β-Mannanase Production Using Coffee Industry Waste for Application in Soluble Coffee Processing

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate
2.2. Microorganisms
2.3. Solid-State Fermentation in Flasks
2.4. Solid-State Fermentation in a Column-Type Bioreactor
2.5. Effect of Temperature and pH on β-Mannanase Activity
2.6. Enzymatic Hydrolysis of Coffee Residue
2.7. Analytical Methods
3. Results and Discussion
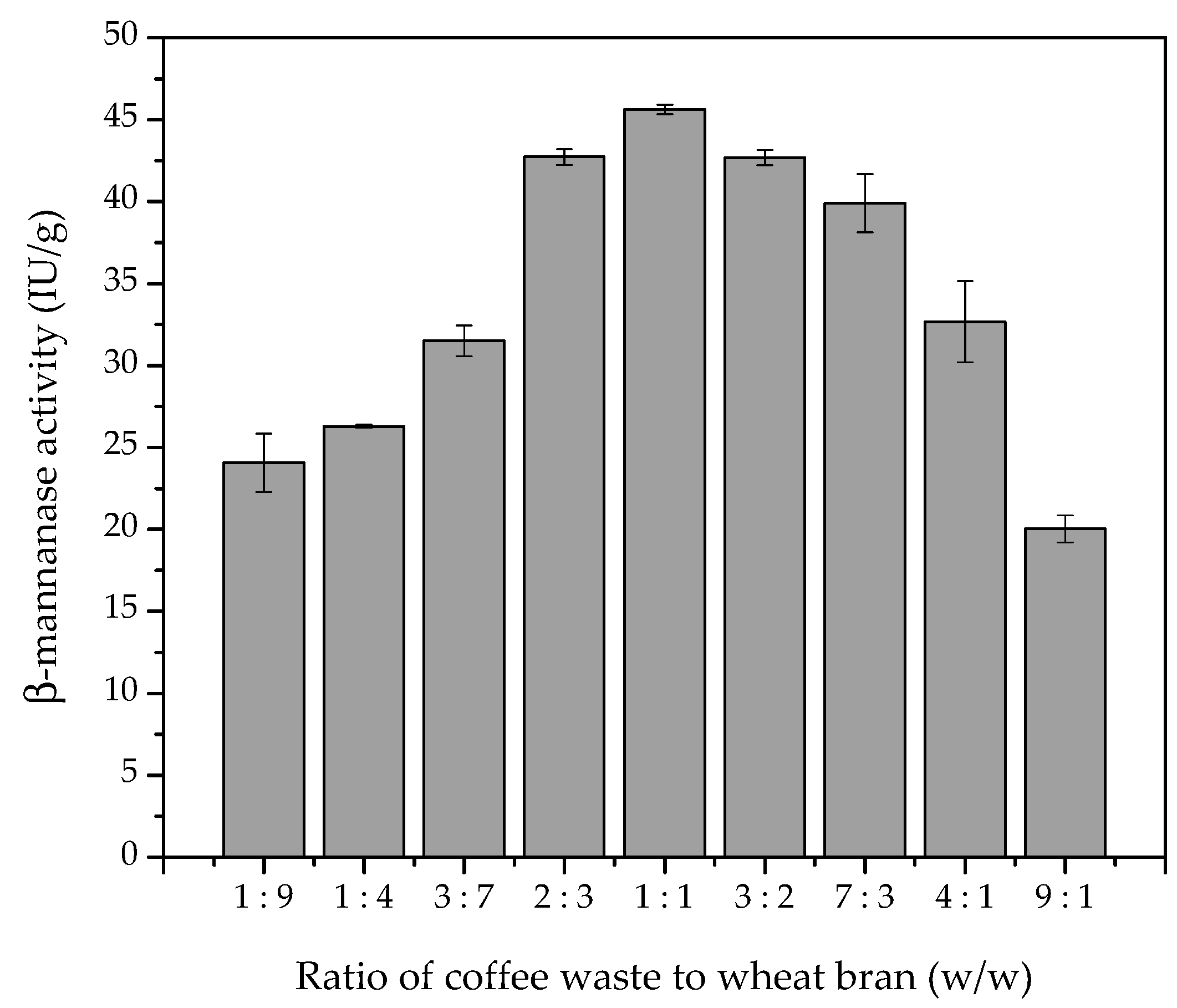
3.1. Effect of Substrate on Production of β-Mannanase
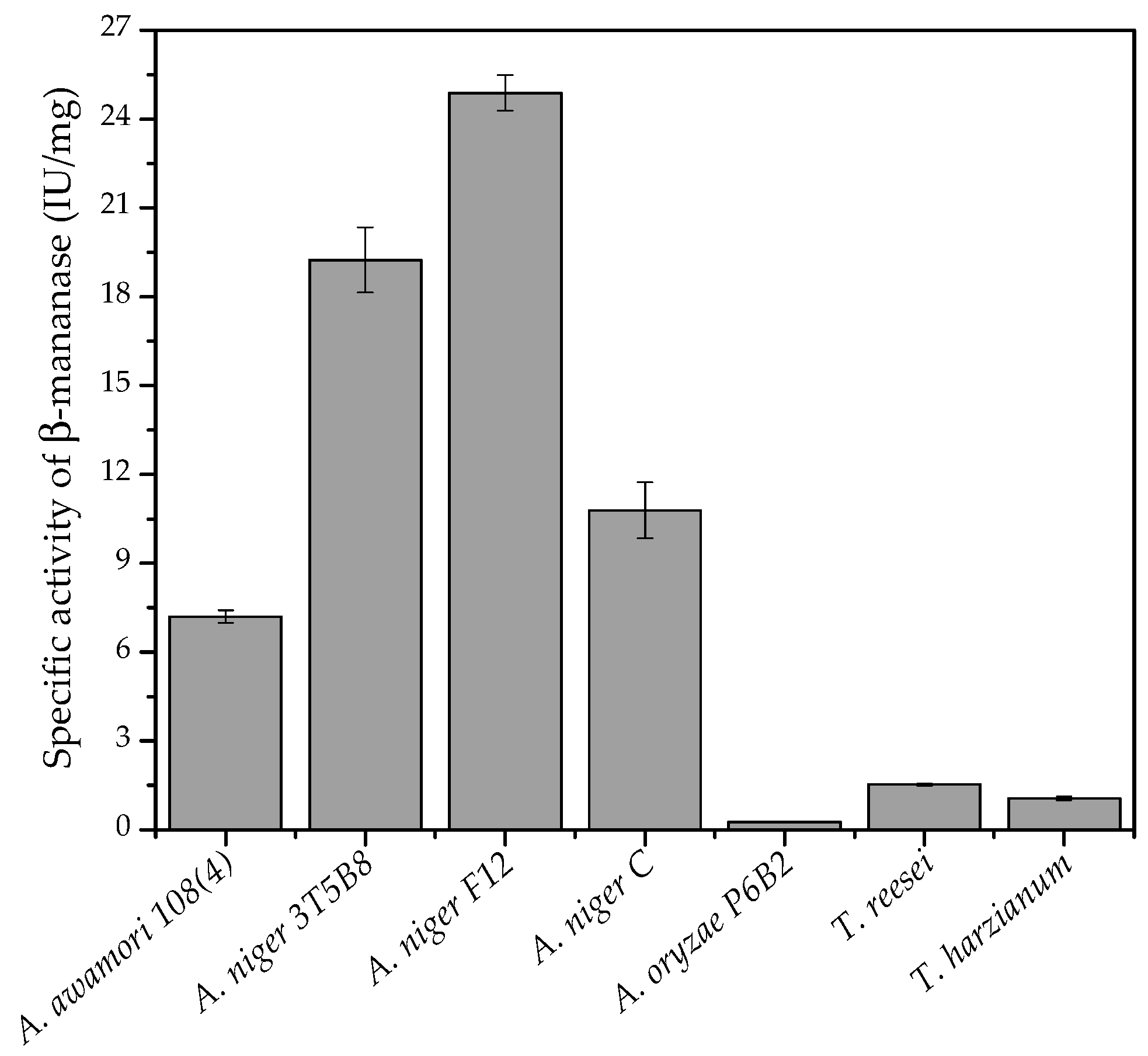
3.2. Effect of Different Fungi on Enzyme Production
3.3. Comparison of the Types of Cultivation Under SSF
3.4. Optimization of β-Mannanase and β-Glucosidase Production in Bioreactor Fermentation
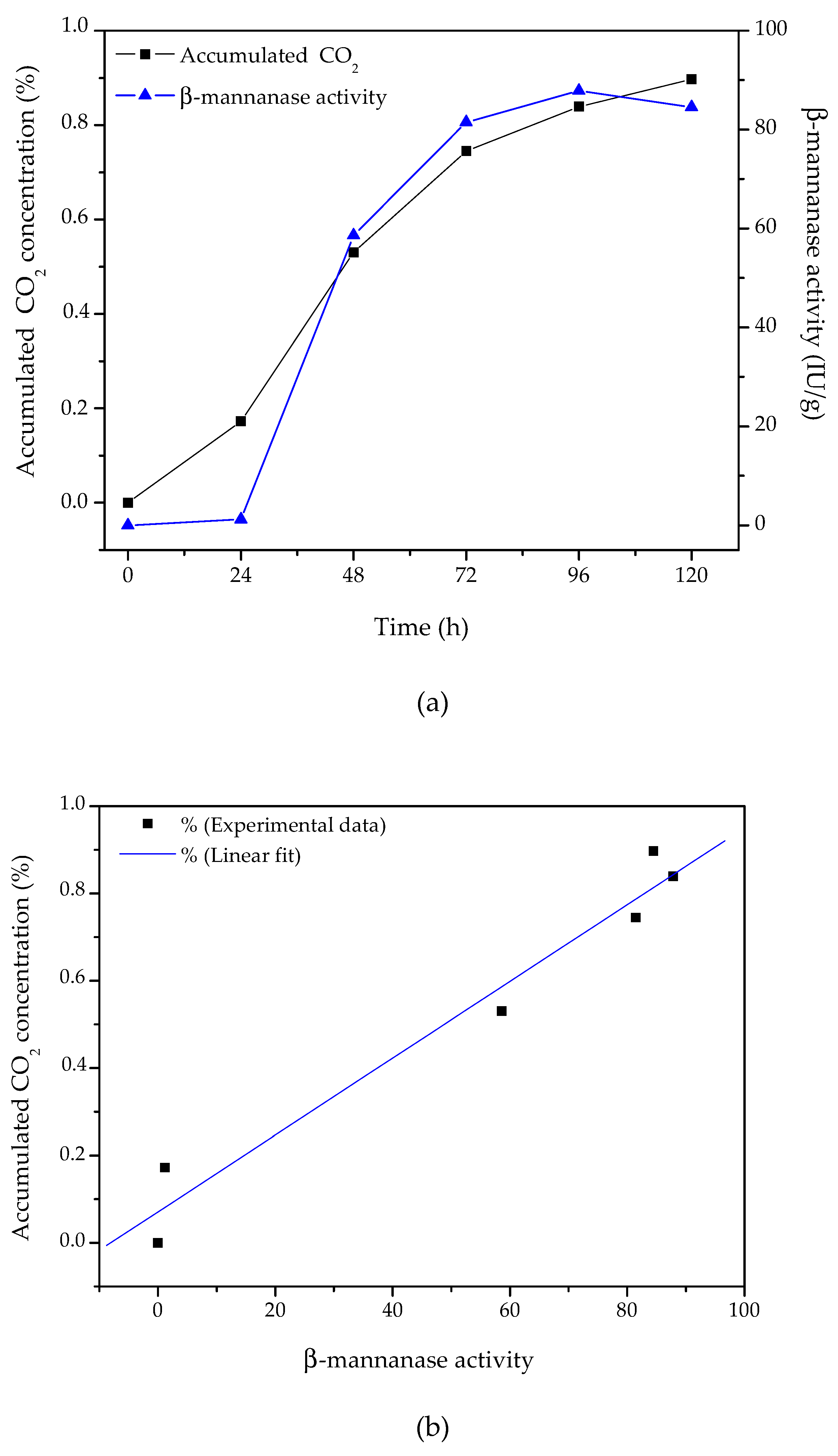
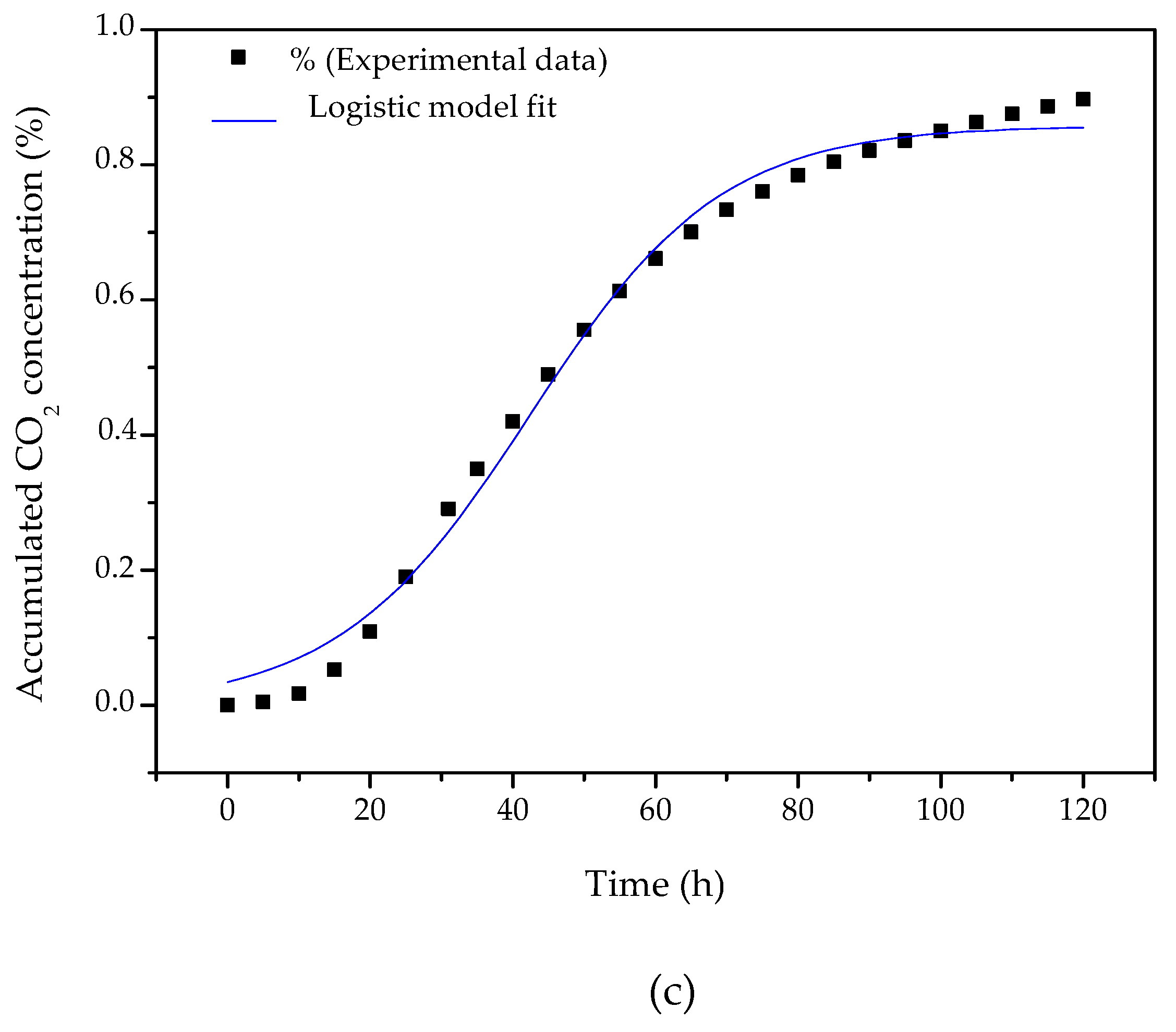
3.5. Time Profile of β-Mannanase Production
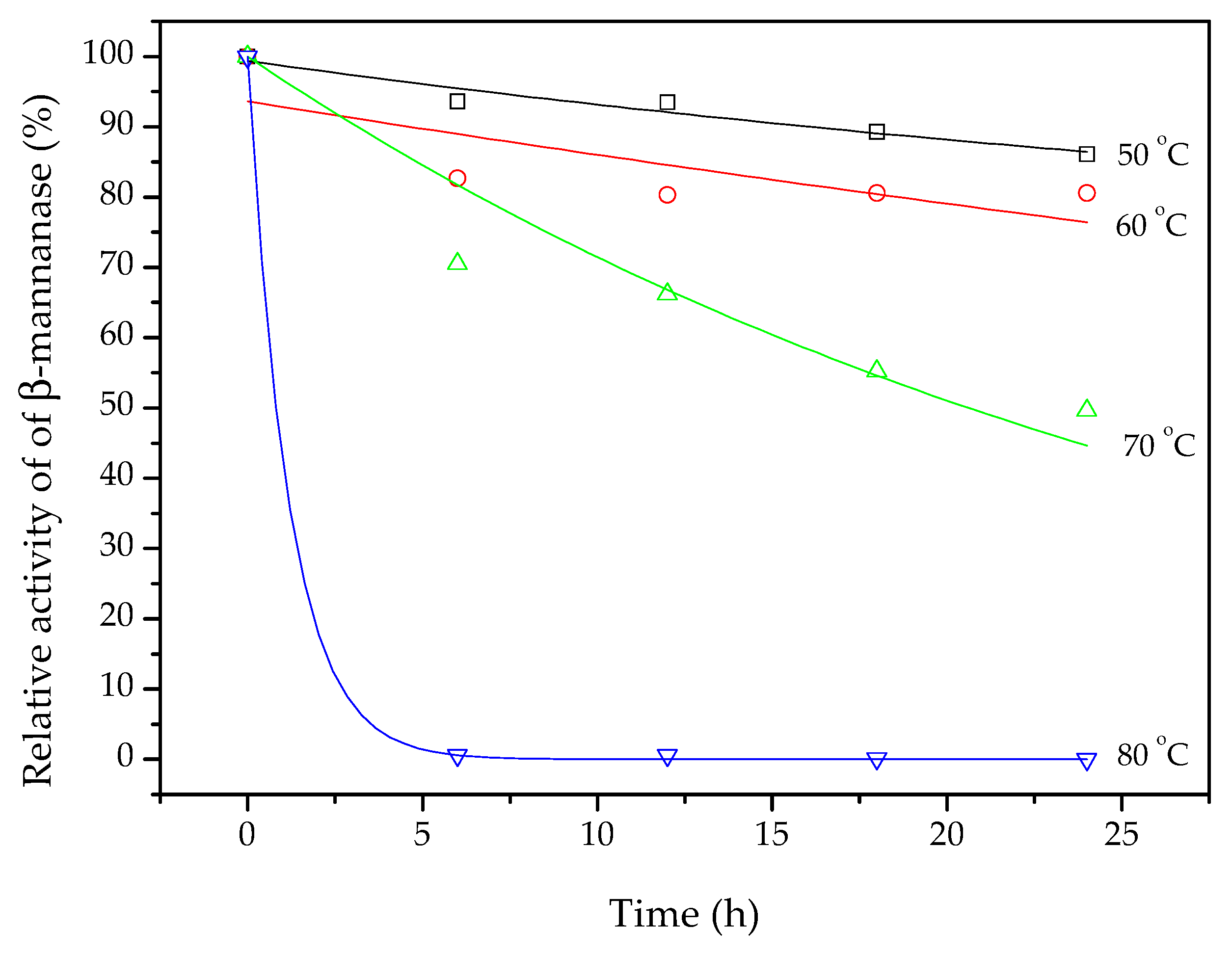
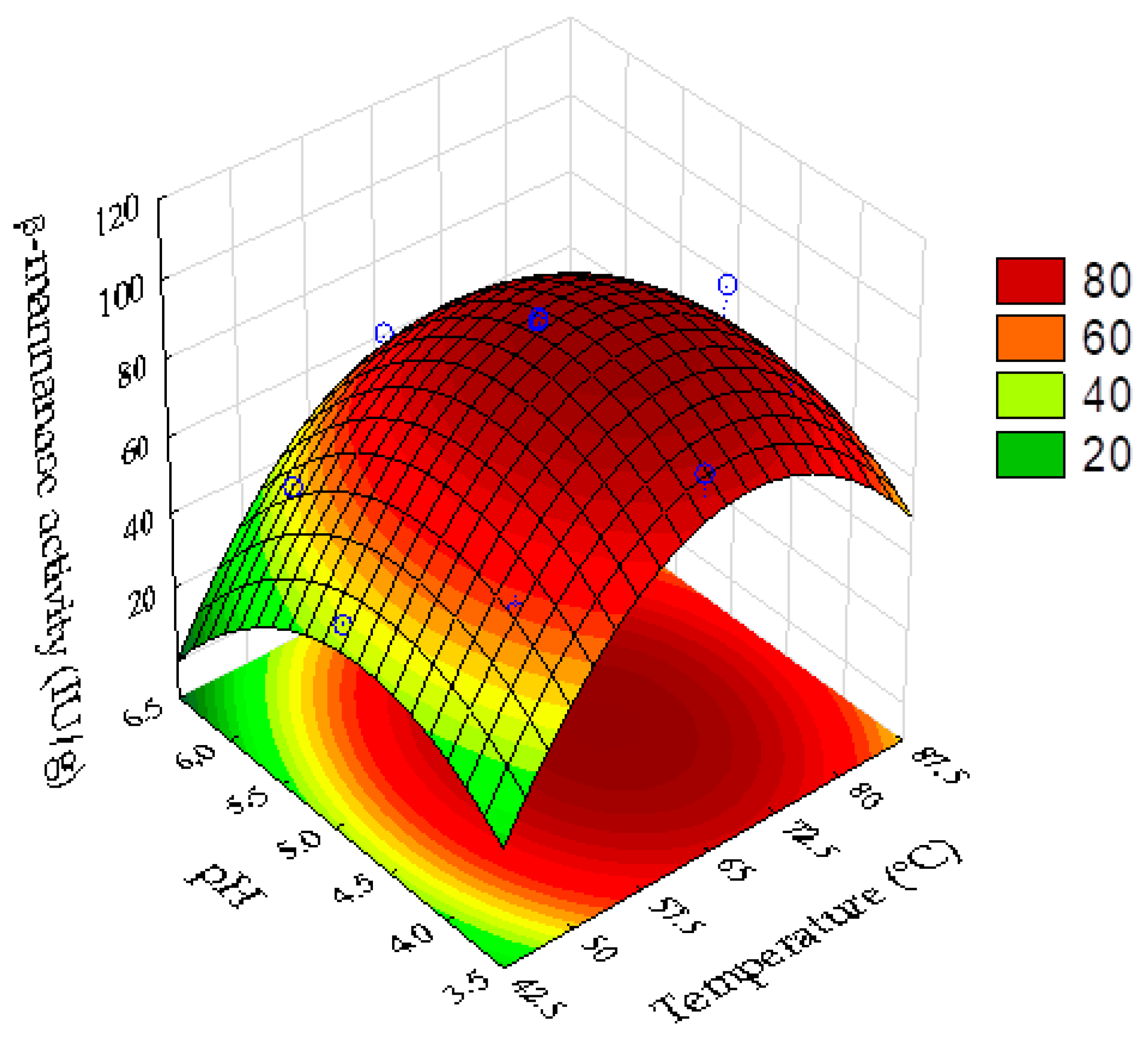
3.6. Thermostability and Optimum Temperature and pH of β-Mannanase
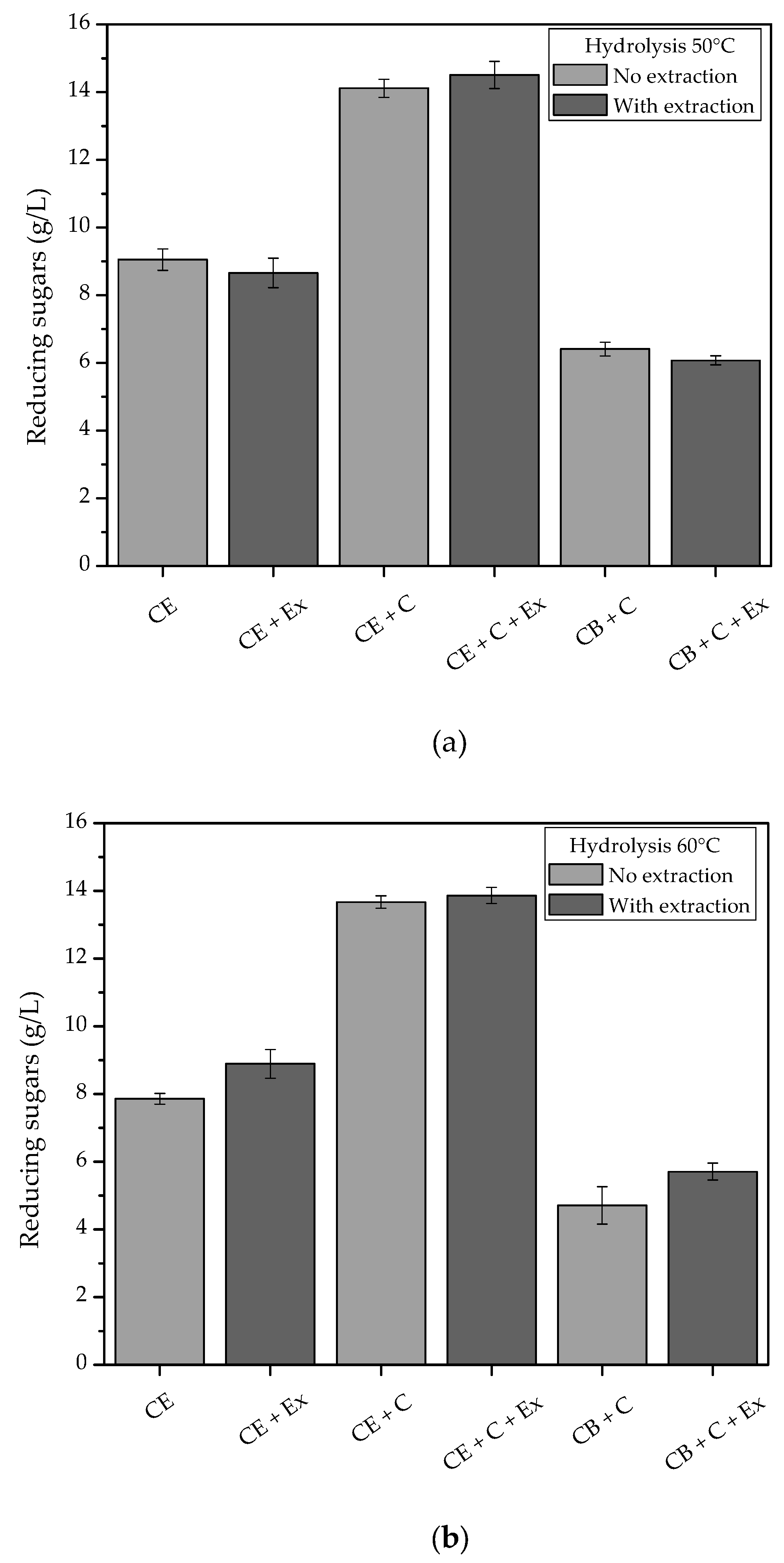
3.7. Enzymatic Hydrolysis of Coffee Residue
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Coffee Organization. Available online: www.ico.org (accessed on 22 November 2019).
- Diaz-Rubio, M.E.; Saura-Calixto, F. Dietary fiber in brewed coffee. J. Agric. Food Chem. 2007, 55, 1999–2003. [Google Scholar] [CrossRef] [Green Version]
- Nunes, F.M.; Coimbra, M.A. Chemical characterization of the high molecular weight material extracted with hot water from green and roasted robusta coffee as affected by the degree of roast. J. Agric. Food Chem. 2002, 50, 7046–7052. [Google Scholar] [CrossRef] [PubMed]
- Redgwell, R.J.; Trovato, V.; Curti, D.; Fischer, M. Effect of roasting on degradation and structural features of polysaccharides in Arabica coffee beans. Carbohydr. Res. 2002, 337, 421–431. [Google Scholar] [CrossRef]
- Baraldi, I.J.; Giordano, R.L.C.; Zangirolami, T.C. Enzymatic hydrolysis as an environmentally friendly process compared to thermal hydrolysis for instant coffee production. Braz. J. Chem. Eng. 2016, 33, 763–771. [Google Scholar] [CrossRef] [Green Version]
- De Maria, C.A.B.; Trugo, L.C.; Neto, F.R.A.; Moreira, R.F.A. Arabinogalactan as a potential furfural precursor in roasted coffee. Int. J. Food Sci. Technol. 1994, 29, 559–562. [Google Scholar] [CrossRef]
- Soni, H.; Kango, N. Microbial Mannanases: Properties and Applications. In Advances in Enzyme Biotechnology; Shukla, P., Pletschke, B., Eds.; Springer: New Delhi, India, 2013; pp. 41–56. [Google Scholar]
- Ozturk, B.; Cekmecelioglu, D.; Ogel, Z.B. Optimal conditions for enhanced beta-mannanase production by recombinant Aspergillus sojae. J. Mol. Catal. B Enzym. 2010, 64, 135–139. [Google Scholar] [CrossRef]
- Abdeshahian, P.; Samat, N.; Hamid, A.A.; Yusoff, W.M.W. Utilization of palm kernel cake for production of beta-mannanase by Aspergillus niger FTCC 5003 in solid substrate fermentation using an aerated column bioreactor. J. Ind. Microbiol. Biotechnol. 2010, 37, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Maijala, P.; Kango, N.; Szijarto, N.; Viikari, L. Characterization of hemicellulases from thermophilic fungi. Antonie Van Leeuwenhoek. 2012, 101, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.Y.; Pradeep, G.C.; Lee, S.K.; Park, D.H.; Cho, S.S.; Choi, Y.H.; Yoo, J.C.; Kim, S.W. Understanding beta-mannanase from Streptomyces sp CS147 and its potential application in lignocellulose based biorefining. Biotechnol. J. 2015, 10, 1894–1902. [Google Scholar] [CrossRef]
- Meenakshi, M.; Singh, G.; Bhalla, A.; Hoondal, G.S. Solid-state fermentation and characterization of partially purified thermostable mannanase from Bacillus sp. MG-33. Bioresources 2010, 5, 1689–1701. [Google Scholar]
- Mou, H.; Zhou, F.; Jiang, X.; Liu, Z. Production, purification and properties of beta-mannanase from soil bacterium Bacillus circulans M-21. J. Food Biochem. 2011, 35, 1451–1460. [Google Scholar] [CrossRef]
- Ge, J.P.; Du, R.P.; Zhao, D.; Song, G.; Jim, M.; Ping, W.X. Bio-chemical characterization of a β-mannanase from Bacillus licheniformis HDYM-04 isolated from flax water-retting liquid and its decolorization ability of dyes. RSC Adv. 2016, 6, 23612–23621. [Google Scholar] [CrossRef]
- Holker, U.; Hofer, M.; Lenz, J. Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Appl. Microbiol. Biotechnol. 2004, 64, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Sukumaran, R.K.; Patel, A.K.; Larroche, C.; Pandey, A. Advancement and comparative profiles in the production technologies using solid-state and submerged fermentation for microbial cellulases. Enzyme Microb. Technol. 2010, 46, 541–549. [Google Scholar] [CrossRef]
- Farinas, C.S. Developments in solid-state fermentation for the production of biomass-degrading enzymes for the bioenergy sector. Renew. Sustain. Energy Rev. 2015, 52, 179–188. [Google Scholar] [CrossRef]
- Casciatori, F.P.; Casciatori-Frassatto, P.A. Enzymes for the Biochemical Route of Second-Generation Ethanol: Production by Solid-State Cultivation as a Feasible and Sustainable Alternative. In Bioethanol and Beyond: Advances in Production Process and Future Directions; Brienzo, M., Ed.; Nova Science Publishers: New York, NY, USA, 2018; pp. 285–313. [Google Scholar]
- Murthy, P.S.; Naidu, M.M. Sustainable management of coffee industry by-products and value addition—A review. Resour. Conserv. Recycl. 2012, 66, 45–58. [Google Scholar] [CrossRef]
- Cerda, A.; Mejías, L.; Gea, T.; Sánchez, A. Cellulase and xylanase production at pilot scale by solid-state fermentation from coffee husk using specialized consortia: The consistency of the process and the microbial communities involved. Bioresour. Technol. 2017, 243, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Kourmentza, C.; Economou, C.N.; Tsafrakidou, R.; Kornaros, M. Spent coffee grounds make much more than waste: Exploring recent advances and future exploitation strategies for the valorization of an emerging food waste stream. J. Clean. Prod. 2018, 172, 980–992. [Google Scholar] [CrossRef]
- Kovalcik, A.; Obruca, S.; Marova, I. Valorization of spent coffee grounds: A review. Food Bioprod. Process. 2018, 110, 104–119. [Google Scholar] [CrossRef]
- McNutt, J.; He, Q. Spent coffee grounds: A review on current utilization. J. Ind. Eng. Chem. 2019, 71, 78–88. [Google Scholar] [CrossRef]
- Mandels, M.; Stenberg, D. Recent advances in cellulase technology. J. Ferment. Technol. 1976, 54, 267–286. [Google Scholar]
- Durand, A. Bioreactor designs for solid state fermentation. Biochem. Eng. J. 2003, 13, 113–125. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Stalbrand, H.; Siika-Aho, M.; Tenkanen, M.; Viikari, L. Purification and characterization of two β-mannanases from Trichoderma reesei. J. Biotechnol. 1993, 29, 229–242. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Javed, M.M.; Zahoor, S.; Shafaat, S.; Mehmooda, I.; Gul, A.; Rasheed, H.; Bukhari, S.A.I.; Aftab, M.N.; Ikram-ul-Haq. Wheat bran as a brown gold: Nutritious value and its biotechnological applications. Afr. J. Microbiol. Res. 2012, 6, 724–733. [Google Scholar]
- Cavalcante, R.S.; Lima, H.L.S.; Pinto, G.A.S.; Gava, C.A.T.; Rodrigues, S. Effect of moisture on Trichoderma conidia production on corn and wheat bran by solid state fermentation. Food Bioprocess. Technol. 2008, 1, 100–104. [Google Scholar] [CrossRef]
- Malathi, S.; Chakraborty, R. Production of alkaline protease by a new Aspergillus flavus isolate under solid-substrate fermentation conditions for use as a depilation agent. Appl. Environ. Microbiol. 1991, 57, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Ellaiah, P.; Adinarayana, K.; Bhavani, Y.; Padmaja, P.; Srinivasulu, B. Optimization of process parameters for glucoamylase production under solid state fermentation by a newly isolated Aspergillus species. Process. Biochem. 2002, 38, 615–620. [Google Scholar] [CrossRef]
- De Vries, R.P.; Visser, J. Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol. Mol. Biol. Rev. 2001, 65, 497. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.M.; Filho, E.X.F. Purification and characterization of a β-mannanase from Trichoderma harzianum strain T4. Carbohydr. Polym. 2004, 57, 23–29. [Google Scholar] [CrossRef]
- Arora, S.; Rani, R.; Ghosh, S. Bioreactors in solid state fermentation technology: Design, applications and engineering aspects. J. Biotechnol. 2018, 269, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Palomino, L.R.; Biasetto, C.R.; Araujo, A.R.; Bianchi, V.L.D. Enhanced extraction of phenolic compounds from coffee industry’s residues through solid state fermentation by Penicillium purpurogenum. Food Sci. Technol. 2015, 35, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Farinas, C.S.; Vitcosque, G.L.; Fonseca, R.F.; Bertucci Neto, V.; Couri, S. Modeling the effects of solid-state fermentation operating conditions on endoglucanase production using an instrumented bioreactor. Ind. Crop. Prod. 2011, 34, 1186–1192. [Google Scholar] [CrossRef]
- Viccini, G.; Mitchell, D.A.; Boit, S.D.; Gern, J.C.; da Rosa, A.S.; Costa, R.M.; Dalsenter, F.D.H.; von Meien, O.F.; Krieger, N. Analysis of growth kinetic profiles in solid-state fermentation. Food Technol. Biotechnol. 2001, 39, 271–294. [Google Scholar]
- Glenn, D.R.; Rogers, P.L. A solid substrate process for an animal feed product: Studies on fungal strain improvement. Aust. J. Biotechnol. 1988, 2, 50–57. [Google Scholar]
- Saucedo-Castañeda, G.; Gutiérrez-Rojas, M.; Bacquet, G.; Raimbault, M.; Viniegra-González, G. Heat transfer simulation in solid substrate fermentation. Biotechnol. Bioeng. 1990, 35, 802–808. [Google Scholar] [CrossRef]
- Sangsurasak, P.; Mitchell, D.A. The investigation of transient multidimensional heat transfer in solid state fermentation. Chem. Eng. J. 1995, 60, 199–204. [Google Scholar] [CrossRef]
- von Meien, O.F.; Mitchell, D.A. A two-phase model for water and heat transfer within an intermittently-mixed solid-state fermentation bioreactor with forced aeration. Biotechnol. Bioeng. 2002, 79, 416–428. [Google Scholar] [CrossRef]
- Casciatori, F.P.; Bück, A.; Thoméo, J.C.; Tsotsas, E. Two-phase and two-dimensional model describing heat and water transfer during solid-state fermentation within a packed-bed bioreactor. Chem. Eng. J. 2016, 287, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Farinas, C.S.; Loyo, M.M.; Baraldo Junior, A.; Tardioli, W.; Bertucci Neto, V.; Couri, S. Finding stable cellulase and xylanase: Evaluation of the synergistic effect of pH and temperature. New Biotechnol. 2010, 27, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Soni, H.; Rawat, H.K.; Pletschke, B.I.; Kango, N. Purification and characterization of beta-mannanase from Aspergillus terreus and its applicability in depolymerization of mannans and saccharification of lignocellulosic biomass. 3 Biotech 2016, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doran, P.M. Bioprocess Engineering Principles, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Sizer, I.W. Effects of temperature on enzyme kinetics. Adv. Enzym. 1943, 3, 35–62. [Google Scholar]
- Catalán, E.; Komilis, D.; Sánchez, A. Environmental impact of cellulase production from coffee husks by solid-state fermentation: A life-cycle assessment. J. Clean. Prod. 2019, 233, 954–962. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Time (h) | Enzymatic Activity (IU/g) * | ||
|---|---|---|---|---|
| SB + CW | CW | WB + CW | ||
| β-mannanase | 72 | 7.33 ± 0.44 c | 10.16 ± 0.65 b | 34.91 ± 0.75 a |
| 96 | 6.75 ± 0.84 c | 15.42 ± 0.32 b | 44.05 ± 0.85 a | |
| 120 | 5.61 ± 0.59 c | 15.94 ± 0.68 b | 51.97 ± 0.67 a | |
| β-glucosidase | 72 | 45.36 ± 1.15 b | 2.12 ± 0.08 c | 67.54 ± 2.29 a |
| 96 | 51.80 ± 0.07 b | 2.66 ± 0.26 c | 69.85 ± 2.40 a | |
| 120 | 47.93 ± 0.42 b | 3.03 ± 0.05 c | 72.54 ± 2.70 a | |
| Time (h) | Productivity (IU/g.h) * | |||
|---|---|---|---|---|
| Erlenmeyer Flasks | Column Bioreactor | |||
| β-Mannanase | β-Glucosidase | β-Mannanase | β-Glucosidase | |
| 72 | 0.48 ± 0.01 | 0.94 ± 0.03 | 0.73 ± 0.02 | 1.00 ± 0.03 |
| 96 | 0.46 ± <0.01 | 0.73 ± 0.02 | 0.62 ± <0.01 | 0.82 ± 0.04 |
| 120 | 0.43 ± <0.01 | 0.60 ± 0.02 | 0.49 ± 0.05 | 0.66 ± 0.03 |
| Run | X1 a | X2 b | X3 c | β-Mannanase (IU/g) * | β-Glucosidase (IU/g) * |
|---|---|---|---|---|---|
| 1 | −1 (29) | −1 (5) | −1 (5.6) | 37.31 ± 0.46 | 35.58 ± 2.06 |
| 2 | +1 (35) | −1 (5) | −1 (5.6) | 39.26 ± 3.71 | 32.92 ± 0.91 |
| 3 | −1 (29) | +1 (7) | −1 (5.6) | 35.96 ± 0.47 | 40.01 ± 0.61 |
| 4 | +1 (35) | +1 (7) | −1 (5.6) | 44.71 ± 2.19 | 33.65 ± 2.12 |
| 5 | −1 (29) | −1 (5) | +1 (10.4) | 35.14 ± 1.03 | 29.87 ± 2.01 |
| 6 | +1 (35) | −1 (5) | +1 (10.4) | 42.62 ± 2.03 | 34.62 ± 0.98 |
| 7 | −1 (29) | +1 (7) | +1 (10.4) | 37.91 ± 2.00 | 36.29 ± 1.72 |
| 8 | +1 (35) | +1 (7) | +1 (10.4) | 49.65 ± 0.83 | 36.09 ± 1.67 |
| 9 | 0 (32) | 0 (6) | 0 (8) | 59.03 ± 2.30 | 43.10 ± 1.60 |
| 10 | 0 (32) | 0 (6) | 0 (8) | 62.40 ±2.30 | 43.62 ± 1.39 |
| 11 | 0 (32) | 0 (6) | 0 (8) | 63.50 ± 2.97 | 41.77 ± 1.80 |
| Temperature (°C) | Exponential Decay | |
| kd (h−1) | R2 | |
| 50 | 0.006 | 0.93 |
| 60 | 0.012 | 0.39 |
| 70 | 0.033 | 0.90 |
| 80 | 0.833 | 0.99 |
| Arrhenius Parameters | ||
| Frequency factor, A (h−1) | 1.8 × 1021 | |
| Ed (kJ mol−1) | 147 | |
| R2 | 0.85 | |
| Run | X1 a | X2 b | β-Mannanase Activity (UI/g) c | β-Mannanase Predicted (UI/g) d | Relative Error (%) e |
|---|---|---|---|---|---|
| 1 | −1 (50) | −1 (4.0) | 60.55 | 61.25 | −1.15 |
| 2 | 1 (80) | −1 (4.0) | 70.97 | 80.1 | −12.87 |
| 3 | −1 (50) | 1 (6.0) | 48.49 | 47.31 | 2.44 |
| 4 | 1 (80) | 1 (6.0) | 59.12 | 66.16 | −11.9 |
| 5 | −1.41 (43.8) | 0 (5.0) | 41.99 | 43.74 | −4.17 |
| 6 | 1.41 (86.8) | 0 (5.0) | 80.38 | 70.38 | 12.44 |
| 7 | 0 (65) | −1.41 (3.59) | 85.72 | 80.37 | 6.25 |
| 8 | 0 (65) | 1.41 (6.41) | 63.25 | 60.71 | 4.01 |
| 9 | 0 (65) | 0 (5.0) | 94.92 | 94.6 | 0.33 |
| 10 | 0 (65) | 0 (5.0) | 95.29 | 94.6 | 0.72 |
| 11 | 0 (65) | 0 (5.0) | 93.51 | 94.6 | −1.17 |
| Source of Variation | Coefficient | p-Value |
|---|---|---|
| Mean | 94.6 | 0.0000 * |
| Temperature (L a) | 9.43 | 0.0155 * |
| Temperature (Q b) | −18.79 | 0.0018 * |
| pH (L) | −6.97 | 0.0446 * |
| pH (Q) | −12.1 | 0.0117 * |
| Temperature × pH | 0.05 | 0.9891 |
| R2 | 0.925 | |
| Fcalculated | 18.4 | |
| Fcalculated/Flisted | 4.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favaro, C.P.; Baraldi, I.J.; Casciatori, F.P.; Farinas, C.S. β-Mannanase Production Using Coffee Industry Waste for Application in Soluble Coffee Processing. Biomolecules 2020, 10, 227. https://doi.org/10.3390/biom10020227
Favaro CP, Baraldi IJ, Casciatori FP, Farinas CS. β-Mannanase Production Using Coffee Industry Waste for Application in Soluble Coffee Processing. Biomolecules. 2020; 10(2):227. https://doi.org/10.3390/biom10020227
Chicago/Turabian StyleFavaro, Camila P., Ilton J. Baraldi, Fernanda P. Casciatori, and Cristiane S. Farinas. 2020. "β-Mannanase Production Using Coffee Industry Waste for Application in Soluble Coffee Processing" Biomolecules 10, no. 2: 227. https://doi.org/10.3390/biom10020227
APA StyleFavaro, C. P., Baraldi, I. J., Casciatori, F. P., & Farinas, C. S. (2020). β-Mannanase Production Using Coffee Industry Waste for Application in Soluble Coffee Processing. Biomolecules, 10(2), 227. https://doi.org/10.3390/biom10020227