Lignin and Quercetin Synthesis Underlies Berry Russeting in ‘Sunshine Muscat’ Grape


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Total RNA Extraction and Sequencing
2.3. Data Quality Control
2.4. Differentially Expressed Gene Analysis and Enrichment Analysis
2.5. qRT-PCR Analysis
2.6. Measurement and Analysis of Metabolomes
3. Results
3.1. Transcriptome Sequencing Yield Statistics
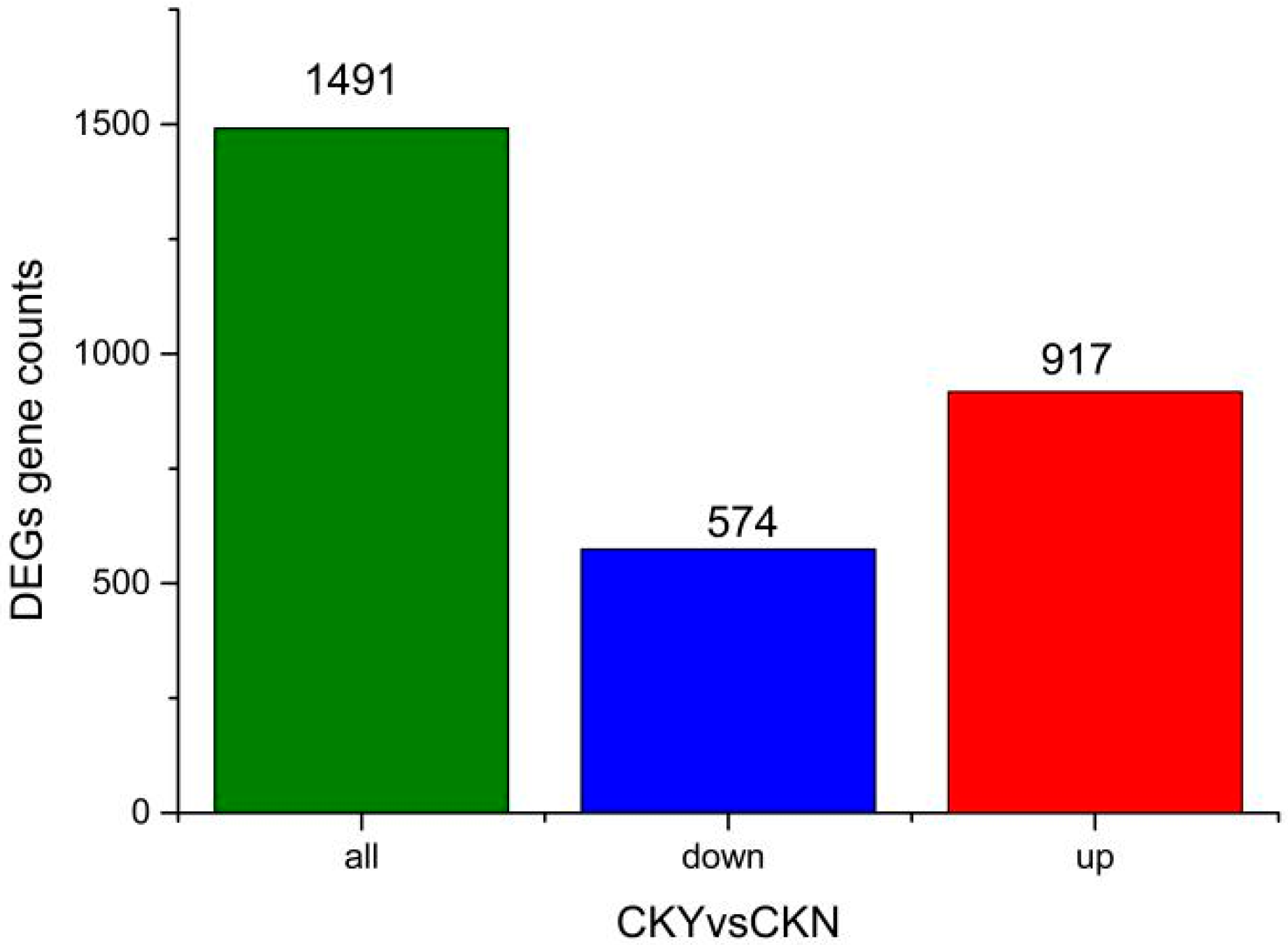
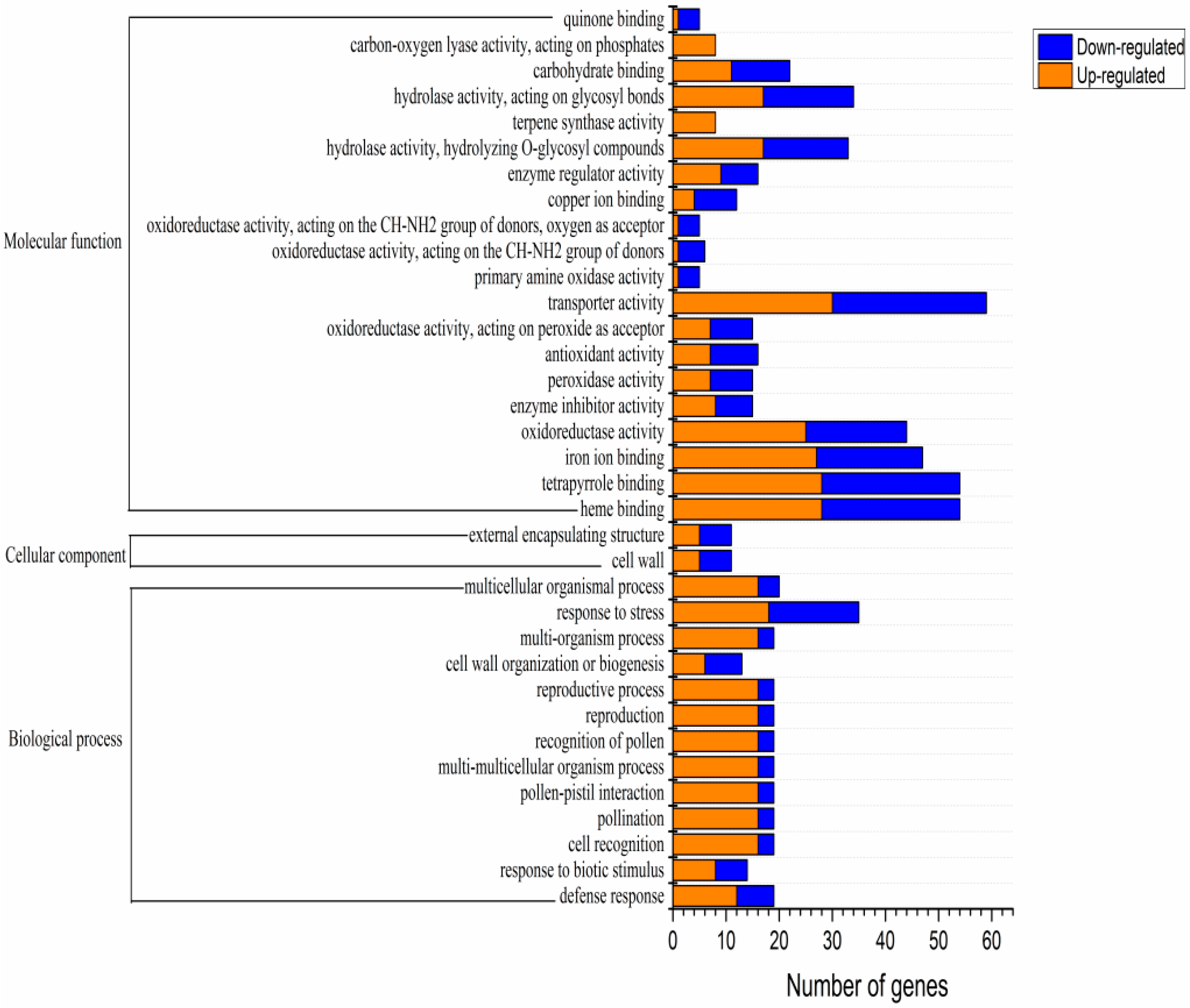
3.2. Analysis of Differentially Expressed Genes and Gene Enrichment in CKY versus CKN Libraries
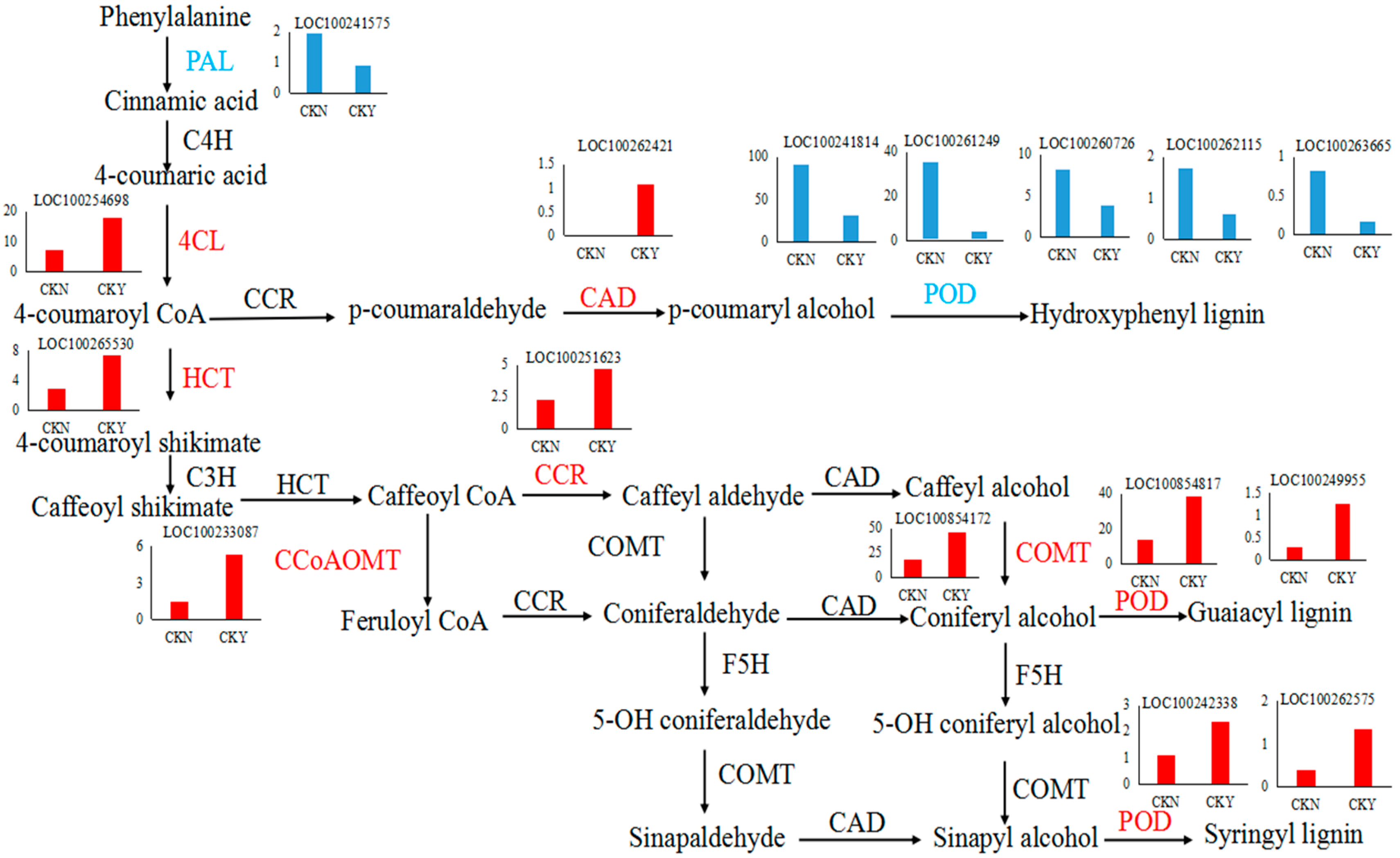
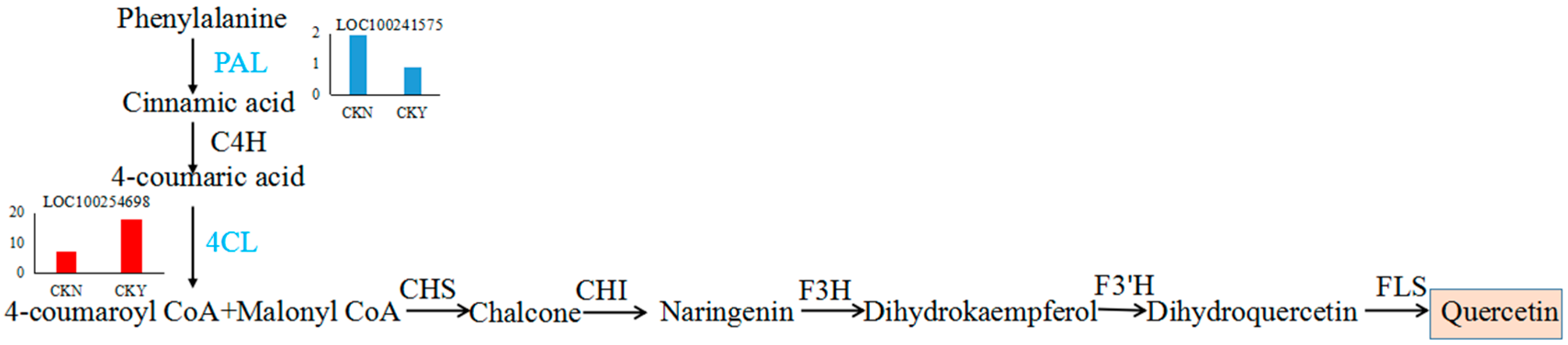
3.3. Differentially Expressed Genes Involved in Phenylpropane Synthesis
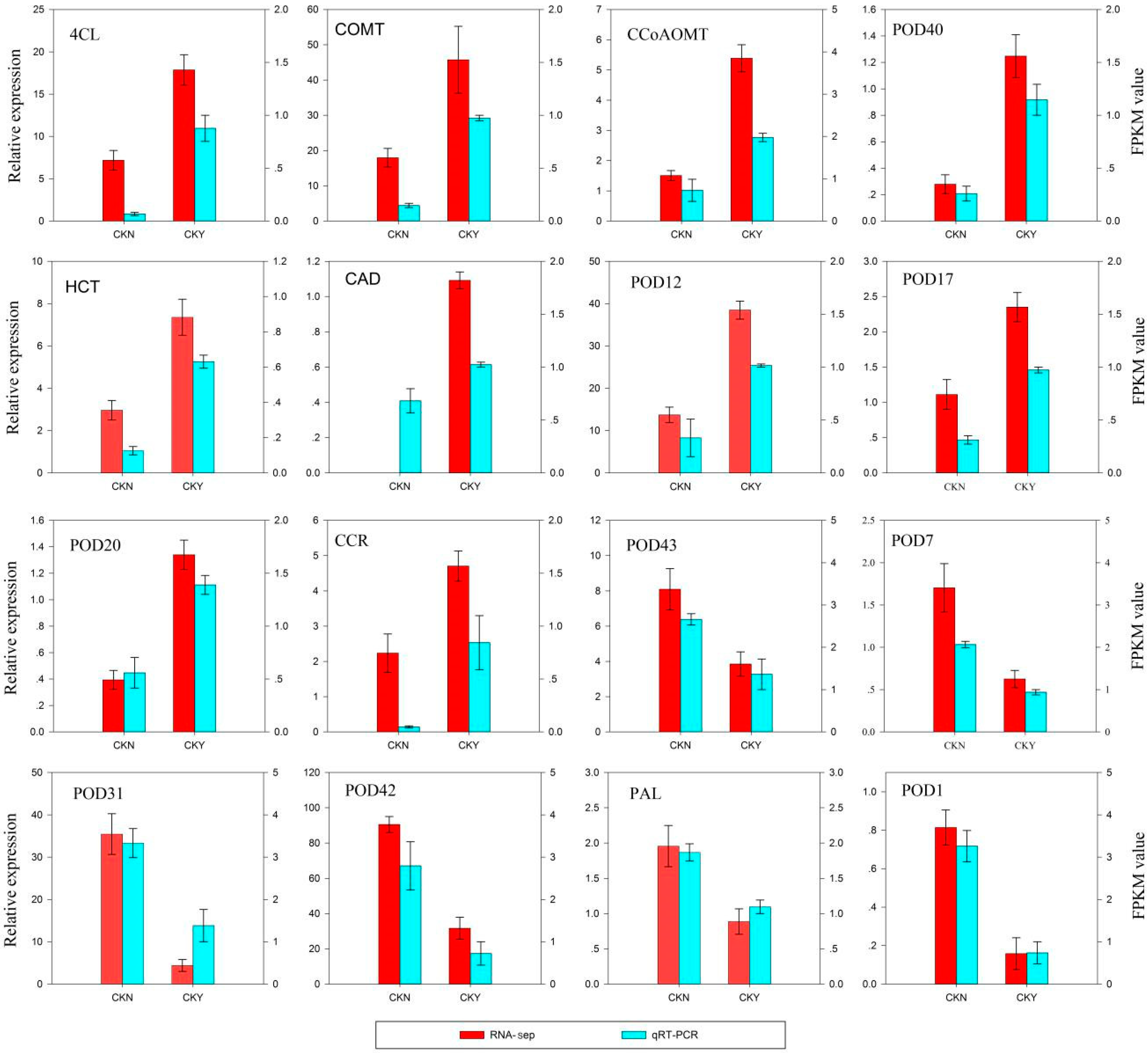
3.4. Reliability Validation of RNA-seq by qRT-PCR
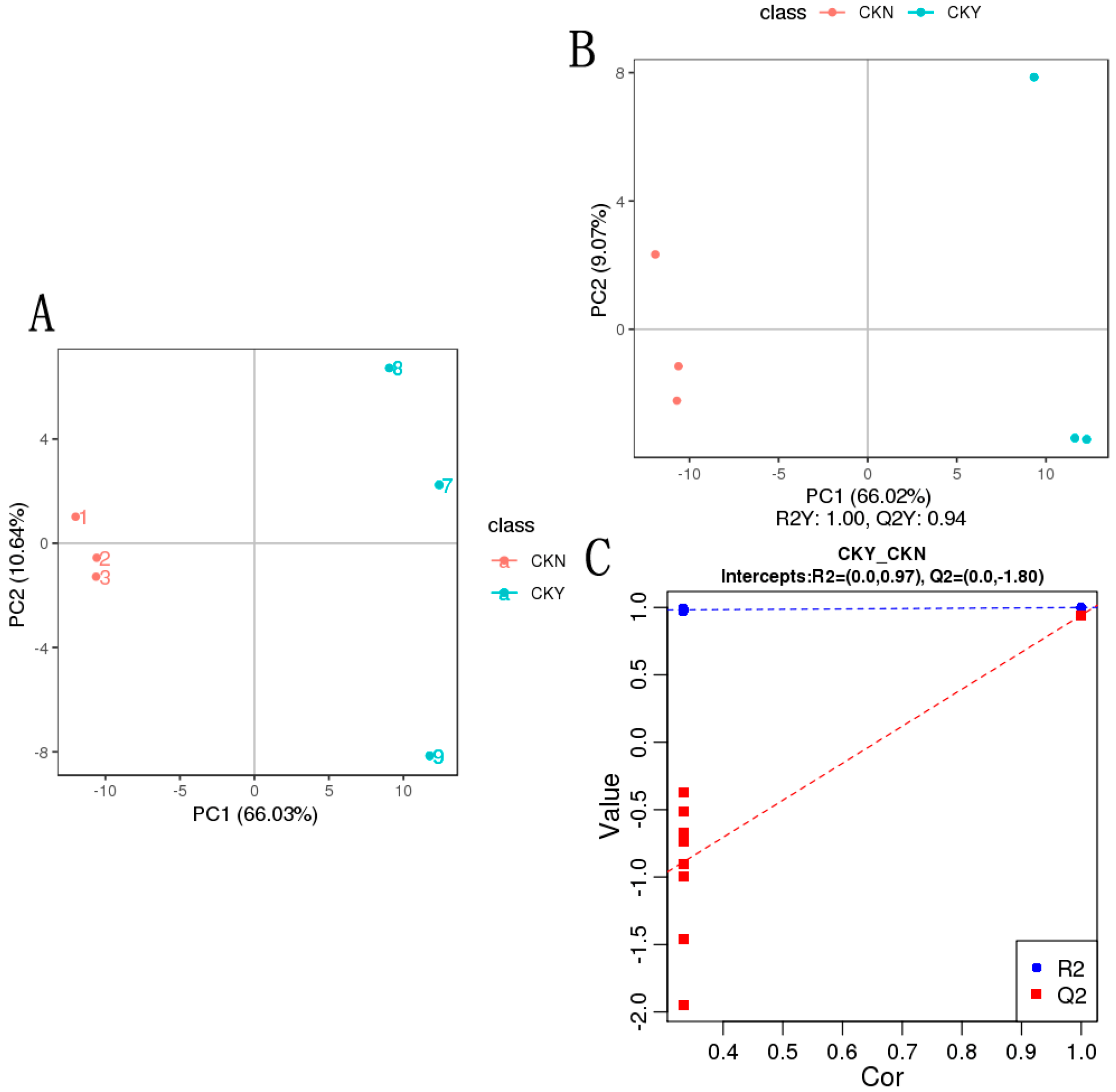
3.5. Metabolite PCA and PLS-DA
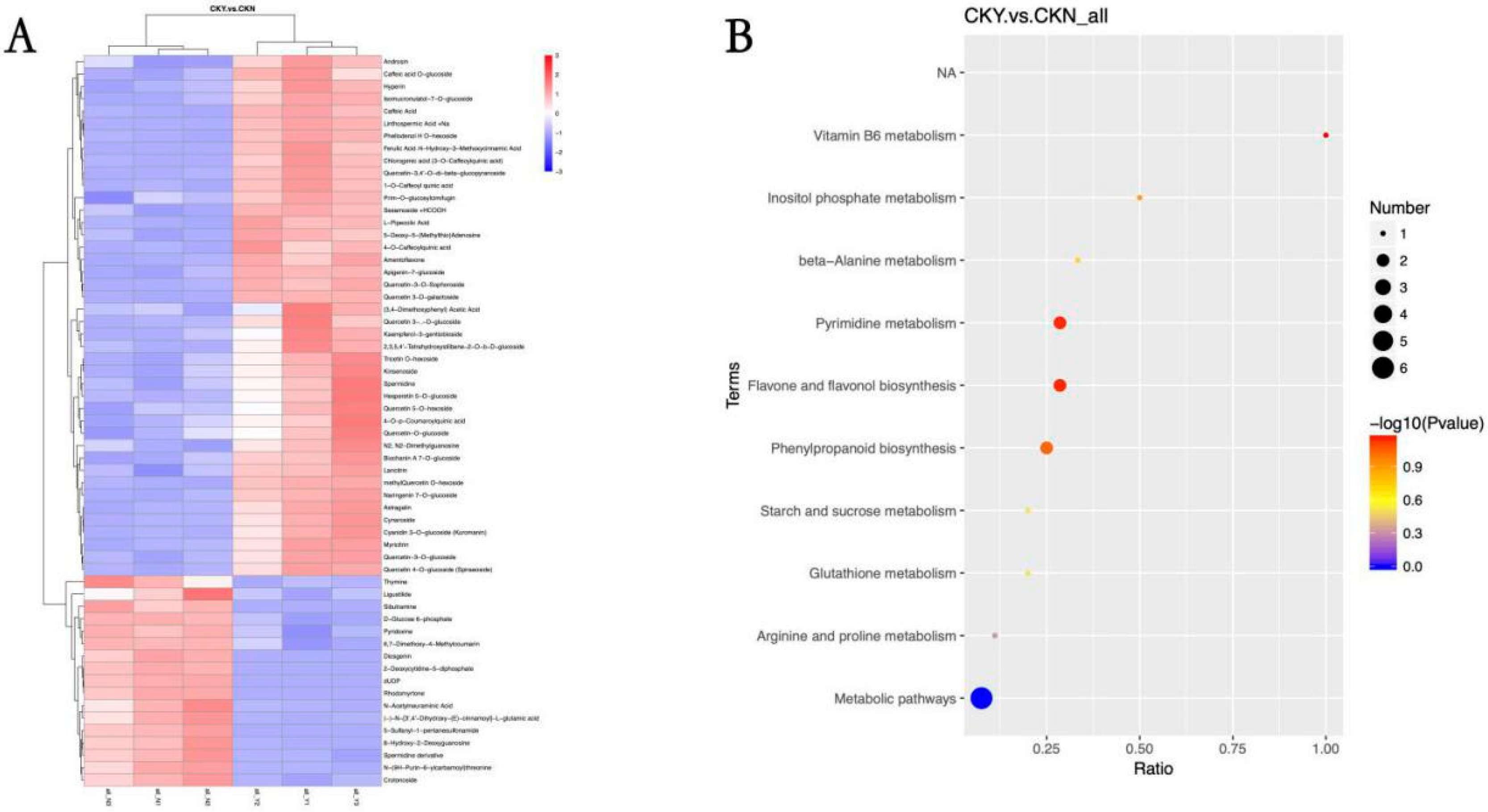
3.6. Differential Metabolite Analysis
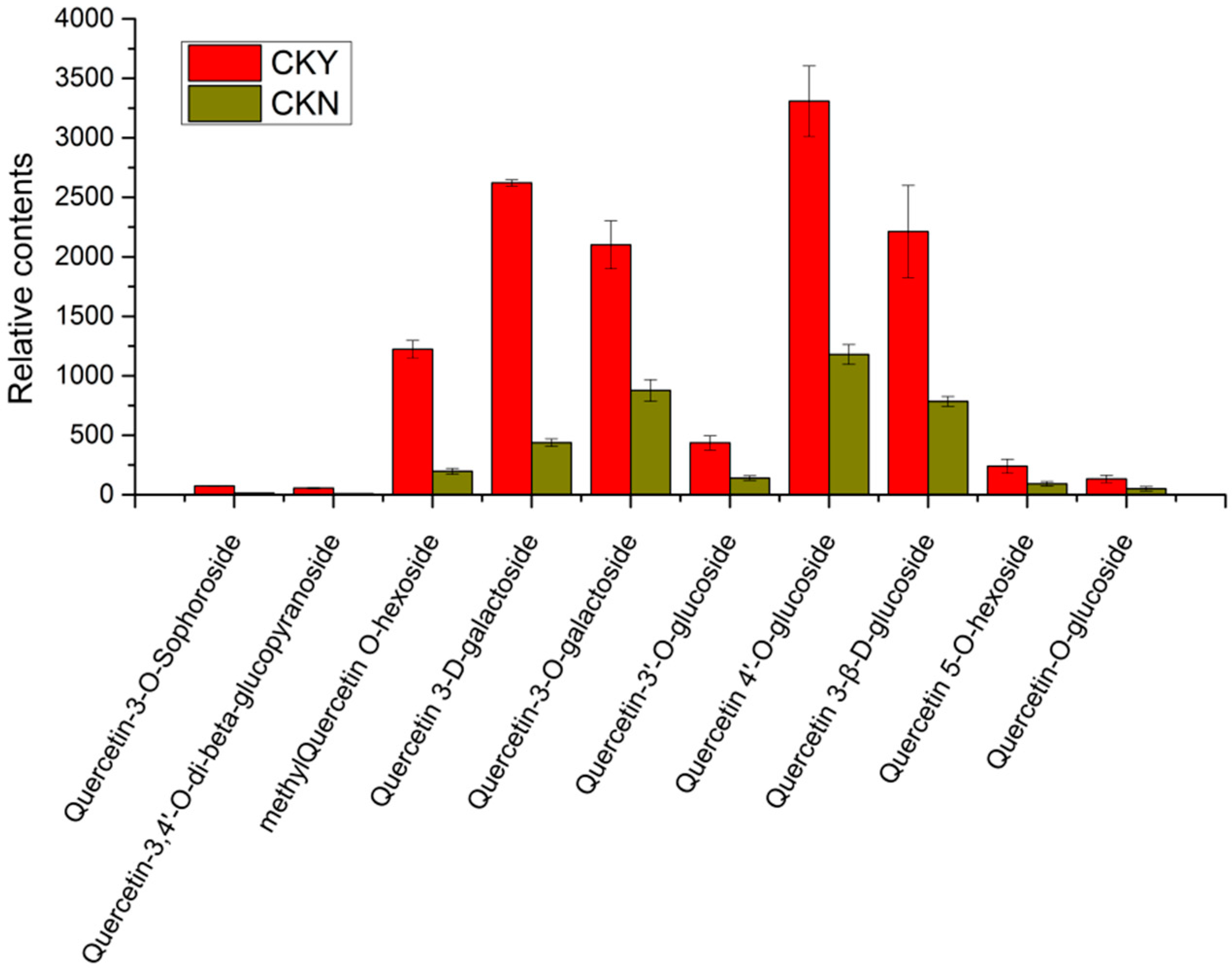
3.7. Analysis of Relative Contents of Key Differential Metabolites
3.8. Analysis of Different Metabolites and Metabolic Pathways
3.9. Association Analysis between Transcriptomic and Metabolomic Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falginella, L.; Cipriani, G.; Monte, C.; Gregori, R.; Tartarini, S. A major QTL controlling apple skin russeting maps on the linkage group 12 of ‘renetta grigia di torriana’. BMC Plant Biol. 2015, 15, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuri, J.A.; Castelli, R. Pear russet control with gibberellins and other products, in cv. Packham’s Triumph. Acta Hortic. 1998, 475, 303–310. [Google Scholar] [CrossRef]
- Liang, W.; Dondini, L.; De Franceschi, P.; Paris, R.; Sansavini, S.; Tartarini, S. Genetic diversity, population structure and construction of a core collection of apple cultivars from Italian germplasm. Plant Mol. Biol. Rep. 2015, 33, 458–473. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Dai, M.S.; Zhang, S.J.; Shi, Z.B. Exploring candidate genes for pericarp russet pigmentation of sand pear (Pyrus pyrifolia) via RNA-seq data in two genotypes contrasting for pericarp color. PLoS ONE 2014, 9, e83675. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.H.; Qi, B.X.; Wang, X.Q.; Shen, L.Y.; Luo, J.; Zhang, Y.X. Proteomic analysis of the key mechanism of exocarp russet pigmentation of semi-russet pear under rainwater condition. Sci. Hortic. 2019, 254, 178–186. [Google Scholar] [CrossRef]
- Legay, S.; Guerriero, G.; Deleruelle, A.; Lateur, M.; Evers, D.; André, C.M.; Hausman, J.F. Apple russeting as seen through the RNA-seq lens: Strong alterations in the exocarp cell wall. Plant Mol. Biol. 2015, 88, 21–40. [Google Scholar] [CrossRef]
- Lashbrooke, J.; Aharoni, A.; Costa, F. Genome investigation suggests MdSHN3, an APETALA2-domain transcription factor gene, to be a positive regulator of apple fruit cuticle formation and an inhibitor of russet development. J. Exp. Bot. 2015, 66, 6579–6589. [Google Scholar] [CrossRef] [Green Version]
- Heng, W.; Liu, L.; Wang, M.D.; Jia, B.; Liu, P.; Ye, Z.F.; Zhu, L.W. Differentially expressed genes related to the formation of russet fruit skin in a mutant of ‘Dangshansuli’pear (Pyrus bretchnederi Rehd.) determined by suppression subtractive hybridization. Euphytica 2014, 196, 285–297. [Google Scholar] [CrossRef]
- Suehiro, Y.; Mochida, K.; Itamura, H.; Esumi, T. Skin Browning and Expression of PPO, STS, and CHS Genes in the Grape Berries of ‘Shine Muscat’. J. Jpn. Soc. Hortic. Sci. 2014, 83, 122–132. [Google Scholar] [CrossRef]
- Katayama-Ikegami, A.; Suehiro, Y.; Katayama, T.; Jindo, K.; Itamura, H.; Esumi, T. Recombinant expression, purification, and characterization of polyphenol oxidase 2 (Vvppo2) from “shine muscat” (Vitis labruscana Bailey × Vitis vinifera L.). Biosci. Biotechnol. Biochem. 2017, 81, 2330–2338. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, Biological functions and Biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, L.L.; Liu, Z.G.; Zhao, Z.H.; Zhao, J.; Wang, Z.T.; Zhou, G.F.; Liu, P.; Liu, M.J. Transcriptome and metabolome profiling unveil the mechanisms of Ziziphus jujuba Mill. peel coloration. Food Chem. 2020, 312, 125903. [Google Scholar] [CrossRef]
- Ning, T.; Nan, C.; Nan, H.; Wei, D.; Zexiong, C.; Zhengguo, L. Comparative metabolomics and transcriptomic profiling reveal the mechanism of fruit quality deterioration and the resistance of citrus fruit against, Penicillium digitatum. Postharvest Biol. Technol. 2018, 145, 61–73. [Google Scholar]
- Suzuki, M.; Nakabayashi, R.; Ogata, Y.; Sakurai, N.; Tokimatsu, T.; Goto, S.; Jasinski, M.; Martinoia, E.; Otagaki, S.; Matsumoto, S.; et al. Multiomics in grape berry skin revealed specific induction of the stilbene synthetic pathway by ultraviolet-C irradiation. Plant Physiol. 2015, 168, 47–59. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical methods for identifying differentially expressed genes in DNA microarrays. Methods Mol. Biol. 2003, 224, 149–157. [Google Scholar]
- De Vos, R.C.; Moco, S.; Lommen, A.; Keurentjes, J.J.; Bino, R.J.; Hall, R.D. Untargeted large-scale plant metabolomics using liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2007, 2, 778–791. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Want, E.J.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC–MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar]
- Boulesteix, A.L.; Strimmer, K. Partial least squares: A versatile tool for the analysis of high-dimensional genomic data. Brief. Bioinform. 2007, 8, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Ren. Physiol. 2016, 310, F857–F871. [Google Scholar] [CrossRef] [Green Version]
- Heischmann, S.; Quinn, K.; Cruickshank-Quinn, C.; Liang, L.P.; Reisdorph, R.; Reisdorph, N.; Patel, M. Exploratory metabolomics profiling in the kainic acid rat model reveals depletion of 25-hydroxyvitamin d3 during epileptogenesis. Sci. Rep. 2016, 6, 31424. [Google Scholar] [CrossRef] [Green Version]
- Haspel, J.A.; Chettimada, S.; Shaik, R.S.; Chu, J.H.; Raby, B.A.; Cernadas, M.; Carey, V.; Process, V.; Hunninghake, G.M.; Ifedigbo, E.; et al. Circadian rhythm reprogramming during lung inflammation. Nat. Commun. 2014, 5, 4753. [Google Scholar] [CrossRef]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Chinnaiyan, A.M. Corrigendum: Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 2009, 457, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Heng, W.; Wang, Z.T.; Jiang, X.H.; Jia, B.; Liu, P.; Liu, L.; Ye, Z.F.; Zhu, L.W. The role of polyamines during exocarp formation in a russet mutant of ‘Dangshansuli’ pear (Pyrus bretschneideri Rehd.). Plant Cell Rep. 2016, 35, 1841–1852. [Google Scholar] [CrossRef]
- Frezza, C.; De Vita, D.; Spinaci, G.; Sarandrea, M.; Venditti, A.; Bianco, A. Secondary metabolites of Tilia tomentosa Moench inflorescences collected in Central Italy: Chemotaxonomy relevance and phytochemical rationale of traditional use. Nat. Prod. Res. 2018, 1–8. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhang, S.; Dai, M.S.; Shi, Z.B. Pigmentation in sand pear (Pyrus pyrifolia) fruit: Biochemical characterization, gene discovery and expression analysis with exocarp pigmentation mutant. Plant Mol. Biol. 2014, 85, 123–134. [Google Scholar] [CrossRef]
- Heng, W.; Wang, M.D.; Yang, J.Y.; Wang, Z.T.; Jiang, X.H.; Zhu, L.W. Relationship between H2O2 in polyamine metabolism and lignin in the exocarp of a russet mutant of ‘dangshansuli’ pear (Pyrus bretschneideri Rehd.). Plant Mol. Biol. Report. 2016, 34, 1056–1063. [Google Scholar] [CrossRef]
- Xie, C.; Veitch, N.C.; Houghton, P.J.; Simmonds, M.S.J. Flavonoid glycosides and isoquinolinone alkaloids from Corydalis bungeana. Phytochemistry 2004, 65, 3041–3047. [Google Scholar] [CrossRef]
- Gutiérrez-Hermosín, I.; Sánchez-Palomo, E.; Vicario-Espinosa, A. Phenolic composition and magnitude of copigmentation in young and shortly aged red wines made from the cultivars, Cabernet Sauvignon, Cencibel, and Syrah. Food Chem. 2005, 92, 269–283. [Google Scholar] [CrossRef]
- Lee, J.; Mitchell, A.E. Quercetin and Isorhamnetin Glycosides in Onion (Allium cepa L.): Varietal Comparison, Physical Distribution, Coproduct Evaluation, and Long-Term Storage Stability. J. Agric. Food Chem. 2011, 59, 857–863. [Google Scholar] [CrossRef]
- Flamini, R.; Mattivi, F.; Rosso, M.; Arapitsas, P.; Bavaresco, L. Advanced knowledge of three important classes of grape phenolics: Anthocyanins, stilbenes and flavonols. Int. J. Mol. Sci. 2013, 14, 19651–19669. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Griesser, M.; Hoffmann, T.; Bellido, M.L.; Rosati, C.; Fink, B.; Kurtzer, R.; Aharoni, A.; Muñoz-Blanco, J.; Schwab, W. Redirection of flavonoid biosynthesis through the down-regulation of an anthocyanidin glucosyltransferase in ripening strawberry fruit. Plant Physiol. 2008, 146, 1528–1539. [Google Scholar] [CrossRef] [Green Version]
- Devaiah, S.P.; Tolliver, B.M.; Zhang, C.; Owens, D.K.; Mcintosh, C.A. Mutational analysis of substrate specificity in a Citrus paradisi flavonol 3-O-glucosyltransferase. J. Plant Biochem. Biotechnol. 2018, 27, 13–27. [Google Scholar] [CrossRef]
- Ono, E.; Homma, Y.; Horikawa, M.; Kunikane-Doi, S.; Imai, H.; Takahashi, S.; Kawai, Y.; Ishiguro, M.; Fukui, Y.; Nakayama, T. Functional differentiation of the glycosyltransferases that contribute to the chemical diversity of bioactive flavonol glycosides in grapevines(Vitis vinifera). Plant Cell 2010, 22, 2856–2871. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Gao, J.J.; Feng, X.X.; Shi, Z.X.; Gao, F.Y.; Xu, X.L. Comparison of peel characteristics and antirusset technology of golden crown ′ apple and its non-russet buds. Hortic. J. 2014, 41, 35–43. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3′) | Accession Number of Reference Genes Deposited in NCBI |
|---|---|---|
| 4CL | F:GTGTTGGCGATTGCGAAGA R:AGCTTGGCTCTGACAGTGT | XM_002272746.4 |
| COMT | F:CCTGGTGTGGAGAATGTTGG R:TTGTCTGGAAGTGCCTGATAAC | XM_003634113.2 |
| CAD | F:CGACGGCAAGTTGATTCTCTT R:AGCACTTCCTCTGTCTCCTTC | XM_002285358.4 |
| HCT | F:CGCCAGCAAGATCCACAAC R:CGCACCAGAGCCGTTAGAT | XM_002268952.3 |
| CCo AOMT | F:AATCATCGGCTACGACAACAC R:GCTCCAACACGAAGTCTCTG | NM_001281118.1 |
| POD40 | F:CGACATTCACCTCAAGGCTAAC R:AGGCGTCACAAGGTCAAGTT | XM_002273323.3 |
| POD17 | F:GGGTGTGATGCTTCTTTGTTAC R:TGACTTCTCCAATGCTTCCTTC | XM_002271047.3 |
| POD12 | F:GCTTGCTTCGCCTCCACTT R:TTCTTGTTCACCAGGACCACTT | XM_003634474.3 |
| POD20 | F:GGATGCGATGCCTCTATTCTTC R:TGCCTCTTCTACCAAGTGCTT | XM_002279172.4 |
| CCR | F:CCACACTGCTTCTCCTGTCA R:GCCGCTGCTATTATCACATTCT | XM_002273418.3 |
| POD42 | F:GCCAAGAGCCAAGACTACTTC R:GTGCCAGTGAGAGGATTGTTC | XM_002274733.3 |
| POD31 | F:GCAGGATACCATCACCAACAAG R:AGGAGACAAGAACGGAAGCAT | XM_002280511.3 |
| POD43 | F:GCTGATATGCCTGATGTGAGTG R:GCTGTGGTTCCAATGGTGTG | XM_002270914.4 |
| POD7 | F:AAGCAGAGGTTGAGAAGAGGT R:AATGAGGACGGTGGCATCT | XM_002265631.3 |
| POD1 | F:GTGAACACGGCAGTGAACAA R:TGTCGTCCAGCAGGATTGAT | XM_010655057.2 |
| PAL | F:CCAAGGATACTCAGGCATCAGA R:GAGGCAAGCAAGGACTAATGTT | XM_002285241.3 |
| VvGAPDH | F:TTCTCGTTGAGGGCTATTCCA R:CCACAGACTTCATCGGTGACA | GU585870 |
| Sample | Total Raw Reads | Total Clean Reads | Clean Bases | GC Percentage | Q20 Percentage | Q30 Percentage |
|---|---|---|---|---|---|---|
| CKY CKN | 87,831,088 84,654,916 | 85,425,282 82,194,414 | 6.41 G 6.17 G | 46.26% 46.87% | 97.58% 97.65% | 93.03% 93.27% |
| KEGG ID | Metabolic Pathway | Number of DEGs | p-Value | Up-Regulated Gene Counts | Down-Regulated Gene Counts |
|---|---|---|---|---|---|
| vvi00940 | Phenylpropanoid biosynthesis | 31(11.40%) | 7.22 × 10−12 | 20 | 11 |
| vvi00904 | Diterpenoid biosynthesis | 7(2.57%) | 3.38 × 10−06 | 4 | 3 |
| vvi00950 | Isoquinoline alkaloid biosynthesis | 8(2.94%) | 9.05 × 10−06 | 4 | 4 |
| vvi00350 | Tyrosine metabolism | 11(4.04%) | 1.82 × 10−05 | 5 | 6 |
| vvi00196 | Photosynthesis - antenna proteins | 6(2.21%) | 3.92 × 10−04 | 1 | 5 |
| vvi00960 | Tropane, piperidine, and pyridine alkaloid biosynthesis | 7(2.57%) | 7.32 × 10−04 | 2 | 5 |
| vvi00480 | Glutathione metabolism | 14(5.15%) | 2.65 × 10−03 | 8 | 6 |
| vvi04075 | Plant hormone signal transduction | 23(8.46%) | 4.24 × 10−03 | 19 | 4 |
| Gene ID | log2(FPKMCKY/FPKMCKN) | p-Value | Gene Description | Up- or Down-Regulated |
|---|---|---|---|---|
| 100267863 | 2.31 | 1.60 × 10−16 | cytochrome P450 84A1 | up |
| 100854817 | 1.49 | 5.41 × 10−15 | peroxidase 12 | up |
| 100854172 | 1.35 | 2.57 × 10−12 | caffeic acid 3-O-methyltransferase | up |
| 100253961 | 1.64 | 1.53 × 10−08 | berberine bridge enzyme-like 26 | up |
| 100250740 | 1.01 | 1.46 × 10−07 | berberine bridge enzyme-like 15 | up |
| 100262421 | 6.95 | 1.64 × 10−07 | probable cinnamyl alcohol dehydrogenase 1 | up |
| 100265530 | 1.31 | 4.31 × 10−07 | shikimate O-hydroxycinnamoyltransferase | up |
| 100233087 | 1.84 | 6.10 × 10−06 | caffeoyl-CoA O-methyltransferase | up |
| 100261642 | 3.98 | 1.03 × 10−04 | probable mannitol dehydrogenase | up |
| 100249955 | 2.15 | 1.18 × 10−04 | peroxidase 40 | up |
| 100265092 | 2.77 | 2.36 × 10−04 | anthocyanidin 3-O-glucosyltransferase 5 | up |
| 100260786 | 1.89 | 3.24 × 10−04 | quercetin 3-O-methyltransferase 1-like | up |
| 100242338 | 1.08 | 8.54 × 10−04 | peroxidase 17 | up |
| 100254698 | 1.31 | 9.87 × 10−04 | 4-coumarate--CoA ligase | up |
| 100262575 | 1.75 | 1.05 × 10−03 | peroxidase 20 | up |
| 100250160 | 2.21 | 1.24 × 10−03 | beta-glucosidase 12 | up |
| 100251623 | 1.07 | 1.64 × 10−03 | cinnamoyl-CoA reductase 1 | up |
| 100855376 | −1.74 | 1.15 × 10−32 | probable mannitol dehydrogenase | down |
| 100241814 | −1.51 | 1.47 × 10−17 | peroxidase 42 | down |
| 100261249 | −3.01 | 1.83 × 10−14 | peroxidase 31 | down |
| 100854583 | −1.19 | 5.62 × 10−13 | probable mannitol dehydrogenase | down |
| 100247559 | −2.10 | 1.56 × 10−07 | probable mannitol dehydrogenase | down |
| 100252642 | −1.70 | 7.76 × 10−07 | probable mannitol dehydrogenase | down |
| 100260726 | −1.07 | 3.99 × 10−06 | peroxidase 43 | down |
| 100854646 | −1.98 | 1.15 × 10−05 | probable mannitol dehydrogenase | down |
| 100262115 | −1.45 | 6.79 × 10−05 | peroxidase 7 | down |
| 100241575 | −1.14 | 2.52 × 10−04 | phenylalanine ammonia-lyase-like | down |
| 100263665 | −2.35 | 1.17 × 10−03 | cationic peroxidase 1 | down |
| Compared Samples | Num. of Total Ident. | Num. of Total Sig. | Num. of Total Sig. Up | Num. of Total Sig. Down |
|---|---|---|---|---|
| CKY versus CKN | 443 | 60 | 43 | 17 |
| Number | Metabolites | VIP | FC | p-Value | Up- or Down-Regulated |
|---|---|---|---|---|---|
| Phenols | |||||
| 1 | Caffeic Acid | 1.76 | 3.63 | 0.00003 | up |
| 2 | 1-O-Caffeoyl quinic acid | 2.07 | 4.56 | 0.00008 | up |
| 3 | Isomucronulatol-7-O-glucoside | 1.23 | 2.45 | 0.00112 | up |
| 4 | 4-O-Caffeoylquinic acid | 1.57 | 3.16 | 0.00116 | up |
| 5 | Chlorogenic acid (3-O-Caffeoylquinic acid) | 2.02 | 4.43 | 0.00162 | up |
| 6 | Naringenin 7-O-glucoside | 2.87 | 8.02 | 0.0021 | up |
| 7 | Astragalin | 1.43 | 2.87 | 0.00316 | up |
| 8 | Myricitrin | 1.27 | 2.55 | 0.004 | up |
| 9 | Cynaroside | 1.56 | 3.16 | 0.00522 | up |
| 10 | Amentoflavone | 3.35 | 11.00 | 0.00631 | up |
| 11 | Cyanidin 3-O-glucoside (Kuromanin) | 1.51 | 3.05 | 0.00815 | up |
| 12 | ε-Viniferin 2 | 2.30 | 5.23 | 0.01003 | up |
| 13 | Kaempferol-3-gentiobioside | 3.12 | 8.91 | 0.01074 | up |
| 14 | Hesperetin 5-O-glucoside | 1.50 | 3.07 | 0.01526 | up |
| 15 | Biochanin A 7-O-glucoside | 2.63 | 6.13 | 0.02243 | up |
| 16 | Laricitrin | 1.06 | 2.14 | 0.02356 | up |
| 17 | 4-O-p-Coumaroylquinic acid | 1.31 | 2.61 | 0.01148 | up |
| 18 | Ferulic acid | 2.44 | 6.03 | 0.00234 | up |
| 19 | Caffeic acid O-glucoside | 1.49 | 2.97 | 0.00134 | up |
| 20 | Quercetin-3,4′-O-di-beta-glucopyranoside | 2.59 | 6.62 | 0.00008 | up |
| 21 | Quercetin-3-O-Sophoroside | 2.39 | 5.74 | 0.00001 | up |
| 22 | Quercetin-O-glucoside | 1.36 | 2.60 | 0.04101 | up |
| 23 | Quercetin 5-O-hexoside | 1.29 | 2.60 | 0.0182 | up |
| 24 | Quercetin 3-β-d-glucoside | 1.40 | 2.82 | 0.00861 | up |
| 25 | Quercetin 4′-O-glucoside (Spiraeoside) | 1.29 | 2.59 | 0.00171 | up |
| 26 | Quercetin-3′-O-glucoside | 1.54 | 3.10 | 0.00147 | up |
| 27 | Quercetin-3-O-galactoside | 1.19 | 2.40 | 0.00103 | up |
| 28 | Methylquercetin O-hexoside | 2.50 | 6.20 | 0.00041 | up |
| 29 | Quercetin 3-d-galactoside | 2.45 | 5.98 | 0.00068 | up |
| Nucleic acids and their derivatives | |||||
| 30 | dUDP(2′-deoxyuridine 5′-diphosphate) | 5.65 | 0.02 | 0 | down |
| 31 | 2′-Deoxycytidine-5′-diphosphate | 5.55 | 0.02 | 0.00004 | down |
| 32 | 8-Hydroxy-2-Deoxyguanosine | 2.41 | 0.17 | 0.00018 | down |
| 33 | N-(9H-Purin-6-ylcarbamoyl)threonine | 2.30 | 0.18 | 0.00057 | down |
| 34 | 5′-Deoxy-5′-(Methylthio)Adenosine | 1.23 | 2.46 | 0.0016 | up |
| 35 | Crotonoside | 2.43 | 0.17 | 0.00481 | down |
| 36 | N2,N2-Dimethylguanosine | 1.23 | 2.44 | 0.01038 | up |
| 37 | Thymine | 1.06 | 0.46 | 0.01272 | down |
| Amino acids and their derivatives | |||||
| 38 | l-Pipecolic Acid | 1.23 | 2.45 | 0.00008 | up |
| 39 | N-Acetylneuraminic Acid | 1.29 | 0.39 | 0.00897 | down |
| 40 | (-)-N-[3′,4′-Dihydroxy-(E)-cinnamoyl]-l-glutamic acid | 1.33 | 0.37 | 0.01037 | down |
| Others | |||||
| 41 | Diosgenin | 3.82 | 0.06 | 0.00003 | down |
| 42 | Rhodomyrtone | 3.40 | 0.08 | 0.00006 | down |
| 43 | 5-Sulfanyl-1-pentanesulfonamide | 3.22 | 0.09 | 0.0001 | down |
| 44 | 6,7-Dimethoxy-4-Methylcoumarin | 1.16 | 0.44 | 0.02876 | down |
| 45 | Spermidine derivative | 2.31 | 0.19 | 0.00193 | down |
| 46 | d-Glucose 6-phosphate | 1.19 | 0.42 | 0.00937 | down |
| 47 | 2,3,5,4′-Tetrahydroxystilbene-2-O-β-d-glucoside | 1.37 | 2.77 | 0.01094 | up |
| 48 | Sibutramine | 5.84 | 0.02 | 0.01287 | down |
| 49 | Spermidine | 1.25 | 2.50 | 0.01395 | up |
| 50 | Ligustilide | 1.08 | 0.45 | 0.01732 | down |
| 51 | Sesamoside +HCOOH | 1.69 | 3.33 | 0.02052 | up |
| 52 | Pyridoxine | 1.09 | 0.46 | 0.02286 | down |
| 53 | (3,4-Dimethoxyphenyl) acetic acid | 1.30 | 2.64 | 0.03423 | up |
| 54 | Androsin | 1.51 | 2.84 | 0.03835 | up |
| 55 | Kinsenoside | 1.53 | 3.05 | 0.00821 | up |
| 56 | Lithospermic acid +Na | 3.53 | 13.20 | 0.00003 | up |
| 57 | Prim-O-glucosylcimifugin | 1.08 | 2.13 | 0.04191 | up |
| 58 | Apigenin-7-glucoside | 3.21 | 8.93 | 0.02621 | up |
| 59 | Phellodenol H O-hexoside | 1.37 | 2.74 | 0.00008 | up |
| 60 | Tricetin O-hexoside | 1.39 | 2.78 | 0.00623 | up |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Liang, D.; Xia, H.; Lin, L.-J.; Wang, J.; Lv, X.-L. Lignin and Quercetin Synthesis Underlies Berry Russeting in ‘Sunshine Muscat’ Grape. Biomolecules 2020, 10, 690. https://doi.org/10.3390/biom10050690
Huang Y, Liang D, Xia H, Lin L-J, Wang J, Lv X-L. Lignin and Quercetin Synthesis Underlies Berry Russeting in ‘Sunshine Muscat’ Grape. Biomolecules. 2020; 10(5):690. https://doi.org/10.3390/biom10050690
Chicago/Turabian StyleHuang, Yan, Dong Liang, Hui Xia, Li-Jin Lin, Jin Wang, and Xiu-Lan Lv. 2020. "Lignin and Quercetin Synthesis Underlies Berry Russeting in ‘Sunshine Muscat’ Grape" Biomolecules 10, no. 5: 690. https://doi.org/10.3390/biom10050690
APA StyleHuang, Y., Liang, D., Xia, H., Lin, L. -J., Wang, J., & Lv, X. -L. (2020). Lignin and Quercetin Synthesis Underlies Berry Russeting in ‘Sunshine Muscat’ Grape. Biomolecules, 10(5), 690. https://doi.org/10.3390/biom10050690