ± 0.020 | 0.179 ± 0.026 | 0.194 ± 0.030 | 0.93 |
| Full Cohort Non-Obese (n = 43) | Full Cohort Obese (n = 29) | p-Value | Panx1-400C Non-Obese (n = 25) | Panx1-400C Obese (n = 15) | p-Value | Panx1-400A Non-Obese (n = 18) | Panx1-400A Obese (n = 14) | p-Value | |
|---|---|---|---|---|---|---|---|---|---|
| MBF at rest (mL/g/min), median (IQR) | 0.72 (0.63–0.76) | 0.74 (0.62–0.84) | 0.35 | 0.72 (0.65–0.75) | 0.69 (0.57–0.84) | 0.74 | 0.70 (0.60–0.79) | 0.75 (0.63–0.80) | 0.42 |
| MBF during CPT (mL/g/min), median (IQR) | 0.93 (0.81–1.05) | 0.78 (0.71–0.92) | <0.01 | 0.93 (0.83–1.04) | 0.76 (0.70–0.93) | 0.06 | 0.92 (0.78–1.10) | 0.80 (0.75–0.87) | 0.12 |
| MBF during hyperemia (mL/g/min), median (IQR) | 2.18 (1.71–2.44) | 2.08 (1.84–2.31) | 0.43 | 2.24 (1.98–2.47) | 1.95 (1.44–2.41) | 0.14 | 1.94 (1.59–2.40) | 2.10 (1.95–2.23) | 0.51 |
| MFR, median (IQR) | 3.22 (2.6–3.5) | 2.69 (2.34–3.42) | 0.16 | 3.3 (2.78–3.58) | 2.64 (2.22–3.49) | 0.20 | 2.92 (2.36–3.38) | 2.69 (2.36–3.30) | 0.70 |
| ΔMBF to CPT from rest (mL/g/min), mean ± SEM | 0.246 ± 0.025 | 0.095 ± 0.023 | <0.001 | 0.238 ± 0.032 | 0.081 ± 0.033 | <0.01 | 0.259 ± 0.042 | 0.111 ± 0.035 | <0.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molica, F.; Quercioli, A.; Montecucco, F.; Schindler, T.H.; Kwak, B.R.; Morel, S. A Genetic Polymorphism in the Pannexin1 Gene Predisposes for The Development of Endothelial Dysfunction with Increasing BMI. Biomolecules 2020, 10, 208. https://doi.org/10.3390/biom10020208
Molica F, Quercioli A, Montecucco F, Schindler TH, Kwak BR, Morel S. A Genetic Polymorphism in the Pannexin1 Gene Predisposes for The Development of Endothelial Dysfunction with Increasing BMI. Biomolecules. 2020; 10(2):208. https://doi.org/10.3390/biom10020208
Chicago/Turabian StyleMolica, Filippo, Alessandra Quercioli, Fabrizio Montecucco, Thomas H. Schindler, Brenda R. Kwak, and Sandrine Morel. 2020. "A Genetic Polymorphism in the Pannexin1 Gene Predisposes for The Development of Endothelial Dysfunction with Increasing BMI" Biomolecules 10, no. 2: 208. https://doi.org/10.3390/biom10020208
APA StyleMolica, F., Quercioli, A., Montecucco, F., Schindler, T. H., Kwak, B. R., & Morel, S. (2020). A Genetic Polymorphism in the Pannexin1 Gene Predisposes for The Development of Endothelial Dysfunction with Increasing BMI. Biomolecules, 10(2), 208. https://doi.org/10.3390/biom10020208