Src/NF-κB-Targeted Anti-Inflammatory Effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. Ethanol Extract

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Cell Culture
2.4. Cell Viability Test
2.5. Nitric Oxide (NO) Assay
2.6. High-Performance Liquid Chromatography (HPLC)
2.7. mRNA Analysis by a Quantitative Reverse Transcriptase-Polymerase Chain Reaction
2.8. Plasmid Transfection and Luciferase Reporter Gene Activity Assay
2.9. Western Blot Analysis and Immunoprecipitation
2.10. Cellular Thermal Shift Assay (CETSA)
2.11. HCl/EtOH-Induced Gastritis Model
2.12. Statistical Analysis
3. Results
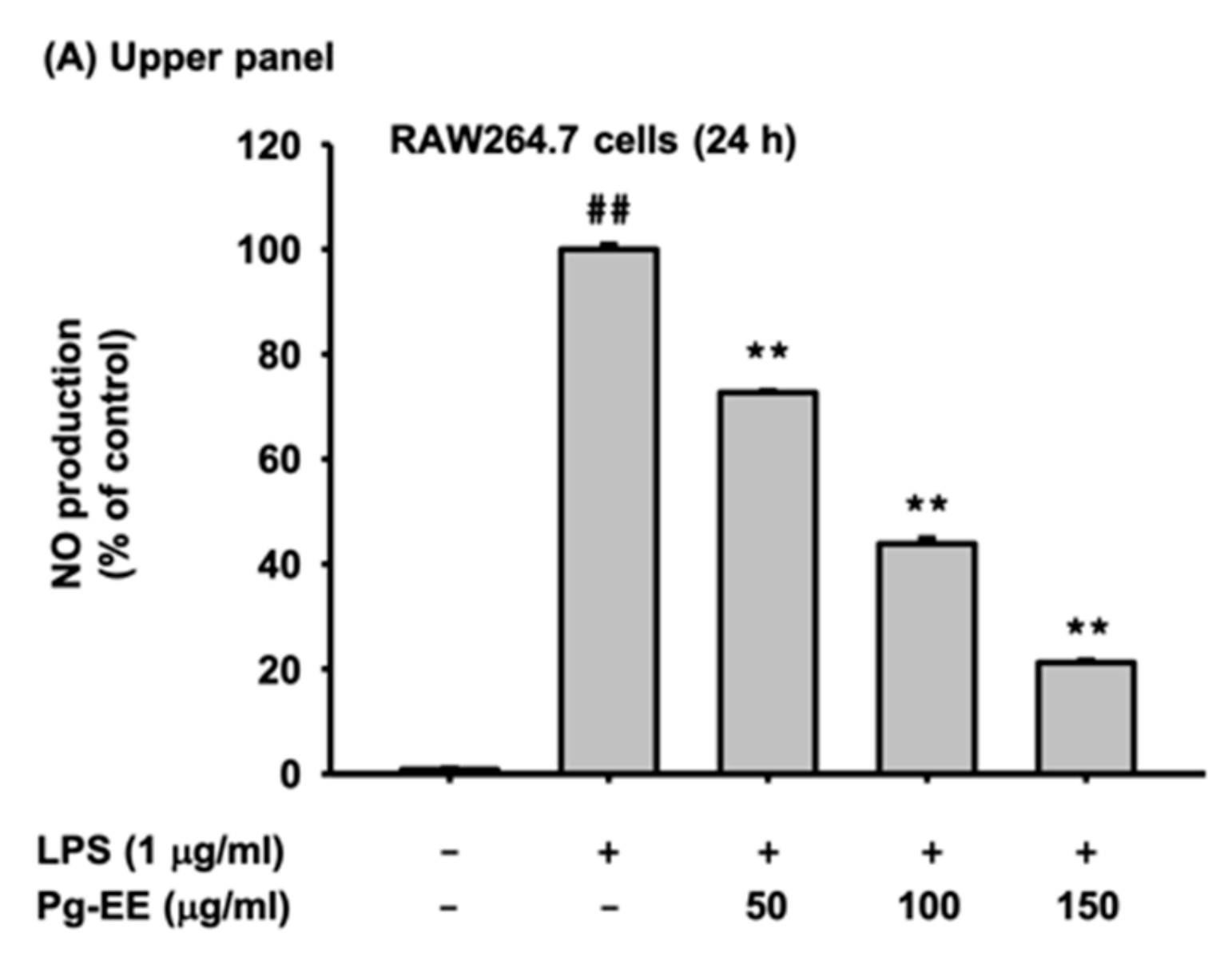
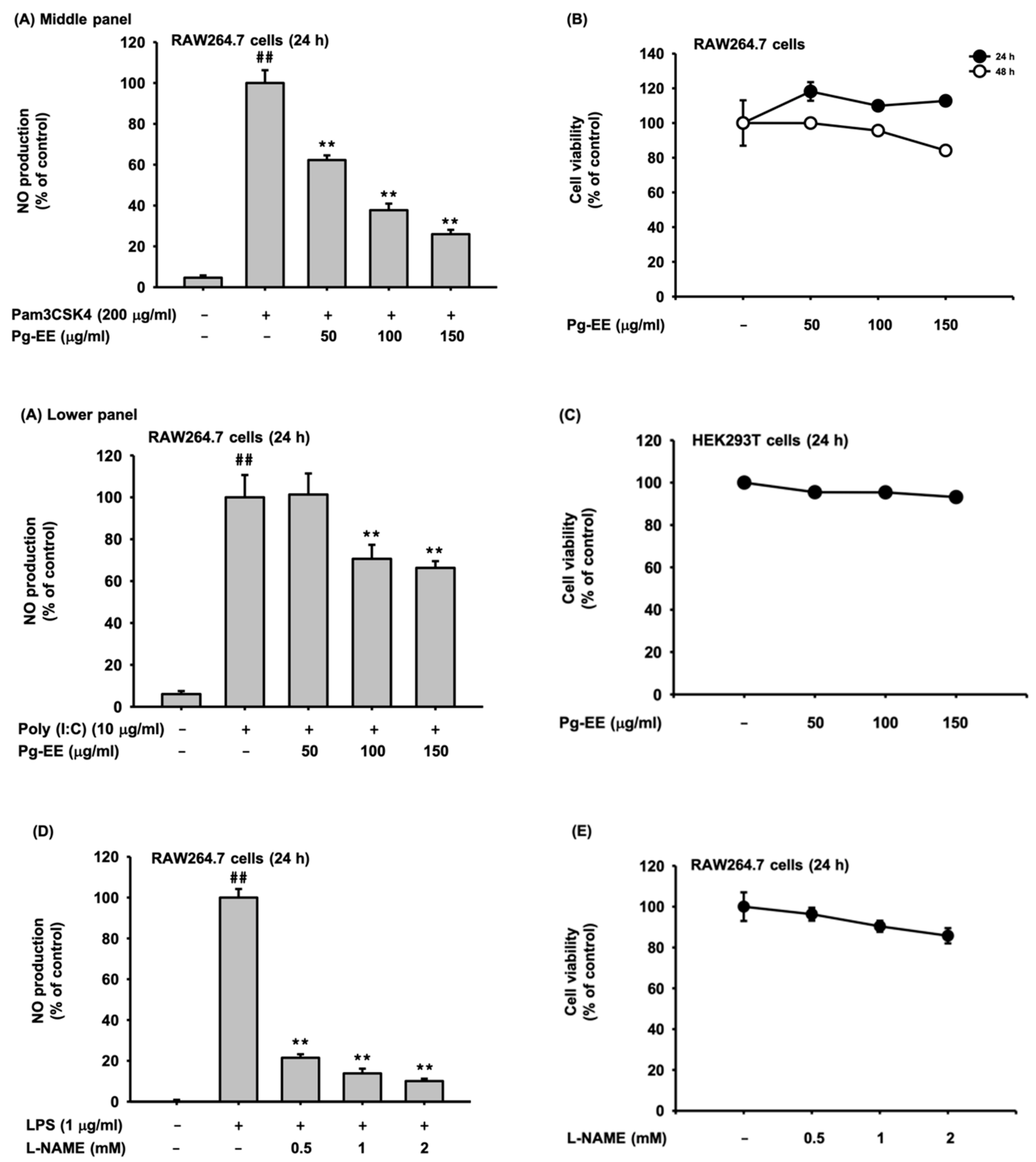
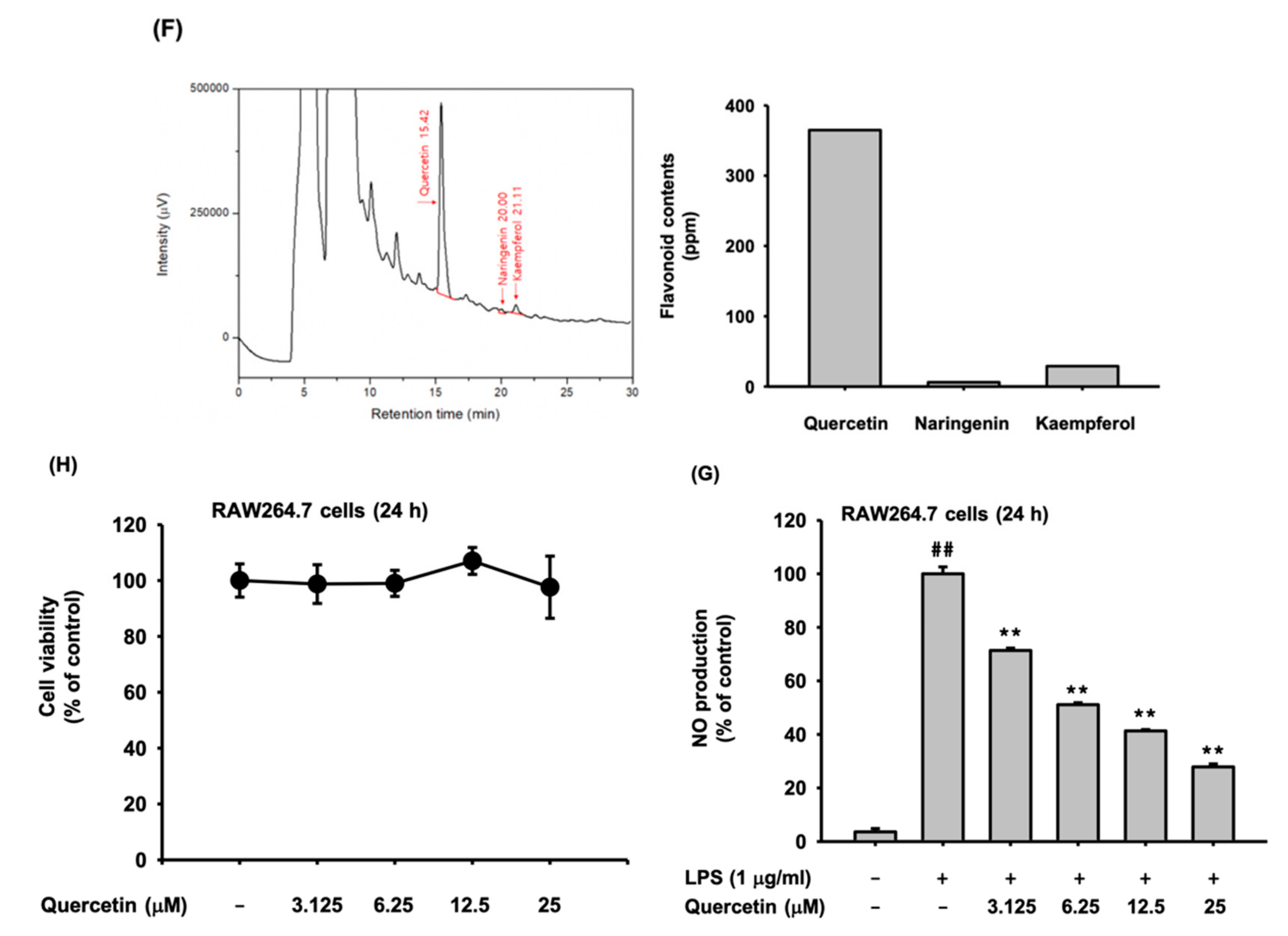
3.1. Pg-EE Suppressed the NO Production Level
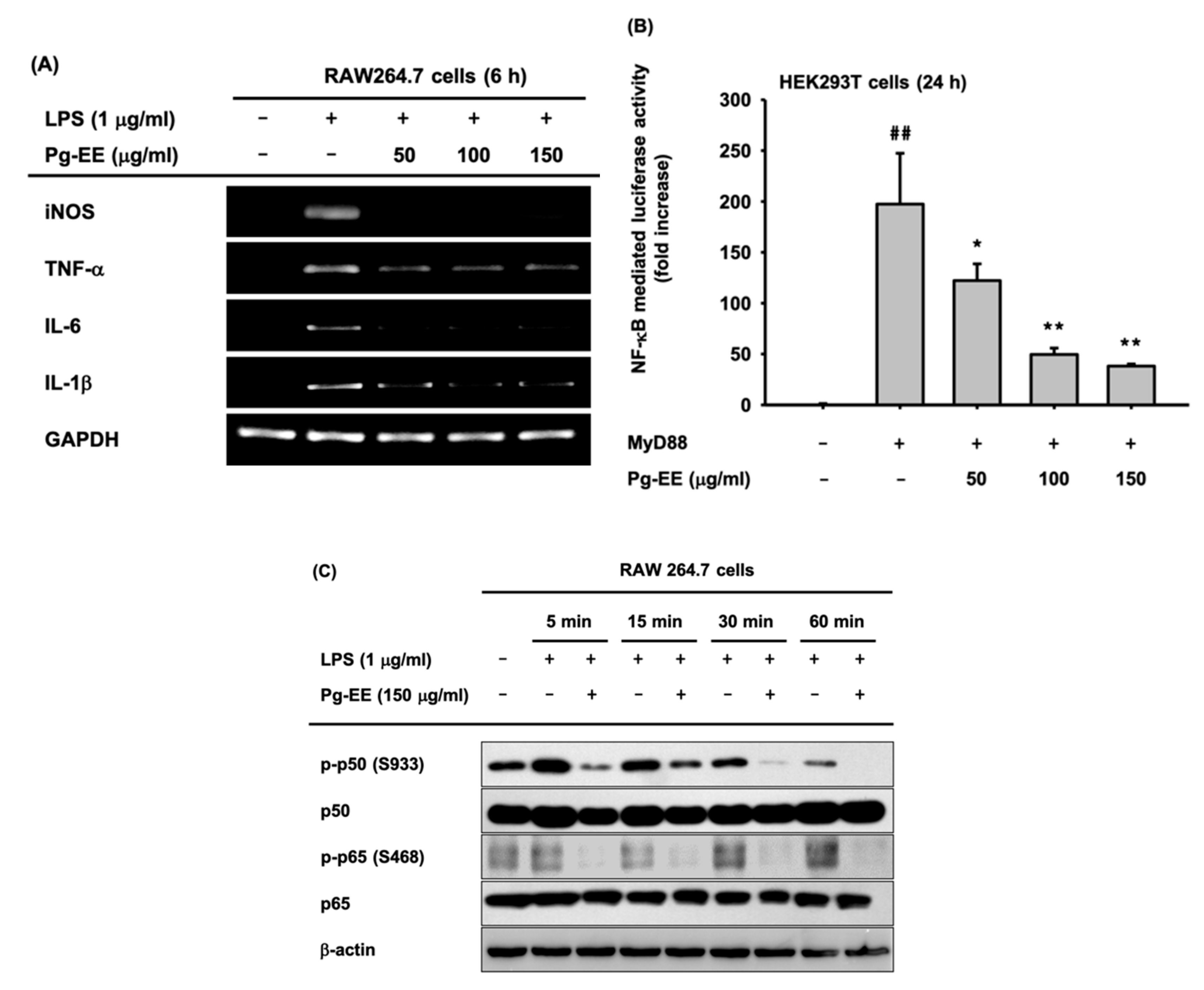
3.2. Anti-Inflammatory Effects of Pg-EE at the Transcriptional Level
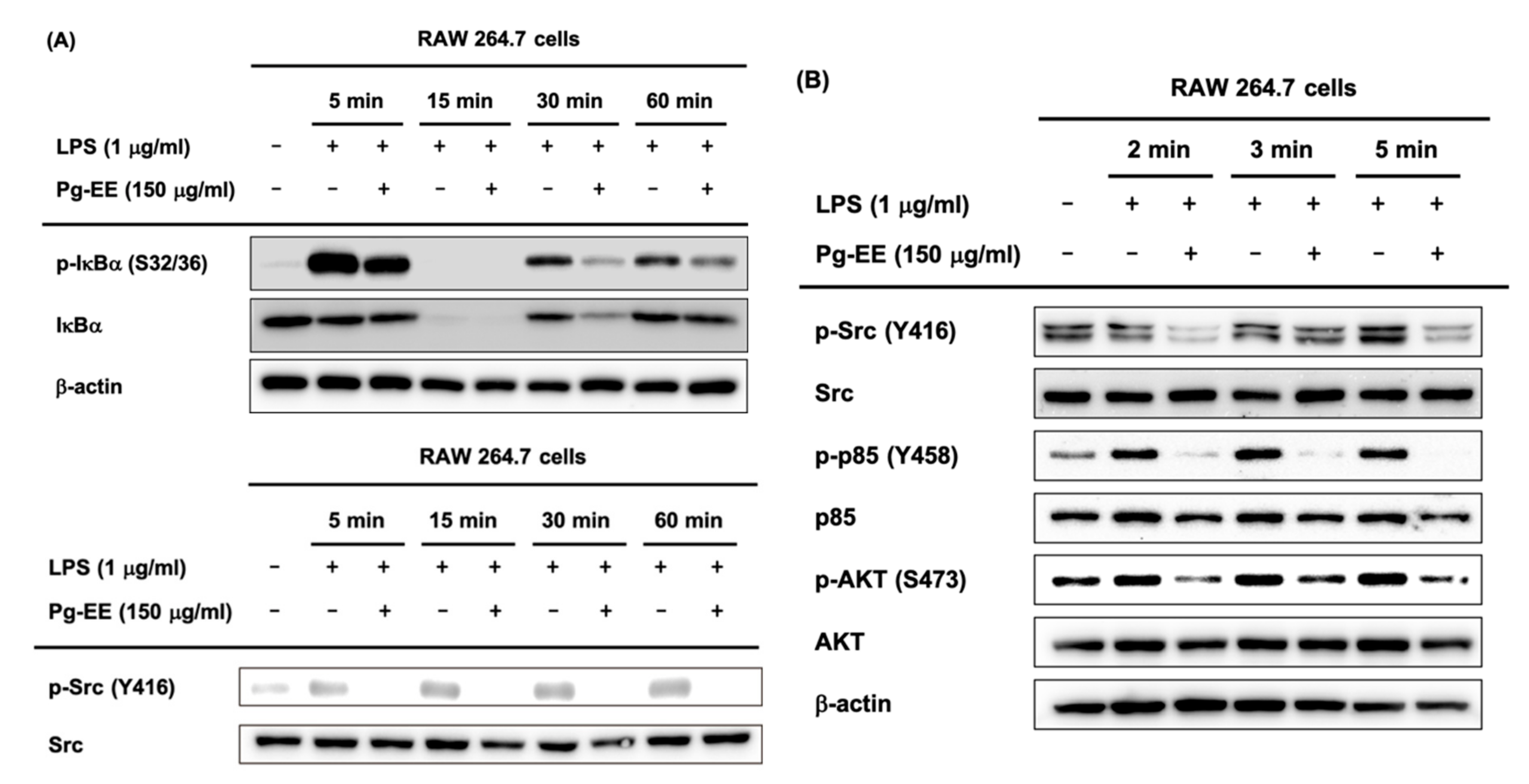
3.3. Regulatory Mechanism of Pg-EE in NF-κB Pathways
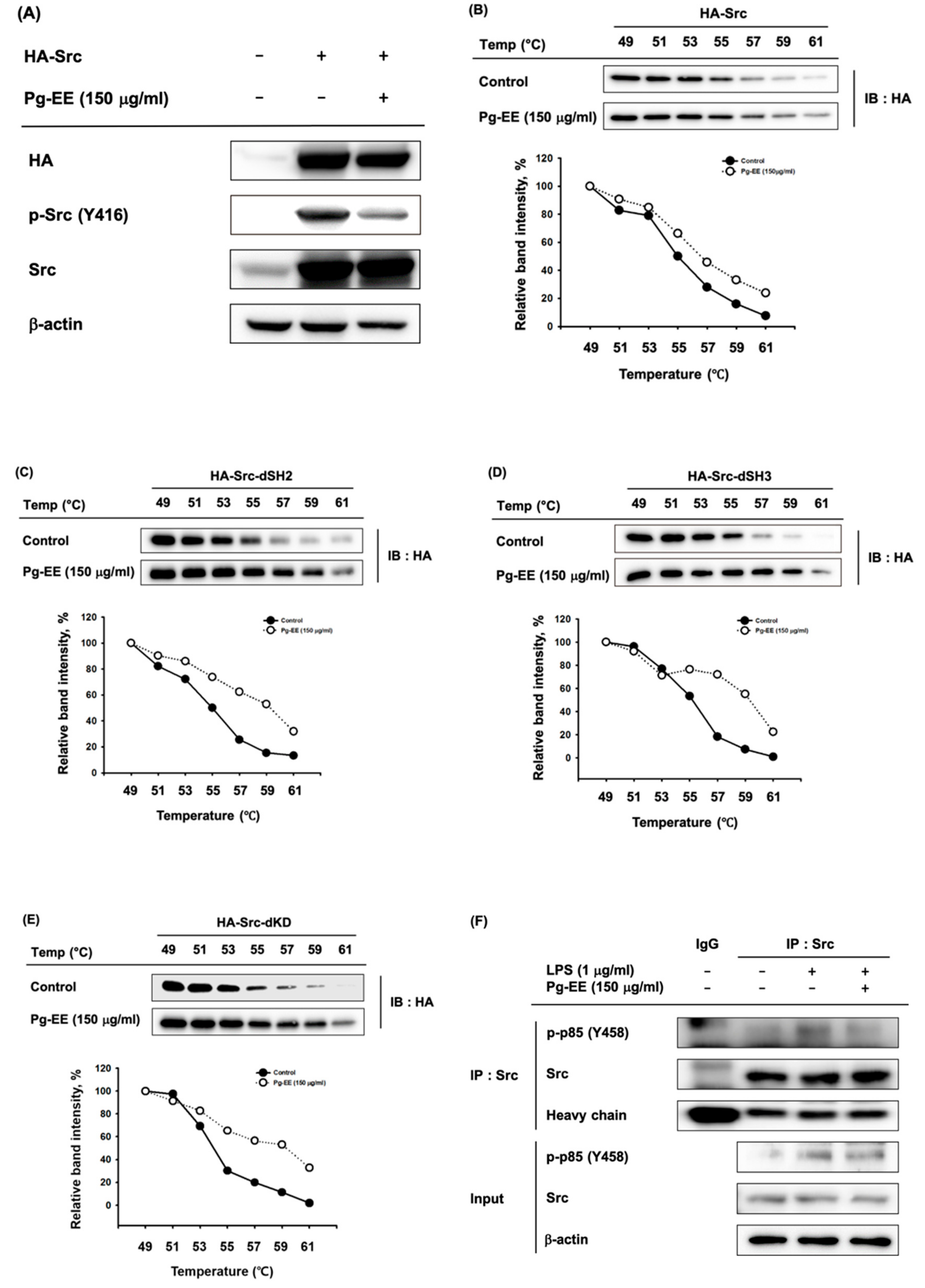
3.4. Anti-Inflammatory Effects of Pg-EE by Targeting Src Kinases
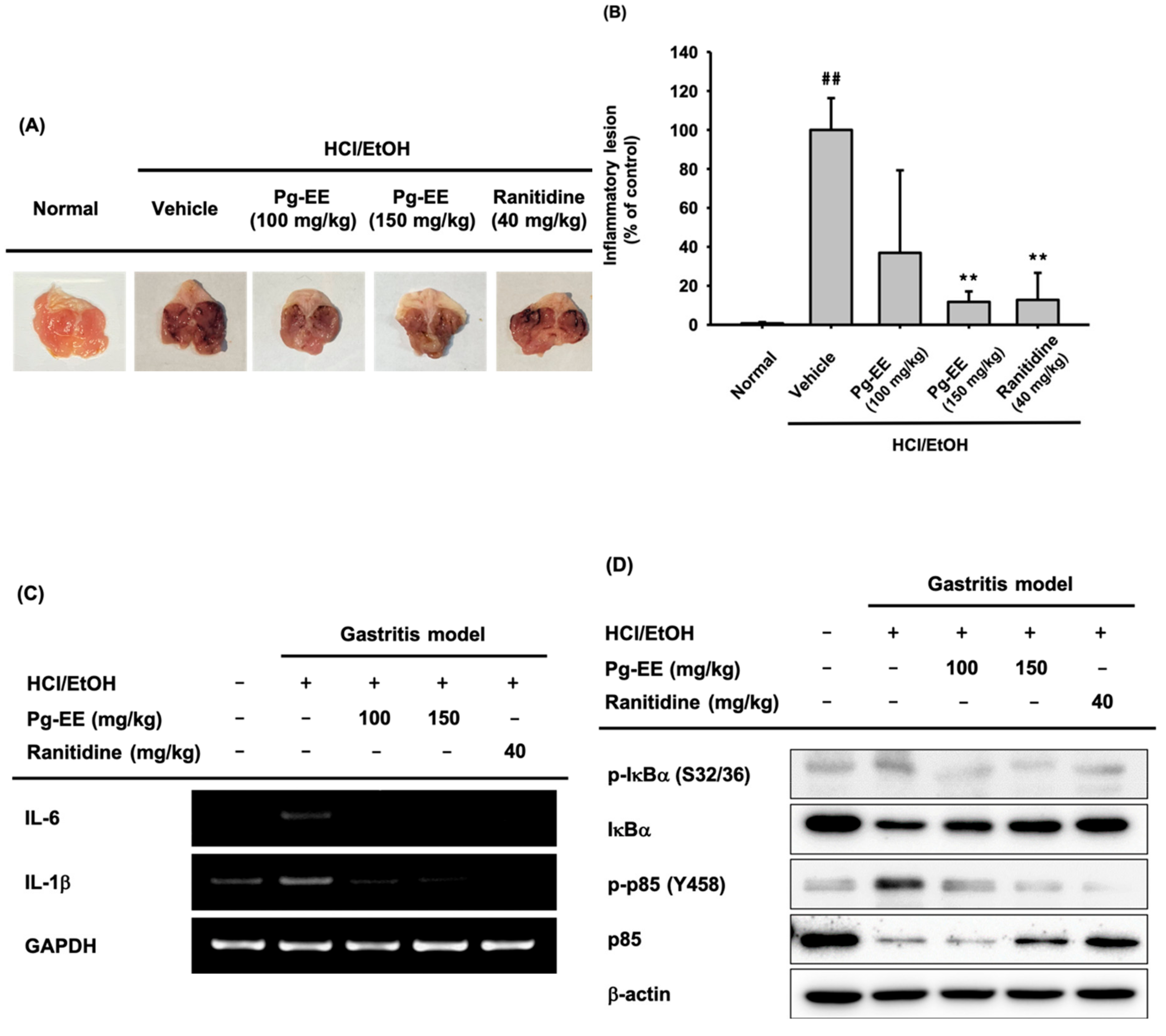
3.5. In Vivo Anti-Inflammatory Effect of Pg-EE on HCl/EtOH-Induced Acute Gastritis
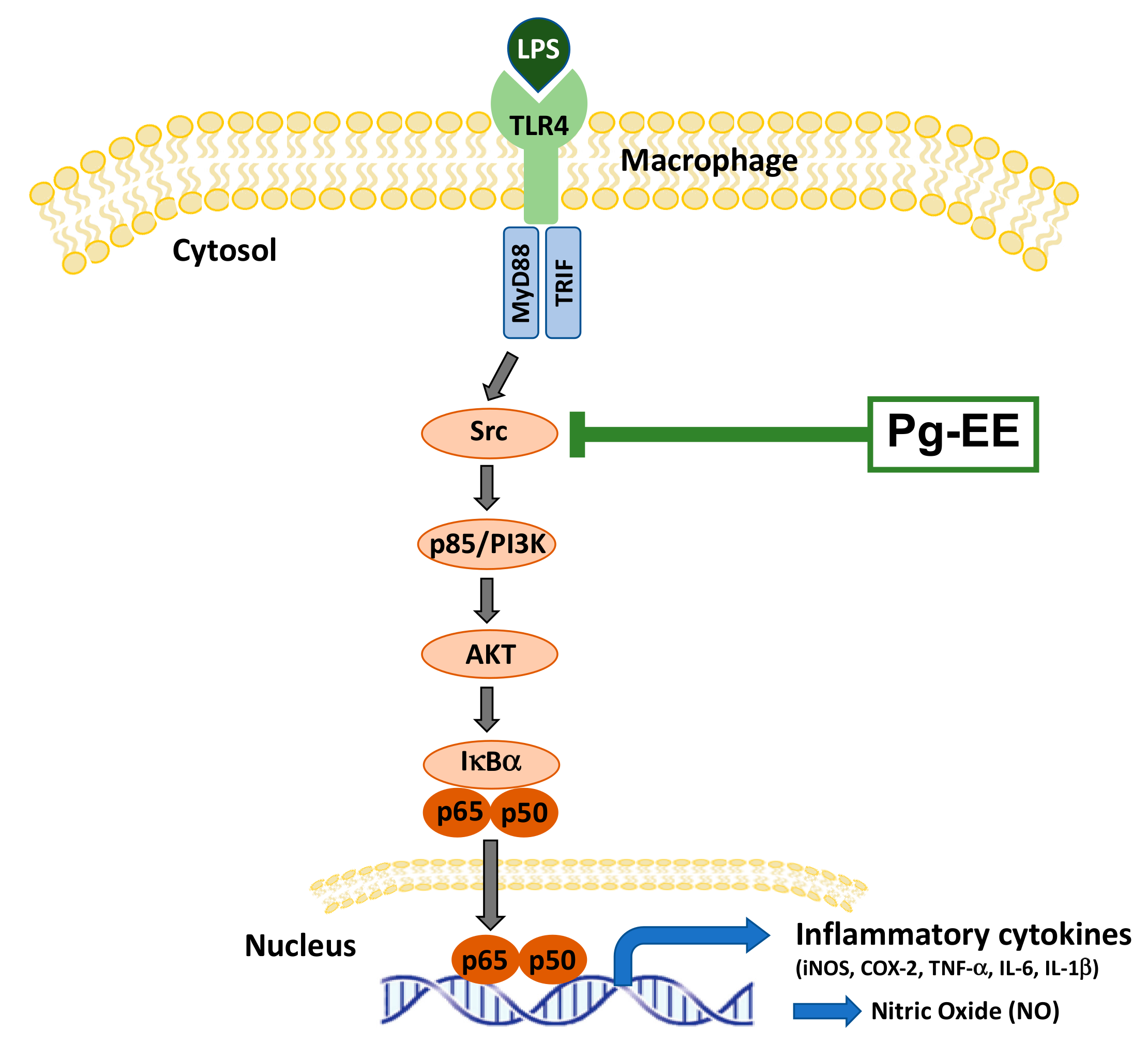
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations/Nomenclature
| Activator protein 1 | AP-1 |
| Cyclooxygenase-2 | COX-2 |
| Inducible NO synthase | iNOS |
| Inhibitor of kappa B alpha | IκBα |
| Inhibitor of kappa B kinase | IKK |
| Interleukin | IL |
| Interleukin | IL |
| Lipopolysaccharide | LPS |
| Myeloid differentiation primary response 88 | MYD88 |
| Nitric oxide | NO |
| Nuclear factor kappa-light-chain-enhancer of activated B cells | NF-κB |
| Pathogen-associated molecular patterns | PAMPs |
| Pattern recognition receptors | PRRs |
| Phosphoinositide-3-kinase p85 | PI3K p85 |
| Potentilla glabra Var. mandshurica (Maxim.) Hand.-Mazz ethanol extract | Pg-EE |
| Spleen tyrosine kinase | Syk |
| TIR-domain-containing adapter-inducing interferon-β | TRIF |
| Toll-like receptors | TLRs |
| Tumor necrosis factor-alpha | TNF-α |
References
- Charles, A.; Janeway, J.; Medzhitov, R. Innate Immune Recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero-Miliani, L.; Nielsen, O.; Andersen, P.; Girardin, S. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1β generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Yi, Y.-S.; Oh, J.; Yoo, B.C.; Hong, S.; Cho, J.Y. The role of Src kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2012, 2012, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiró, N.; Vizoso, F.J. Inflammation and cancer. World J. Gastrointest. Surg. 2012, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C., Jr. Innate immunity. New Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef]
- Janeway, C.A. Approaching the asymptote? Evolution and revolution in immunology. In Proceedings of the Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; pp. 1–13. [Google Scholar]
- Jin, M.S.; Lee, J.-O. Structures of TLR–ligand complexes. Curr. Opin. Immunol. 2008, 20, 414–419. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Ritter, M.; Mennerich, D.; Weith, A.; Seither, P. Characterization of Toll-like receptors in primary lung epithelial cells: Strong impact of the TLR3 ligand poly (I: C) on the regulation of Toll-like receptors, adaptor proteins and inflammatory response. J. Inflamm. 2005, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Lucas, K.; Maes, M. Role of the Toll Like receptor (TLR) radical cycle in chronic inflammation: Possible treatments targeting the TLR4 pathway. Mol. Neurobiol. 2013, 48, 190–204. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granado-Serrano, A.B.; Martín, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin induces NF-κB, activator protein-1 (AP-1) and nuclear transcription factor erythroid 2p45-related factor-2 (Nrf2) via phosphatidylinositol-3-kinase/protein kinase B (PI3K/AKT) and extracellular regulated kinase (ERK) signalling in HepG2 cells. Br. J. Nutr. 2010, 103, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2018, 42, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Farlik, M.; Reutterer, B.; Schindler, C.; Greten, F.; Vogl, C.; Müller, M.; Decker, T. Nonconventional initiation complex assembly by STAT and NF-κB transcription factors regulates nitric oxide synthase expression. Immunity 2010, 33, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calixto, J.B.; Campos, M.M.; Otuki, M.F.; Santos, A.R. Anti-inflammatory compounds of plant origin. Part II. Modulation of pro-inflammatory cytokines, chemokines and adhesion molecules. Planta Med. 2004, 70, 93–103. [Google Scholar] [PubMed] [Green Version]
- Lee, J.O.; Choi, E.; Shin, K.K.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Hossain, M.A.; Kim, H.S.; Yi, Y.S.; Kim, D.; et al. Compound K, a ginsenoside metabolite, plays an antiinflammatory role in macrophages by targeting the AKT1-mediated signaling pathway. J. Ginseng Res. 2019, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef] [Green Version]
- Ninnemann, J.L. Prostaglandins in inflammation and disease. Immunol. Today 1984, 5, 173–175. [Google Scholar] [CrossRef]
- Vogl, S.; Picker, P.; Mihaly-Bison, J.; Fakhrudin, N.; Atanasov, A.G.; Heiss, E.H.; Wawrosch, C.; Reznicek, G.; Dirsch, V.M.; Saukel, J. Ethnopharmacological in vitro studies on Austria’s folk medicine—An unexplored lore in vitro anti-inflammatory activities of 71 Austrian traditional herbal drugs. J. Ethnopharmacol. 2013, 149, 750–771. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, W.S.; Yu, T.; Sung, G.-H.; Park, K.W.; Yoon, K.; Son, Y.-J.; Hwang, H.; Kwak, Y.-S.; Lee, C.-M. ATF-2/CREB/IRF-3-targeted anti-inflammatory activity of Korean red ginseng water extract. J. Ethnopharmacol. 2014, 154, 218–228. [Google Scholar] [CrossRef]
- Tomczyk, M.; Paduch, R.; Wiater, A.; Pleszczyńska, M.; Kandefer-Szerszeń, M.; Szczodrak, J. The influence of aqueous extracts of selected Potentilla species on normal human colon cells. Acta Pol. Pharm. 2013, 70, 523–531. [Google Scholar] [PubMed]
- Leporatti, M.L.; Ivancheva, S. Preliminary comparative analysis of medicinal plants used in the traditional medicine of Bulgaria and Italy. J. Ethnopharmacol. 2003, 87, 123–142. [Google Scholar] [CrossRef]
- Wang, S.-S.; Wang, D.-M.; Pu, W.-J.; Li, D.-W. Phytochemical profiles, antioxidant and antimicrobial activities of three Potentilla species. BMC Complementary Altern. Med. 2013, 13, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng Res. 2019, 43, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Starkenmann, C.; Luca, L.; Niclass, Y.; Praz, E.; Roguet, D. Comparison of volatile constituents of Persicaria odorata (Lour.) Sojak (Polygonum odoratum Lour.) and Persicaria hydropiper L. Spach (Polygonum hydropiper L.). J. Agric. Food Chem. 2006, 54, 3067–3071. [Google Scholar] [CrossRef]
- Hossen, M.J.; Jeon, S.H.; Kim, S.C.; Kim, J.H.; Jeong, D.; Sung, N.Y.; Yang, S.; Baek, K.-S.; Kim, J.H.; Yoon, D.H. In vitro and in vivo anti-inflammatory activity of Phyllanthus acidus methanolic extract. J. Ethnopharmacol. 2015, 168, 217–228. [Google Scholar] [CrossRef]
- Yang, W.S.; Yi, Y.-S.; Kim, D.; Kim, M.H.; Park, J.G.; Kim, E.; Lee, S.Y.; Yoon, K.; Kim, J.-H.; Park, J. Nuclear factor kappa-B-and activator protein-1-mediated immunostimulatory activity of compound K in monocytes and macrophages. J. Ginseng Res. 2017, 41, 298–306. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, D.; Nam, G.; Yoo, S.; Han, S.Y.; Jeong, S.-G.; Kim, E.; Jeong, D.; Yoon, K.; Kim, S. Photoaging protective effects of BIOGF1K, a compound-K-rich fraction prepared from Panax ginseng. J. Ginseng Res. 2018, 42, 81–89. [Google Scholar] [CrossRef]
- Arakawa, H.; Tsuji, A.; Maeda, M. Chemiluminescent assay of β-D-galactosidase based on indole luminescence. J. Biolumin. Chemilumin. 1998, 13, 349–354. [Google Scholar] [CrossRef]
- Kim, H.G.; Choi, S.; Lee, J.; Hong, Y.H.; Jeong, D.; Yoon, K.; Yoon, D.H.; Sung, G.-H.; Lee, S.; Hong, S. Src is a prime target inhibited by Celtis choseniana methanol extract in its anti-inflammatory action. Evid. Based Complement. Altern. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Liu, B.; Xu, Y.; Li, W.-L.; Zeng, L. Proteomic analysis of differentially expressed skin proteins in iRhom2Uncv mice. BMB Rep. 2015, 48, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.; Yi, Y.-S.; Sung, G.-H.; Yang, W.S.; Park, J.G.; Yoon, K.; Yoon, D.H.; Song, C.; Lee, Y.; Rhee, M.H. Anti-inflammatory activities and mechanisms of Artemisia asiatica ethanol extract. J. Ethnopharmacol. 2014, 152, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Rhee, M.H.; Lee, J.; Kim, S.H.; Yang, Y.; Kim, H.G.; Kim, Y.; Kim, C.; Kwak, Y.-S.; Kim, J.-H.; et al. Ginsenoside Rc from Korean red ginseng (Panax ginseng CA Meyer) attenuates inflammatory symptoms of gastritis, hepatitis and arthritis. Am. J. Chin. Med. 2016, 44, 595–615. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Park, S.-J.; Kwon, M.-J.; Jeong, T.-S.; Bok, S.-H.; Choi, W.-Y.; Jeong, W.-I.; Ryu, S.-Y.; Do, S.-H.; Lee, C.-S. Quercetin suppresses proinflammatory cytokines production through MAP kinases and NF-κB pathway in lipopolysaccharide-stimulated macrophage. Mol. Cell. Biochem. 2003, 243, 153–160. [Google Scholar] [CrossRef]
- Waseem, T.; Duxbury, M.; Ito, H.; Ashley, S.W.; Robinson, M.K. Exogenous ghrelin modulates release of pro-inflammatory and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 2008, 143, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-H.; Mandelker, D.; Gabelli, S.B.; Amzel, L.M. Insights into the oncogenic effects of/PIK3CA/mutations from the structure of p110α/p85α. Cell Cycle 2008, 7, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Daulhac, L.; Kowalski-Chauvel, A.; Pradayrol, L.; Vaysse, N.; Seva, C. Src-family tyrosine kinases in activation of ERK-1 and p85/p110-phosphatidylinositol 3-kinase by G/CCKB receptors. J. Biol. Chem. 1999, 274, 20657–20663. [Google Scholar] [CrossRef] [Green Version]
- Roskoski, R., Jr. Src protein-tyrosine kinase structure and regulation. Biochem. Biophys. Res. Commun. 2004, 324, 1155–1164. [Google Scholar] [CrossRef]
- Her, N.-G.; Babic, I.; Yenugonda, V.M.; Kesari, S.; Nurmemmedov, E. Cellular thermal shift analysis for interrogation of CRISPR-assisted proteomic changes. BioTechniques 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskoski, R., Jr. Src kinase regulation by phosphorylation and dephosphorylation. Biochem. Biophys. Res. Commun. 2005, 331, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.G., Jr.; Backer, J.M.; Sun, X.J.; Shoelson, S.; Hu, P.; Schlessinger, J.; Yoakim, M.; Schaffhausen, B.; White, M.F. IRS-1 activates phosphatidylinositol 3′-kinase by associating with src homology 2 domains of p85. Proc. Natl. Acad. Sci. USA 1992, 89, 10350–10354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264.7 macrophages and DNCB-induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [CrossRef]
- Xagorari, A.; Papapetropoulos, A.; Mauromatis, A.; Economou, M.; Fotsis, T.; Roussos, C. Luteolin inhibits an endotoxin-stimulated phosphorylation cascade and proinflammatory cytokine production in macrophages. J. Pharmacol. Exp. Ther. 2001, 296, 181–187. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences (5′ to 3′) | |
|---|---|---|
| iNOS | Forward | 5′-CCCTTCCGAAGTTTCTGGCAGCAG −3′ |
| Reverse | 5′-GGCTGTCAGAGCCTCGTGGCTTTGG −3′ | |
| TNF-α | Forward | 5′-TTGACCTCAGCGCTGAGTTG −3′ |
| Reverse | 5′-CCTGTAGCCCACGTCGTAGC −3′ | |
| IL-6 | Forward | 5′-GGAAATCGTGGAAATGAG −3′ |
| Reverse | 5′-GCTTAGGCATAACGCACT −3′ | |
| IL-1β | Forward | 5′-CAGGATGAGGACATGAGCACC −3′ |
| Reverse | 5′-CTCTGCAGACTCAAACTCCAC −3′ | |
| GAPDH (Mouse) | Forward | 5′-CACTCACGGCAAATTCAACGGCAC −3′ |
| Reverse | 5′-GACTCCACGACATACTCAGCAC −3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Shin, K.K.; Kim, H.G.; Jo, M.; Kim, J.K.; Lee, J.S.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, K.-H.; et al. Src/NF-κB-Targeted Anti-Inflammatory Effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. Ethanol Extract. Biomolecules 2020, 10, 648. https://doi.org/10.3390/biom10040648
Kim H, Shin KK, Kim HG, Jo M, Kim JK, Lee JS, Choung ES, Li WY, Lee SW, Kim K-H, et al. Src/NF-κB-Targeted Anti-Inflammatory Effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. Ethanol Extract. Biomolecules. 2020; 10(4):648. https://doi.org/10.3390/biom10040648
Chicago/Turabian StyleKim, Haeyeop, Kon Kuk Shin, Han Gyung Kim, Minkyeong Jo, Jin Kyeong Kim, Jong Sub Lee, Eui Su Choung, Wan Yi Li, Sang Woo Lee, Kyung-Hee Kim, and et al. 2020. "Src/NF-κB-Targeted Anti-Inflammatory Effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. Ethanol Extract" Biomolecules 10, no. 4: 648. https://doi.org/10.3390/biom10040648
APA StyleKim, H., Shin, K. K., Kim, H. G., Jo, M., Kim, J. K., Lee, J. S., Choung, E. S., Li, W. Y., Lee, S. W., Kim, K. -H., Yoo, B. C., & Cho, J. Y. (2020). Src/NF-κB-Targeted Anti-Inflammatory Effects of Potentilla glabra var. Mandshurica (Maxim.) Hand.-Mazz. Ethanol Extract. Biomolecules, 10(4), 648. https://doi.org/10.3390/biom10040648