Danegaptide Enhances Astrocyte Gap Junctional Coupling and Reduces Ischemic Reperfusion Brain Injury in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Permanent Middle Cerebral Artery Occlusion
2.2. Quantification of Cerebral Infarction
2.3. Matrix-Assisted Laser Desorption/Ionization Imaging Mass Spectrometry (MALDI-IMS)
2.4. Astrocyte Isolation and In Vitro Hypoxia
2.5. In Vitro Scrape Loading Dye-Transfer Assay
2.6. Western Blot Analysis
2.7. Hemichannel Assay
2.8. Statistics
3. Results
3.1. Concentration-Dependent Increase in Cx43 Dye Coupling in Astrocytes Exposed to Danegaptide
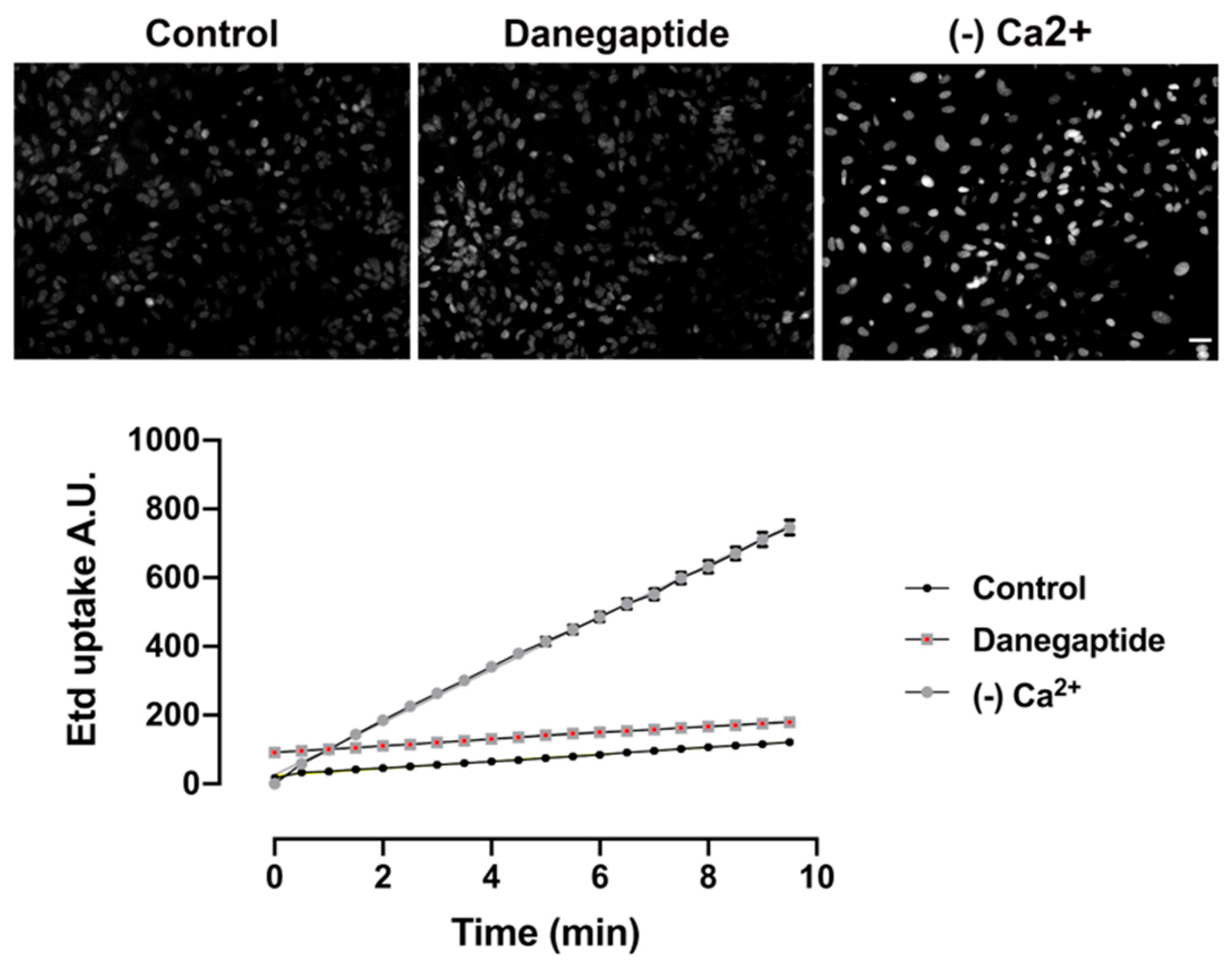
3.2. Danegaptide Does Not Affect Hemichannel Activity in Astrocytes In Vitro
3.3. MALDI IMS Was Able to Detect the Presence of Danegaptide in Brain Tissue
3.4. Danegaptide Reduced Infarct Volume in Mice Subjected to Ischemia/Reperfusion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Del Zoppo, G.J.; Sharp, F.R.; Heiss, W.D.; Albers, G.W. Heterogeneity in the penumbra. J. Cereb. Blood Flow Metab. 2011, 31, 1836–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.Z.; Peeling, J.; Sutherland, G.R.; Hertzberg, E.L.; Nagy, J.I. Ischemia-induced cellular redistribution of the astrocytic gap junctional protein connexin43 in rat brain. Brain Res. 1994, 652, 311–322. [Google Scholar] [CrossRef]
- Nakase, T.; Maeda, T.; Yoshida, Y.; Nagata, K. Ischemia alters the expression of connexins in the aged human brain. J. Biomed. Biotech. 2009, 2009, 147946. [Google Scholar] [CrossRef] [PubMed]
- Nakase, T.; Yoshida, Y.; Nagata, K. Enhanced connexin 43 immunoreactivity in penumbral areas in the human brain following ischemia. Glia 2006, 54, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Andrade, M.; She, J.; Bechberger, J.; Naus, C.C.; Sin, W.C. Acute connexin43 temporal and spatial expression in response to ischemic stroke. J. Cell Commun. Signal. 2018, 12, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Siushansian, R.; Bechberger, J.F.; Cechetto, D.F.; Hachinski, V.C.; Naus, C.C. Connexin43 null mutation increases infarct size after stroke. J. Comp. Neurol. 2001, 440, 387–394. [Google Scholar] [CrossRef]
- Kozoriz, M.G.; Bechberger, J.F.; Bechberger, G.R.; Suen, M.W.; Moreno, A.P.; Maass, K.; Willecke, K.; Naus, C.C. The connexin43 C-terminal region mediates neuroprotection during stroke. J. Neuropathol. Exp. Neurol. 2010, 69, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Nakase, T.; Fushiki, S.; Naus, C.C. Astrocytic gap junctions composed of connexin 43 reduce apoptotic neuronal damage in cerebral ischemia. Stroke 2003, 34, 1987–1993. [Google Scholar] [CrossRef] [Green Version]
- Naus, C.C.; Ozog, M.A.; Bechberger, J.F.; Nakase, T. A neuroprotective role for gap junctions. Cell Commun. Adhes. 2001, 8, 325–328. [Google Scholar] [CrossRef]
- Naus, C.C.; Bechberger, J.F.; Caveney, S.; Wilson, J.X. Expression of gap junction genes in astrocytes and C6 glioma cells. Neurosci. Lett. 1991, 126, 33–36. [Google Scholar] [CrossRef]
- Giaume, C. Communicating junctions of glial cells in the central nervous system. Annales d’endocrinologie 1996, 57, 487–491. [Google Scholar] [PubMed]
- Nagy, J.I.; Ionescu, A.V.; Lynn, B.D.; Rash, J.E. Coupling of astrocyte connexins Cx26, Cx30, Cx43 to oligodendrocyte Cx29, Cx32, Cx47: Implications from normal and connexin32 knockout mice. Glia 2003, 44, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naus, C.C.; Giaume, C. Bridging the gap to therapeutic strategies based on connexin/pannexin biology. J. Transl. Med. 2016, 14, 330. [Google Scholar] [CrossRef] [Green Version]
- Giaume, C.; Leybaert, L.; Naus, C.C.; Saez, J.C. Connexin and pannexin hemichannels in brain glial cells: Properties, pharmacology, and roles. Front. Pharmacol. 2013, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Hansson, E.; Muyderman, H.; Leonova, J.; Allansson, L.; Sinclair, J.; Blomstrand, F.; Thorlin, T.; Nilsson, M.; Ronnback, L. Astroglia and glutamate in physiology and pathology: Aspects on glutamate transport, glutamate-induced cell swelling and gap-junction communication. Neurochem. Int. 2000, 37, 317–329. [Google Scholar] [CrossRef]
- Giaume, C.; Koulakoff, A.; Roux, L.; Holcman, D.; Rouach, N. Astroglial networks: A step further in neuroglial and gliovascular interactions. Nat. Rev. Neurosci. 2010, 11, 87–99. [Google Scholar] [CrossRef]
- Ben Haim, L.; Rowitch, D.H. Functional diversity of astrocytes in neural circuit regulation. Nat. Rev. Neurosci. 2017, 18, 31–41. [Google Scholar] [CrossRef]
- Nakase, T.; Sohl, G.; Theis, M.; Willecke, K.; Naus, C.C. Increased apoptosis and inflammation after focal brain ischemia in mice lacking connexin43 in astrocytes. Am. J. Pathol. 2004, 164, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Shinotsuka, T.; Yasui, M.; Nuriya, M. Astrocytic gap junctional networks suppress cellular damage in an in vitro model of ischemia. Biochem. Biophys. Res. Commun. 2014, 444, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Bellot-Saez, A.; Kekesi, O.; Morley, J.W.; Buskila, Y. Astrocytic modulation of neuronal excitability through K+ spatial buffering. Neurosci. Biobehav. Rev. 2017, 77, 87–97. [Google Scholar] [CrossRef]
- Retamal, M.A.; Froger, N.; Palacios-Prado, N.; Ezan, P.; Saez, P.J.; Saez, J.C.; Giaume, C. Cx43 hemichannels and gap junction channels in astrocytes are regulated oppositely by proinflammatory cytokines released from activated microglia. J. Neurosci. 2007, 27, 13781–13792. [Google Scholar] [CrossRef] [PubMed]
- Froger, N.; Orellana, J.A.; Calvo, C.F.; Amigou, E.; Kozoriz, M.G.; Naus, C.C.; Saez, J.C.; Giaume, C. Inhibition of cytokine-induced connexin43 hemichannel activity in astrocytes is neuroprotective. Mol. Cell Neurosci. 2010, 45, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Orellana, J.A.; Froger, N.; Ezan, P.; Jiang, J.X.; Bennett, M.V.; Naus, C.C.; Giaume, C.; Saez, J.C. ATP and glutamate released via astroglial connexin 43 hemichannels mediate neuronal death through activation of pannexin 1 hemichannels. J. Neurochem. 2011, 118, 826–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, S.A.; Kondo, R.; Wang, S.Y.; Goldhaber, J.I.; Weiss, J.N. Connexin-43 hemichannels opened by metabolic inhibition. J. Biol. Chem. 1999, 274, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Orellana, J.A.; Hernandez, D.E.; Ezan, P.; Velarde, V.; Bennett, M.V.; Giaume, C.; Saez, J.C. Hypoxia in high glucose followed by reoxygenation in normal glucose reduces the viability of cortical astrocytes through increased permeability of connexin 43 hemichannels. Glia 2010, 58, 329–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas-Andrade, M.; Wang, N.; Bechberger, J.F.; De Bock, M.; Lampe, P.D.; Leybaert, L.; Naus, C.C. Targeting MAPK phosphorylation of Connexin43 provides neuroprotection in stroke. J. Exp. Med. 2019, 216, 916–935. [Google Scholar] [CrossRef]
- Chen, B.; Yang, L.; Chen, J.; Chen, Y.; Zhang, L.; Wang, L.; Li, X.; Li, Y.; Yu, H. Inhibition of Connexin43 hemichannels with Gap19 protects cerebral ischemia/reperfusion injury via the JAK2/STAT3 pathway in mice. Brain Res. Bull. 2019, 146, 124–135. [Google Scholar] [CrossRef]
- Butera, J.A.; Larsen, B.D.; Hennan, J.K.; Kerns, E.; Di, L.; Alimardanov, A.; Swillo, R.E.; Morgan, G.A.; Liu, K.; Wang, Q.; et al. Discovery of (2S,4R)-1-(2-aminoacetyl)-4-benzamidopyrrolidine-2-carboxylic acid hydrochloride (GAP-134)13, an orally active small molecule gap-junction modifier for the treatment of atrial fibrillation. J. Med. Chem. 2009, 52, 908–911. [Google Scholar] [CrossRef]
- Hennan, J.K.; Swillo, R.E.; Morgan, G.A.; Rossman, E.I.; Kantrowitz, J.; Butera, J.; Petersen, J.S.; Gardell, S.J.; Vlasuk, G.P. GAP-134 ([2S,4R]-1-[2-aminoacetyl]4-benzamidopyrrolidine-2-carboxylic acid) prevents spontaneous ventricular arrhythmias and reduces infarct size during myocardial ischemia/reperfusion injury in open-chest dogs. J. Cardiovasc. Pharmacol. Ther. 2009, 14, 207–214. [Google Scholar] [CrossRef]
- Skyschally, A.; Walter, B.; Schultz Hansen, R.; Heusch, G. The antiarrhythmic dipeptide ZP1609 (danegaptide) when given at reperfusion reduces myocardial infarct size in pigs. Naunyn. Schmiedebergs Arch. Pharmacol. 2013, 386, 383–391. [Google Scholar] [CrossRef]
- Clarke, T.C.; Thomas, D.; Petersen, J.S.; Evans, W.H.; Martin, P.E. The antiarrhythmic peptide rotigaptide (ZP123) increases gap junction intercellular communication in cardiac myocytes and HeLa cells expressing connexin 43. Br. J. Pharmacol. 2006, 147, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Mouritzen, U.; Larsen, B.D.; Roy, S. Inhibition of Cx43 gap junction uncoupling prevents high glucose-induced apoptosis and reduces excess cell monolayer permeability in retinal vascular endothelial cells. Exp. Eye Res. 2018, 173, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Clarke, D.; Al Ahmad, A.; Kahle, M.; Parham, C.; Auckland, L.; Shaw, C.; Fidanboylu, M.; Orr, A.W.; Ogunshola, O.; et al. Perlecan domain V is neuroprotective and proangiogenic following ischemic stroke in rodents. J. Clin. Investig. 2011, 121, 3005–3023. [Google Scholar] [CrossRef] [Green Version]
- Maniskas, M.; Bix, G.; Fraser, J. Selective intra-arterial drug administration in a model of large vessel ischemia. J. Neurosci. Methods 2015, 240, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, A.; Graham, D.I.; McCulloch, J.; Teasdale, G.M. Focal cerebral ischaemia in the rat: 1. Description of technique and early neuropathological consequences following middle cerebral artery occlusion. J. Cereb. Blood Flow Metab. 1981, 1, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodard, G. Principles of drug administration. In Methods of Animal Experimentation; Gay, W.J., Ed.; Academic: New York, NY, USA, 1965; Volume 1, pp. 343–359. [Google Scholar]
- Vogel, J.; Mobius, C.; Kuschinsky, W. Early delineation of ischemic tissue in rat brain cryosections by high-contrast staining. Stroke 1999, 30, 1134–1141. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.N.; He, Y.Y.; Wu, G.; Khan, M.; Hsu, C.Y. Effect of brain edema on infarct volume in a focal cerebral ischemia model in rats. Stroke 1993, 24, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Le, H.T.; Sin, W.C.; Lozinsky, S.; Bechberger, J.; Vega, J.L.; Guo, X.Q.; Saez, J.C.; Naus, C.C. Gap junction intercellular communication mediated by connexin43 in astrocytes is essential for their resistance to oxidative stress. J. Biol. Chem. 2014, 289, 1345–1354. [Google Scholar] [CrossRef] [Green Version]
- Kozoriz, M.G.; Lai, S.; Vega, J.L.; Saez, J.C.; Sin, W.C.; Bechberger, J.F.; Naus, C.C. Cerebral ischemic injury is enhanced in a model of oculodentodigital dysplasia. Neuropharmacology 2013, 75, 549–556. [Google Scholar] [CrossRef]
- Freitas-Andrade, M.; Carmeliet, P.; Charlebois, C.; Stanimirovic, D.B.; Moreno, M.J. PlGF knockout delays brain vessel growth and maturation upon systemic hypoxic challenge. J. Cereb. Blood Flow Metab. 2012, 32, 663–675. [Google Scholar] [CrossRef]
- Laurent, G.; Leong-Poi, H.; Mangat, I.; Moe, G.W.; Hu, X.; So, P.P.; Tarulli, E.; Ramadeen, A.; Rossman, E.I.; Hennan, J.K.; et al. Effects of chronic gap junction conduction-enhancing antiarrhythmic peptide GAP-134 administration on experimental atrial fibrillation in dogs. Circ. Arrhythm. Electrophysiol. 2009, 2, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, A.; Dermietzel, R. Visualization and functional blocking of gap junction hemichannels (connexons) with antibodies against external loop domains in astrocytes. Glia 1998, 24, 141–154. [Google Scholar] [CrossRef]
- Wang, J.S.H.; Freitas-Andrade, M.; Bechberger, J.F.; Naus, C.C.; Yeung, K.K.; Whitehead, S.N. Matrix-assisted laser desorption/ionization imaging mass spectrometry of intraperitoneally injected danegaptide (ZP1609) for treatment of stroke-reperfusion injury in mice. Rapid Commun. Mass Spectrom. 2018, 32, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Amdisen, C.; Keller, A.K.; Hansen, R.S.; Norregaard, R.; Krag, S.P.; Moldrup, U.; Pedersen, M.; Jespersen, B.; Birn, H. Testing Danegaptide Effects on Kidney Function after Ischemia/Reperfusion Injury in a New Porcine Two Week Model. PLoS ONE 2016, 11, e0164109. [Google Scholar] [CrossRef]
- Boengler, K.; Bulic, M.; Schreckenberg, R.; Schluter, K.D.; Schulz, R. The gap junction modifier ZP1609 decreases cardiomyocyte hypercontracture following ischaemia/reperfusion independent from mitochondrial connexin 43. Br. J. Pharmacol. 2017, 174, 2060–2073. [Google Scholar] [CrossRef]
- Laird, D.W.; Lampe, P.D. Therapeutic strategies targeting connexins. Nat. Rev. Drug Discov. 2018. [Google Scholar] [CrossRef]
- Martinez, A.D.; Saez, J.C. Regulation of astrocyte gap junctions by hypoxia-reoxygenation. Brain Res. Rev 2000, 32, 250–258. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas-Andrade, M.; Bechberger, J.; Wang, J.; Yeung, K.K.C.; Whitehead, S.N.; Shultz Hansen, R.; Naus, C.C. Danegaptide Enhances Astrocyte Gap Junctional Coupling and Reduces Ischemic Reperfusion Brain Injury in Mice. Biomolecules 2020, 10, 353. https://doi.org/10.3390/biom10030353
Freitas-Andrade M, Bechberger J, Wang J, Yeung KKC, Whitehead SN, Shultz Hansen R, Naus CC. Danegaptide Enhances Astrocyte Gap Junctional Coupling and Reduces Ischemic Reperfusion Brain Injury in Mice. Biomolecules. 2020; 10(3):353. https://doi.org/10.3390/biom10030353
Chicago/Turabian StyleFreitas-Andrade, Moises, John Bechberger, Jasmine Wang, Ken K.C. Yeung, Shawn N. Whitehead, Rie Shultz Hansen, and Christian C. Naus. 2020. "Danegaptide Enhances Astrocyte Gap Junctional Coupling and Reduces Ischemic Reperfusion Brain Injury in Mice" Biomolecules 10, no. 3: 353. https://doi.org/10.3390/biom10030353
APA StyleFreitas-Andrade, M., Bechberger, J., Wang, J., Yeung, K. K. C., Whitehead, S. N., Shultz Hansen, R., & Naus, C. C. (2020). Danegaptide Enhances Astrocyte Gap Junctional Coupling and Reduces Ischemic Reperfusion Brain Injury in Mice. Biomolecules, 10(3), 353. https://doi.org/10.3390/biom10030353