snoRNPs: Functions in Ribosome Biogenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Structure and Components
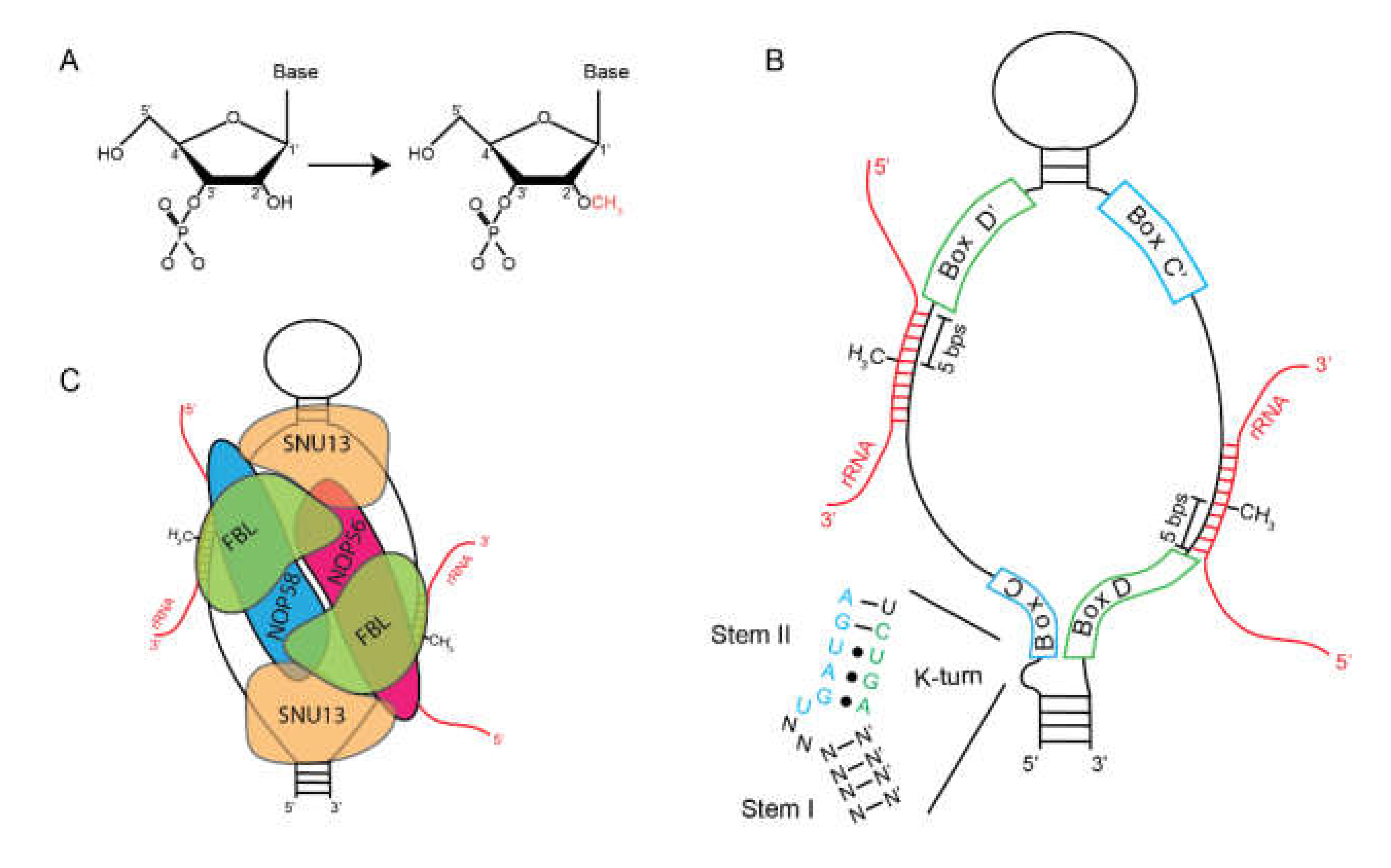
2.1. Box C/D snoRNAs
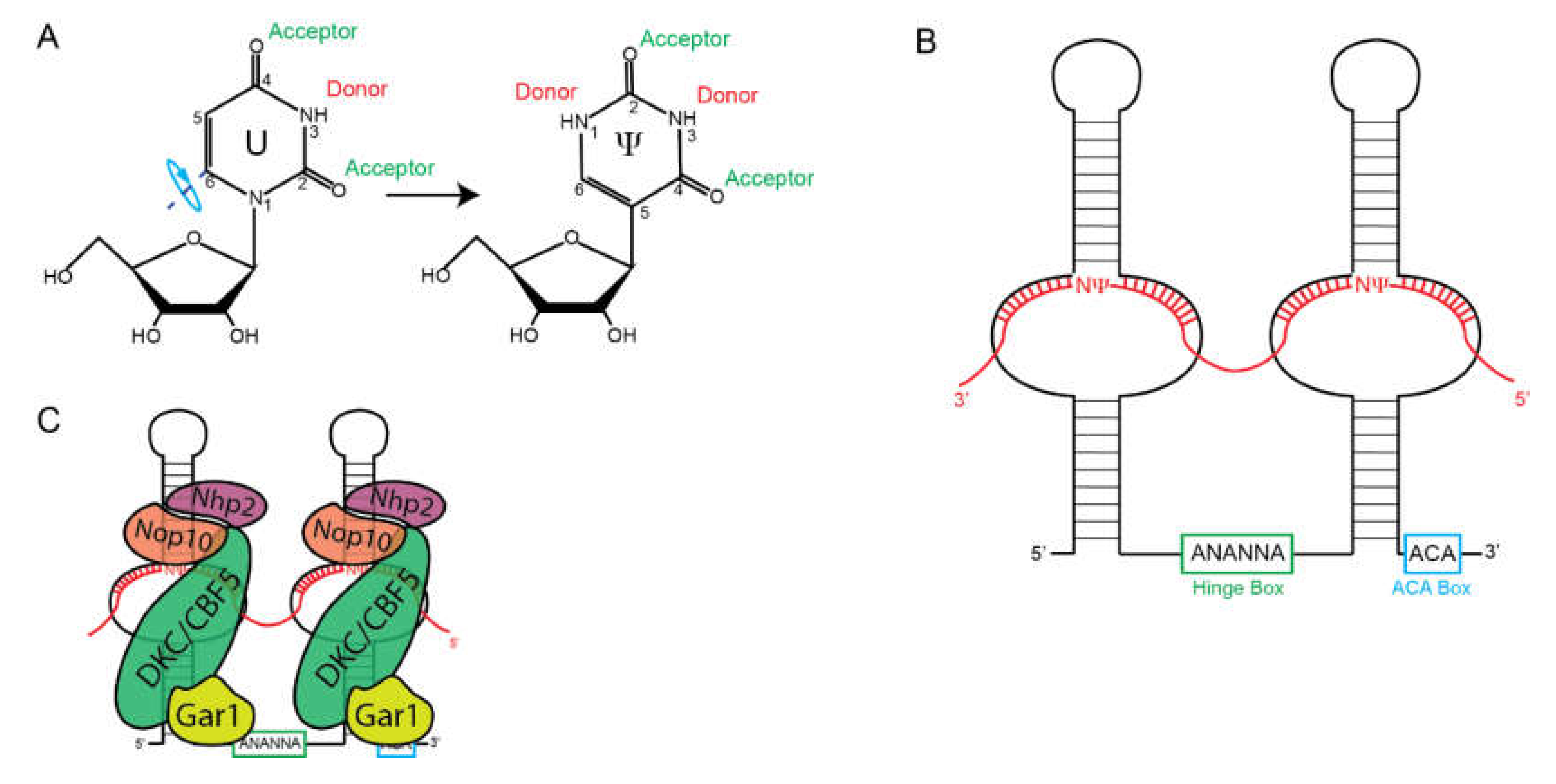
2.2. Box H/ACA snoRNAs
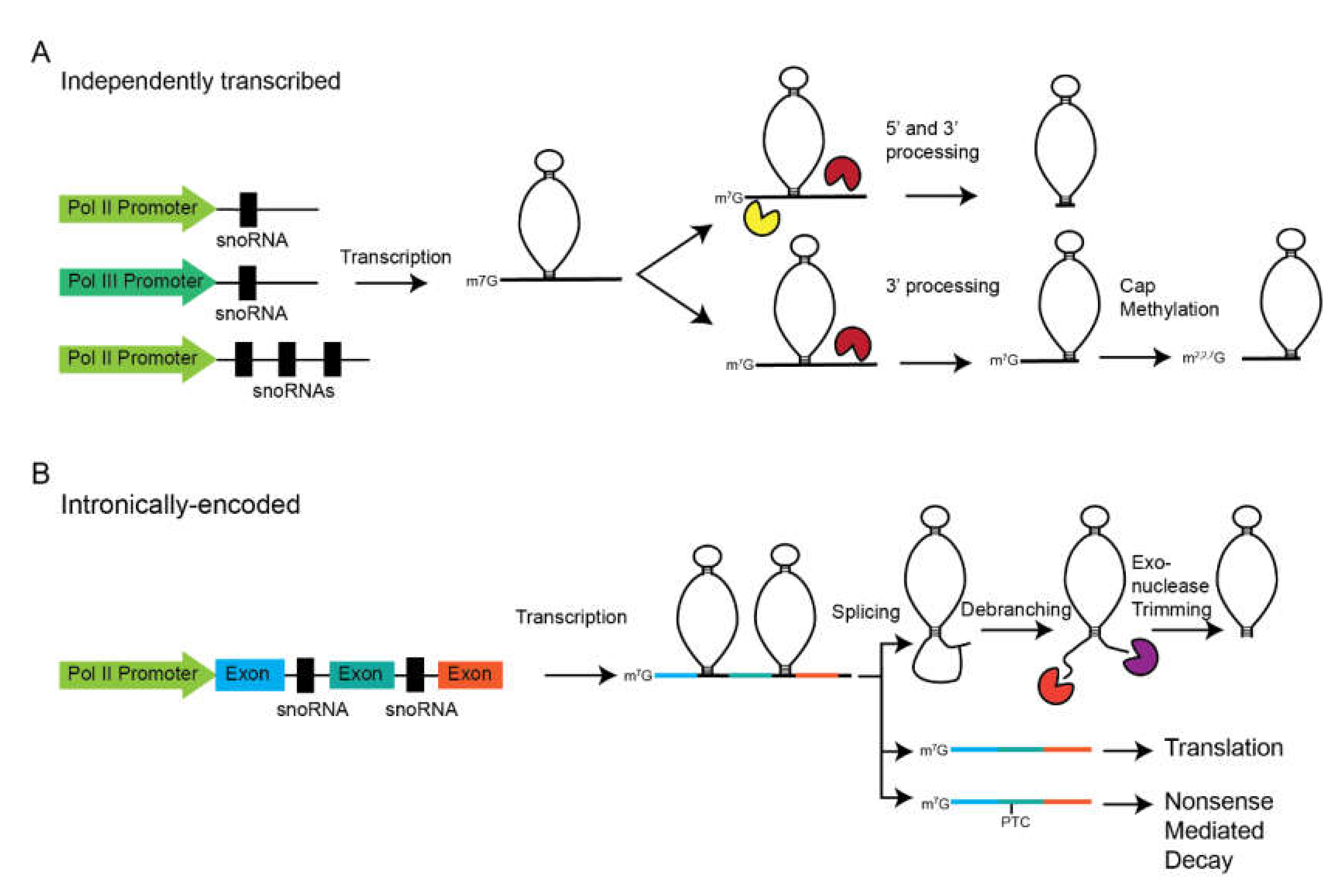
3. Transcription and Processing of snoRNAs
4. Role of Modifications in Ribosome Function
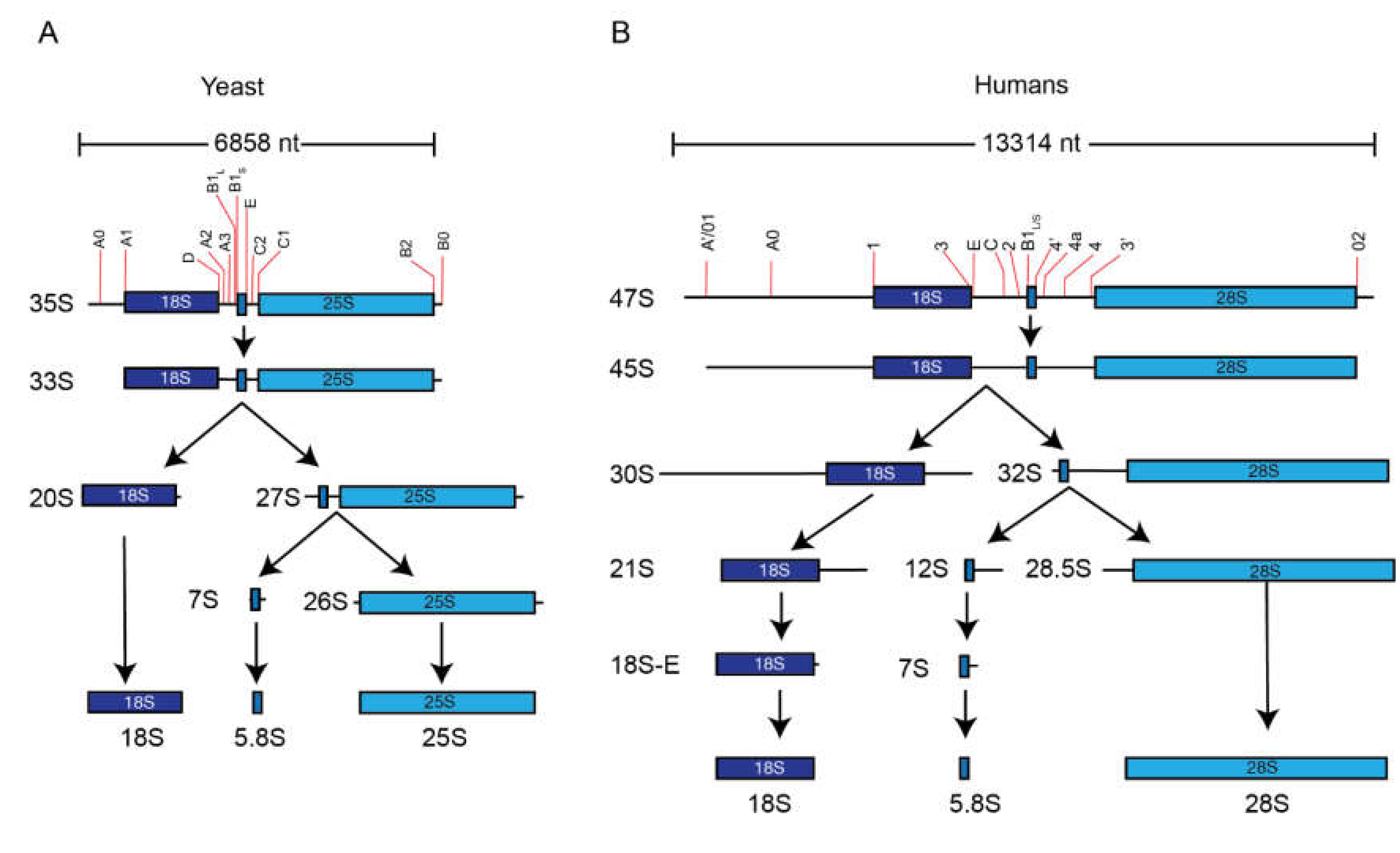
5. Pre-rRNA Processing
6. The snoRNPs’ Role in rRNA Processing
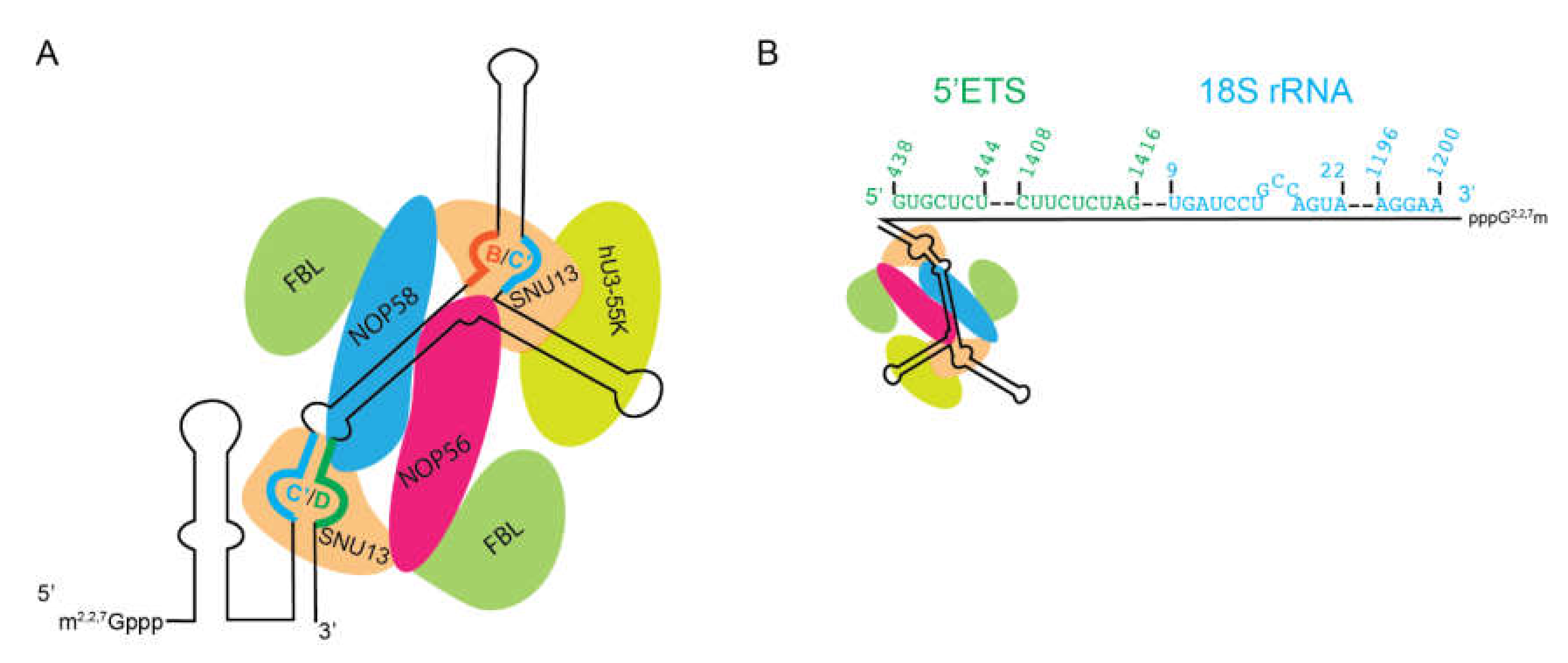
6.1. U3 (SNORD3) snoRNP
6.2. RNA Component of RNase MRP Complex (RMRP)
6.3. Other snoRNAs Necessary for rRNA Processing
7. Role of snoRNP-Mediated Modifications in rRNA Processing
8. Diseases
9. Future Considerations
Abbreviations/Nomenclature
| RNA | ribonucleic acid |
| bp | base pair |
| nt | nucleotide |
| rRNA | ribosomal RNA |
| mRNA | messenger RNA |
| snoRNP | small nucleolar ribonucleoprotein |
| snoRNA | small nucleolar RNA |
| snRNA | small nuclear RNA |
| G | guanosine |
| C | cytosine |
| A | adenosine |
| T | thymidine |
| U | uridine |
| Ψ | pseudouridine |
| 2′Ome | 2′-O-methyl |
| RMRP | RNA component of mitochondrial RNA processing endonuclease |
| TERC | telomerase, RNA component |
| SNU13 | small nuclear ribonucleoprotein 13 |
| NOP58 | nucleolar protein 58 |
| NOP56 | nucleolar protein 56 |
| FBL | fibrillarin |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| NHP2 | non-histone Protein 2 |
| NOP10 | nucleolar Protein 10 |
| Gar1 | glycine/arginine rich-domain containing protein 1 |
| DKC1 | dyskerin |
| Cbf5 | centromere binding factor 5 |
| TBF1 | TTAGGG repeat-binding factor 1 |
| Rap1 | repressor/activator site binding protein 1 |
| NME1 | nuclear mitochondrial processing endoribonuclease |
| RNase | ribonuclease |
| TGS1 | trimethylguanosine synthetase 1 |
| RPS3 | ribosomal protein small subunit 3 |
| RPL7A | ribosomal protein large subunit 7A |
| RPL13A | ribosomal protein large subunit 13A |
| GAS5 | growth arrest specific 5 |
| mTOR | mammalian target of rapamycin |
| HSP90 | heat shock protein 90 |
| R2TP | Rvb1, Rvb2, Tah1, Pih1 complex |
| Rvb1 | RuvB-like protein 1 |
| Rvb2 | RuvB-like protein 2 |
| Tah1 | TPR repeat-containing protein associated with Hsp90 |
| Pih1 | protein interacting with Hsp90 1 |
| RUVBL1 | RuvB-like AAA ATPase 1 |
| RUVBL2 | RuvB-like AAA ATPase 2 |
| RPAP3 | RNA polymerase II associated protein 3 |
| RIH1D1 | PIH1 domain containing 1 |
| snR10 | snoRNA 10 |
| snR30 | snoRNA 30 |
| Sbp1 | suppressor of PaB mutant 1 |
| SSU | small subunit |
| DNase | deoxyribonuclease |
| hU3-55K | human U3 component, 55 kDa |
| Rrp9 | ribosomal RNA processing enzyme 9 |
| cDNA | complementary DNA |
| CRAC | crosslinking and high-throughput analysis of cDNAs |
| Sof1 | suppressor of fibrillarin 1 |
| Mpp10 | M-phase phosphoprotein 10 |
| MDa | megadalton |
| rDNA | ribosomal DNA |
| U3 | uridine-rich RNA 3 |
| ETS | external transcribed spacer |
| ITS | internal transcribed spacer |
| S | Svedberg units |
| MRP | mitochondrial RNA processing enzyme |
| Cas9 | CRISPR-associated protein 9 |
| UTR | untranslated region |
| KRE33/RRA1 | killer toxin resistant 33/ribosomal RNA cytosine acetyltransferase 1 |
| NAT10 | N-Acetyltransferase 10 |
| MEF | mouse embryonic fibroblast |
| KRAS | kirsten rat sarcoma viral oncogene homolog |
| RAS | rat sarcoma |
| EM | electron microscopy |
| tiRNA | tRNA-derived stress-induced RNA |
| tRF | tRNA fragment |
| YB-1 | Y-box protein 1 |
| CMCT | N-Cyclohexyl-N′-(2′morholinoethyl)carbodiimide metho-p-toluenesulfonate |
| HCT116 | human colon cancer 116 |
Funding
Acknowledgments
Conflicts of Interest
References
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piątkowski, P.; Baginski, B.; Wirecki, T.K.; De Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kötter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef] [PubMed]
- Natchiar, S.K.; Myasnikov, A.G.; Kratzat, H.; Hazemann, I.; Klaholz, B. Visualization of chemical modifications in the human 80S ribosome structure. Nature 2017, 551, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Taoka, M.; Nobe, Y.; Yamaki, Y.; Sato, K.; Ishikawa, H.; Izumikawa, K.; Yamauchi, Y.; Hirota, K.; Nakayama, H.; Takahash, N.; et al. Landscape of the complete RNA chemical modifications in the human 80S ribosome. Nucleic Acids Res. 2018, 46, 9289–9298. [Google Scholar] [CrossRef] [PubMed]
- Pletnev, P.; Guseva, E.; Zanina, A.; Evfratov, S.; Dzama, M.; Treshin, V.; Pogorel’Skaya, A.; Osterman, I.; Golovina, A.; Rubtsova, M.; et al. Comprehensive Functional Analysis of Escherichia coli Ribosomal RNA Methyltransferases. Front. Genet. 2020, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Kiss-László, Z.; Henry, Y.; Bachellerie, J.-P.; Caizergues-Ferrer, M.; Kiss, T. Site-specific ribose methylation of preribosomal RNA: A novel function for small nucleolar RNAs. Cell 1996, 85, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, C.; Stebbins-Boaz, B.; Gerbi, S.A. Nucleotide sequence determination and secondary structure of Xenopus U3 snRNA. Nucleic Acids Res. 1988, 16, 2127–2148. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.M.; Konings, D.A.; Cesareni, G. The yeast homologue of U3 snRNA. EMBO J. 1987, 6, 2145–2155. [Google Scholar] [CrossRef]
- Porter, G.L.; Brennwald, P.J.; Holm, K.A.; Wise, J.A. The sequence of U3 from Schizosaccharomyces pombe suggests structural divergence of this snRNA between metazoans and unicellular eukaryotes. Nucleic Acids Res. 1988, 16, 10131–10152. [Google Scholar] [CrossRef] [Green Version]
- Tyc, K. and J.A. Steitz, U3, U8 and U13 comprise a new class of mammalian snRNPs localized in the cell nucleolus. EMBO J. 1989, 8, 3113–3119. [Google Scholar] [CrossRef]
- Klein, D.; Schmeing, T.; Moore, P.; Steitz, T. The kink-turn: A new RNA secondary structure motif. EMBO J. 2001, 20, 4214–4221. [Google Scholar] [CrossRef]
- Watkins, N.J.; Dickmanns, A.; Luhrmann, R. Conserved stem II of the box C/D motif is essential for nucleolar localization and is required, along with the 15.5K protein, for the hierarchical assembly of the box C/D snoRNP. Mol. Cell. Biol. 2002, 22, 8342–8352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollervey, D.; Lehtonen, H.; Jansen, R.; Kern, H.; Hurt, E.C. Temperature-sensitive mutations demonstrate roles for yeast fibrillarin in pre-rRNA processing, pre-rRNA methylation, and ribosome assembly. Cell 1993, 72, 443–457. [Google Scholar] [CrossRef]
- Christensen, M.E.; Beyer, A.L.; Walker, B.; LeStourgeon, W.M. Identification of NG, NG-dimethylarginine in a nuclear protein from the lower eukaryote physarum polycephalum homologous to the major proteins of mammalian 40S ribonucleoprotein particles. Biochem. Biophys. Res. Commun. 1977, 74, 621–629. [Google Scholar] [CrossRef]
- Ochs, R.L.; Lischwe, M.A.; Spohn, W.H.; Busch, H. Fibrillarin: A new protein of the nucleolus identified by autoimmune sera. Biol. Cell 1985, 54, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Tollervey, D.; Hurt, E.C. A U3 snoRNP protein with homology to splicing factor PRP4 and G beta domains is required for ribosomal RNA processing. EMBO J. 1993, 12, 2549–2558. [Google Scholar] [CrossRef] [PubMed]
- Watkins, N.; Ségault, V.; Charpentier, B.; Nottrott, S.; Fabrizio, P.; Bachi, A.; Wilm, M.; Rosbash, M.; Branlant, C.; Lührmann, R. A common core RNP structure shared between the small nucleoar box C/D RNPs and the spliceosomal U4 snRNP. Cell 2000, 103, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Nottrott, S.; Hartmuth, K.; Fabrizio, P.; Urlaub, H.; Vidović, I.; Ficner, R.; Lührmann, R. Functional interaction of a novel 15.5kD [U4/U6.U5] tri-snRNP protein with the 5’ stem-loop of U4 snRNA. EMBO J. 1999, 18, 6119–6133. [Google Scholar] [CrossRef] [Green Version]
- Vidovic, I.; Nottrott, S.; Hartmuth, K.; Lührmann, R.; Ficner, R. Crystal structure of the spliceosomal 15.5kD protein bound to a U4 snRNA fragment. Mol. Cell 2000, 6, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Trinh-Rohlik, Q.; Maxwell, E.S. Homologous genes for mouse 4.5S hybRNA are found in all eukaryotes and their low molecular weight RNA transcripts intermolecularly hybridize with eukaryotic 18S ribosomal RNAs. Nucleic Acids Res. 1988, 16, 6041–6056. [Google Scholar] [CrossRef]
- Qu, L.H.; Henry, Y.; Nicoloso, M.; Michot, B.; Azum, M.C.; Renalier, M.H.; Caizergues-Ferrer, M.; Bachellerie, J.P. U24, a novel intron-encoded small nucleolar RNA with two 12 nt long, phylogenetically conserved complementarities to 28S rRNA. Nucleic Acids Res. 1995, 23, 2669–2676. [Google Scholar] [CrossRef]
- Nicoloso, M.; Qu, L.H.; Michot, B.; Bachellerie, J.P. Intron-encoded, antisense small nucleolar RNAs: The characterization of nine novel species points to their direct role as guides for the 2’-O-ribose methylation of rRNAs. J. Mol. Biol 1996, 260, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.-H.; Nicoloso, M.; Michot, B.; Azum, M.-C.; Caizergues-Ferrer, M.; Renalier, M.-H.; Bachellerie, J.-P. U21, a novel small nucleolar RNA with a 13 nt. complementarity to 28S rRNA, is encoded in an intron of ribosomal protein L5 gene in chicken and mammals. Nucleic Acids Res. 1994, 22, 4073–4081. [Google Scholar] [CrossRef] [PubMed]
- Bachellerie, J.-P.; Michot, B.; Nicoloso, M.; Balakin, A.; Ni, J.; Fournier, M.J. Antisense snoRNAs: A family of nucleolar RNAs with long complementarities to rRNA. Trends Biochem. Sci. 1995, 20, 261–264. [Google Scholar] [CrossRef]
- Cavaille, J.; Nicoloso, M.; Bachellerie, J.P. Targeted ribose methylation of RNA in vivo directed by tailored antisense RNA guides. Nature 1996, 383, 732–735. [Google Scholar] [CrossRef]
- Tycowski, K.T.; Smith, C.M.; Shu, M.-D.; Steitz, J.A. A small nucleolar RNA requirement for site-specific ribose methylation of rRNA in Xenopus. Proc. Natl. Acad. Sci. USA 1996, 93, 14480–14485. [Google Scholar] [CrossRef] [Green Version]
- Kiss-Laszlo, Z.; Henry, Y.; Kiss, T. Sequence and structural elements of methylation guide snoRNAs essential for site-specific ribose methylation of pre-rRNA. EMBO J. 1998, 17, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Lai, S.; Jia, R.; Xu, A.; Zhang, L.; Lu, J.; Ye, K. Structural basis for site-specific ribose methylation by box C/D RNA protein complexes. Nature 2011, 469, 559–563. [Google Scholar] [CrossRef]
- Ganot, P.; Bortolin, M.L.; Kiss, T. Site-specific pseudouridine formation in preribosomal RNA is guided by small nucleolar RNAs. Cell 1997, 89, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Tien, A.L.; Fournier, M.J. Small nucleolar RNAs direct site-specific synthesis of pseudouridine in ribosomal RNA. Cell 1997, 89, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Balakin, A.G.; Smith, L.; Fournier, M.J. The RNA world of the nucleolus: Two major families of small RNAs defined by different box elements with related functions. Cell 1996, 86, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Ganot, P.; Caizergues-Ferrer, M.; Kiss, T. The family of box ACA small nucleolar RNAs is defined by an evolutionarily conserved secondary structure and ubiquitous sequence elements essential for RNA accumulation. Genes. Dev. 1997, 11, 941–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolin, M.L.; Ganot, P.; Kiss, T. Elements essential for accumulation and function of small nucleolar RNAs directing site-specific pseudouridylation of ribosomal RNAs. EMBO J. 1999, 18, 457–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Zoysa, M.D.; Wu, G.; Katz, R.; Yu, Y.-T. Guide-substrate base-pairing requirement for box H/ACA RNA-guided RNA pseudouridylation. RNA 2018, 24, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.K.; Czekay, D.P.; Kothe, U. Base-pairing interactions between substrate RNA and H/ACA guide RNA modulate the kinetics of pseudouridylation, but not the affinity of substrate binding by H/ACA small nucleolar ribonucleoproteins. RNA 2019, 25, 1393–1404. [Google Scholar] [CrossRef] [Green Version]
- Girard, J.-P.; Lehtonen, H.; Caizergues-Ferrer, M.; Amalric, F.; Tollervey, D.; Lapeyre, B. GAR1 is an essential small nucleolar RNP protein required for pre-rRNA processing in yeast. EMBO J. 1992, 11, 673–682. [Google Scholar] [CrossRef]
- Bousquet-Antonelli, C.; Henry, Y.; Gélugne, J.; Caizergues-Ferrer, M.; Kiss, T. A small nucleolar RNP protein is required for pseudouridylation of eukaryotic ribosomal RNAs. EMBO J. 1997, 16, 4770–4776. [Google Scholar] [CrossRef]
- Lafontaine, D.L.; Bousquet-Antonelli, C.; Henry, Y.; Caizergues-Ferrer, M.; Tollervey, D. The box H + ACA snoRNAs carry Cbf5p, the putative rRNA pseudouridine synthase. Genes. Dev. 1998, 12, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Watkins, N.J.; Gottschalk, A.; Neubauer, G.; Kastner, B.; Fabrizio, P.; Mann, M.; Luehrmann, R. Cbf5p, a potential pseudouridine synthase, and Nhp2p, a putative RNA-binding protein, are present together with Gar1p in all H BOX/ACA-motif snoRNPs and constitute a common bipartite structure. RNA 1998, 4, 1549–1568. [Google Scholar] [CrossRef] [Green Version]
- Henras, A.; Henry, Y.; Bousquet-Antonelli, C.; Noaillac-Depeyre, J.; Gélugne, J.P.; Caizergues-Ferrer, M. Nhp2p and Nop10p are essential for the function of H/ACA snoRNPs. EMBO J. 1998, 17, 7078–7090. [Google Scholar] [CrossRef] [Green Version]
- Caton, E.A.; Kelly, E.K.; Kamalampeta, R.; Kothe, U. Efficient RNA pseudouridylation by eukaryotic H/ACA ribonucleoproteins requires high affinity binding and correct positioning of guide RNA. Nucleic Acids Res. 2018, 46, 905–916. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.-H.; Henras, A.; Lu, Y.-J.; Zhou, H.; Zhou, W.-X.; Zhu, Y.-Q.; Zhao, J.; Henry, Y.; Caizergues-Ferrer, M.; Bachellerie, J.-P. Seven novel methylation guide small nucleolar RNAs are processed from a common polycistronic transcript by Rat1p and RNase III in yeast. Mol. Cell. Biol. 1999, 19, 1144–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosio, M.C.; Fermi, B.; Dieci, G. Transcriptional control of yeast ribosome biogenesis: A multifaceted role for general regulatory factors. Transcription 2017, 8, 254–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preti, M.; Ribeyre, C.; Pascali, C.; Bosio, M.C.; Cortelazzi, B.; Rougemont, J.; Guarnera, E.; Naef, F.; Shore, D.; Dieci, G. The telomere-binding protein Tbf1 demarcates snoRNA gene promoters in Saccharomyces cerevisiae. Mol. Cell 2010, 38, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhou, X.; Wang, X.; Zhu, D.; Zhang, Y. Identification and characterization of human snoRNA core promoters. Genomics 2010, 96, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Harismendy, O.; Gendrel, C.; Soularue, P.; Gidrol, X.; Sentenac, A.; Werner, M.; Lefebvre, O. Genome-wide location of yeast RNA polymerase III transcription machinery. EMBO J. 2003, 22, 4738–4747. [Google Scholar] [CrossRef] [Green Version]
- Moqtaderi, Z.; Struhl, K. Genome-wide occupancy profile of the RNA polymerase III machinery in Saccharomyces cerevisiae reveals loci with incomplete transcription complexes. Mol. Cell Biol. 2004, 24, 4118–4127. [Google Scholar] [CrossRef] [Green Version]
- Roberts, U.N.; Stewart, A.J.; Huff, J.; Cairns, B.R. The RNA polymerase III transcriptome revealed by genome-wide localization and activity-occupancy relationships. Proc. Natl. Acad. Sci. USA 2003, 100, 14695–14700. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, K.; Lieb, J.D. Integral nuclear pore proteins bind to Pol III-transcribed genes and are required for Pol III transcript processing in C. elegans. Mol. Cell 2013, 51, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Isogai, Y.; Takada, S.; Tjian, R.; Keleş, S. Novel TRF1/BRF target genes revealed by genome-wide analysis of Drosophila Pol III transcription. EMBO J. 2007, 26, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Hellung-Larsen, P.; Jensen, E.G.; Frederiksen, S. Effect of 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole on the synthesis of low molecular weight, RNA components. Biochem. Biophys. Res. Commun. 1981, 99, 1303–1310. [Google Scholar] [CrossRef]
- Hellung-Larsen, P.; Kulamowicz, I.; Frederiksen, S. Synthesis of low molecular weight RNA components in cells with a temperature-sensitive polymerase II. Biochim. Biophys. Acta 1980, 609, 201–204. [Google Scholar] [CrossRef]
- Chandrasekharappa, S.C.; Smith, J.H.; Eliceiri, G.L. Biosynthesis of small nuclear RNAs in human cells. J. Cell Physiol. 1983, 117, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Marshallsay, C.; Filipowicz, W. Alteration of the RNA polymerase specificity of U3 snRNA genes during evolution and in vitro. Cell 1991, 65, 517–526. [Google Scholar] [CrossRef]
- Yuan, Y.; Reddy, R. 5’ flanking sequences of human MRP/7-2 RNA gene are required and sufficient for the transcription by RNA polymerase III. Biochim. Biophys. Acta 1991, 1089, 33–39. [Google Scholar] [CrossRef]
- Schmitt, M.E.; Clayton, D.A. Yeast site-specific ribonucleoprotein endoribonuclease MRP contains an RNA component homologous to mammalian RNase MRP RNA and essential for cell viability. Genes Dev. 1992, 6, 1975–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kufel, J.; Grzechnik, P. Small Nucleolar RNAs Tell a Different Tale. Trends Genet. 2019, 35, 104–117. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, E.J.; Conrad, N.K.; A Brow, D.; Corden, J.L. RNA-binding protein Nrd1 directs poly(A)-independent 3’-end formation of RNA polymerase II transcripts. Nature 2001, 413, 327–331. [Google Scholar] [CrossRef]
- Grzechnik, P.; Gdula, M.R.; Proudfoot, N.J. Pcf11 orchestrates transcription termination pathways in yeast. Genes Dev. 2015, 29, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Van Hoof, A.; Lennertz, P.; Parker, R. Yeast exosome mutants accumulate 3’-extended polyadenylated forms of U4 small nuclear RNA and small nucleolar RNAs. Mol. Cell Biol. 2000, 20, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Chanfreau, G.; Legrain, P.; Jacquier, A. Yeast RNase III as a key processing enzyme in small nucleolar RNAs metabolism. J. Mol. Biol. 1998, 284, 975–988. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Lee, A.; Chanfreau, G. The roles of endonucleolytic cleavage and exonucleolytic digestion in the 5’-end processing of S. cerevisiae box C/D snoRNAs. RNA 2003, 9, 1362–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzechnik, P.; Szczepaniak, S.A.; Dhir, S.; Pastucha, A.; Parslow, H.; Matuszek, Z.; Mischo, H.E.; Kufel, J.; Proudfoot, N.J. Nuclear fate of yeast snoRNA is determined by co-transcriptional Rnt1 cleavage. Nat. Commun. 2018, 9, 1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Maxwell, E.S. Mouse U14 snRNA is encoded in an intron of the mouse cognate hsc70 heat shock gene. Nucleic Acids Res. 1990, 18, 6565–6571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leverette, R.D.; Andrews, M.T.; Maxwell, E.S. Mouse U14 snRNA is a processed intron of the cognate hsc70 heat shock pre-messenger RNA. Cell 1992, 71, 1215–1221. [Google Scholar] [CrossRef]
- Tycowski, K.T.; Shu, M.D.; Steitz, J.A. A small nucleolar RNA is processed from an intron of the human gene encoding ribosomal protein S3. Genes Dev. 1993, 7, 1176–1190. [Google Scholar] [CrossRef]
- Bond, U.M.; Yario, T.A.; Steitz, J.A. Multiple processing-defective mutations in a mammalian histone pre-mRNA are suppressed by compensatory changes in U7 RNA both in vivo and in vitro. Genes Dev. 1991, 5, 1709–1722. [Google Scholar] [CrossRef] [Green Version]
- Bizarro, J.; Charron, C.; Boulon, S.; Westman, B.; Pradet-Balade, B.; Vandermoere, F.; Chagot, M.-E.; Hallais, M.; Ahmad, Y.; Leonhardt, H.; et al. Proteomic and 3D structure analyses highlight the C/D box snoRNP assembly mechanism and its control. J. Cell Biol 2014, 207, 463–480. [Google Scholar] [CrossRef]
- Tycowski, K.T.; Shu, M.D.; Steitz, J.A. A mammalian gene with introns instead of exons generating stable RNA products. Nature 1996, 379, 464–466. [Google Scholar] [CrossRef]
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5’-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [Green Version]
- Goustin, A.S.; Thepsuwan, P.; Kosir, M.A.; Lipovich, L. The Growth-Arrest-Specific (GAS)-5 Long Non-Coding RNA: A Fascinating lncRNA Widely Expressed in Cancers. Noncoding RNA 2019, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Pelczar, P.; Filipowicz, W. The host gene for intronic U17 small nucleolar RNAs in mammals has no protein-coding potential and is a member of the 5’-terminal oligopyrimidine gene family. Mol. Cell Biol. 1998, 18, 4509–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolin, M.L.; Kiss, T. Human U19 intron-encoded snoRNA is processed from a long primary transcript that possesses little potential for protein coding. RNA 1998, 4, 445–454. [Google Scholar] [PubMed]
- Tanaka, R.; Satoh, H.; Moriyama, M.; Satoh, K.; Morishita, Y.; Yoshida, S.; Watanabe, T.; Nakamura, Y.; Mori, S. Intronic U50 small-nucleolar-RNA (snoRNA) host gene of no protein-coding potential is mapped at the chromosome breakpoint t(3;6)(q27;q15) of human B-cell lymphoma. Genes Cells 2000, 5, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Thoreen, C.C.; Chantranupong, L.; Keys, H.R.; Wang, T.C.; Gray, N.S.; Sabatini, D.M. A unifying model for mTORC1-mediated regulation of mRNA translation. Nature 2012, 485, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Hirose, T.; Shu, M.D.; Steitz, J.A. Splicing-dependent and -independent modes of assembly for intron-encoded box C/D snoRNPs in mammalian cells. Mol. Cell 2003, 12, 113–123. [Google Scholar] [CrossRef]
- Ooi, S.L.; Samarsky, D.A.; Fournier, M.J.; Boeke, J.D. Intronic snoRNA biosynthesis in Saccharomyces cerevisiae depends on the lariat-debranching enzyme: Intron length effects and activity of a precursor snoRNA. RNA 1998, 4, 1096–1110. [Google Scholar] [CrossRef] [Green Version]
- Kiss, T.; Filipowicz, W. Exonucleolytic processing of small nucleolar RNAs from pre-mRNA introns. Genes Dev. 1995, 9, 1411–1424. [Google Scholar] [CrossRef] [Green Version]
- Massenet, S.; Bertrand, E.; Verheggen, C. Assembly and trafficking of box C/D and H/ACA snoRNPs. RNA Biol. 2017, 14, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Davey, M.; Hsu, Y.-C.; Kaplanek, P.; Tong, A.; Parsons, A.B.; Krogan, N.; Cagney, G.; Mai, D.; Greenblatt, J.; et al. Navigating the chaperone network: An integrative map of physical and genetic interactions mediated by the hsp90 chaperone. Cell 2005, 120, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Te, J.; Jia, L.; Rogers, J.; Miller, A.; Hartson, S.D. Novel subunits of the mammalian Hsp90 signal transduction chaperone. J. Proteome Res. 2007, 6, 1963–1973. [Google Scholar] [CrossRef]
- Zhao, R.; Kakihara, Y.; Gribun, A.; Huen, J.; Yang, G.; Khanna, M.; Costanzo, M.; Brost, R.L.; Boone, C.; Hughes, T.R. Molecular chaperone Hsp90 stabilizes Pih1/Nop17 to maintain R2TP complex activity that regulates snoRNA accumulation. J. Cell Biol. 2008, 180, 563–578. [Google Scholar] [CrossRef] [PubMed]
- King, T.H.; Decatur, W.A.; Bertrand, E.; Maxwell, E.S.; Fournier, M.J. A well-connected and conserved nucleoplasmic helicase is required for production of box C/D and H/ACA snoRNAs and localization of snoRNP proteins. Mol. Cell Biol. 2001, 21, 7731–7746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zebarjadian, Y.; King, T.; Fournier, M.J.; Clarke, L.; Carbon, J. Point mutations in yeast CBF5 can abolish in vivo pseudouridylation of rRNA. Mol. Cell Biol. 1999, 19, 7461–7472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jack, K.; Bellodi, C.; Landry, R.M.; Niederer, R.O.; Meskauskas, A.; Musalgaonkar, S.; Kopmar, N.; Krasnykh, O.; Dean, A.M.; Thompson, S.; et al. rRNA pseudouridylation defects affect ribosomal ligand binding and translational fidelity from yeast to human cells. Mol. Cell 2011, 44, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Esguerra, J.; Warringer, J.; Blomberg, A. Functional importance of individual rRNA 2’-O-ribose methylations revealed by high-resolution phenotyping. RNA 2008, 14, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Tollervey, D.; Guthrie, C. Deletion of a yeast small nuclear RNA gene impairs growth. EMBO J. 1985, 4, 3873–3878. [Google Scholar] [CrossRef]
- Tollervey, D. A yeast small nuclear RNA is required for normal processing of pre-ribosomal RNA. EMBO J. 1987, 6, 4169–4175. [Google Scholar] [CrossRef]
- King, T.H.; Liu, B.; McCully, R.R.; Fournier, M.J. Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center. Mol. Cell 2003, 11, 425–435. [Google Scholar] [CrossRef]
- Bonnerot, C.; Pintard, L.; Lutfalla, G. Functional redundancy of Spb1p and a snR52-dependent mechanism for the 2’-O-ribose methylation of a conserved rRNA position in yeast. Mol. Cell 2003, 12, 1309–1315. [Google Scholar] [CrossRef]
- Caldas, T.; Binet, E.; Bouloc, P.; Costa, A.; Desgres, J.; Richarme, G. The FtsJ/RrmJ heat shock protein of Escherichia coli is a 23 S ribosomal RNA methyltransferase. J. Biol. Chem. 2000, 275, 16414–16419. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shem, A.; De Loubresse, N.G.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.H.; Liu, Q.; Fournier, M.J. rRNA modifications in an intersubunit bridge of the ribosome strongly affect both ribosome biogenesis and activity. Mol. Cell 2007, 28, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.H.; Liu, Q.; Fournier, M.J. Loss of rRNA modifications in the decoding center of the ribosome impairs translation and strongly delays pre-rRNA processing. RNA 2009, 15, 1716–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullineux, S.T.; Lafontaine, D.L. Mapping the cleavage sites on mammalian pre-rRNAs: Where do we stand? Biochimie 2012, 94, 1521–1532. [Google Scholar] [CrossRef] [PubMed]
- Lestrade, L.; Weber, M.J. snoRNA-LBME-db, a comprehensive database of human H/ACA and C/D box snoRNAs. Nucleic Acids Res. 2006, 34, D158–D162. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Prestayko, A.W.; Busch, H. Studies on nucleolar 4 to 6 S ribonucleic acid of Novikoff hepatoma cells. J. Biol. Chem. 1968, 243, 1368–1375. [Google Scholar]
- Hodnett, J.L.; Busch, H. Isolation and characterization of uridylic acid-rich 7 S ribonucleic acid of rat liver nuclei. J. Biol. Chem. 1968, 243, 6334–6342. [Google Scholar]
- Prestayko, A.W.; Tonato, M.; Busch, H. Low molecular weight RNA associated with 28 s nucleolar RNA. J. Mol. Biol. 1970, 47, 505–515. [Google Scholar] [CrossRef]
- Weinberg, R.A.; Penman, S. Small molecular weight monodisperse nuclear RNA. J. Mol. Biol. 1968, 38, 289–304. [Google Scholar] [CrossRef]
- Reddy, R.; Ro-Choi, T.S.; Henning, D.; Shibata, H.; Choi, Y.C.; Busch, H.H. Modified nucleosides of nuclear and nucleolar low molecular weight ribonucleic acid. J. Biol. Chem. 1972, 247, 7245–7250. [Google Scholar]
- Stroke, I.L.; Weiner, A.M. Genes and pseudogenes for rat U3A and U3B small nuclear RNA. J. Mol. Biol. 1985, 184, 183–193. [Google Scholar] [CrossRef]
- Suh, D.; Busch, H.; Reddy, R. Isolation and characterization of a human U3 small nucleolar RNA gene. Biochem. Biophys. Res. Commun. 1986, 137, 1133–1140. [Google Scholar] [CrossRef]
- Calvet, J.P.; Pederson, T. Base-pairing interactions between small nuclear RNAs and nuclear RNA precursors as revealed by psoralen cross-linking in vivo. Cell 1981, 26 Pt 1, 363–370. [Google Scholar] [CrossRef]
- Maser, R.L.; Calvet, J.P. U3 small nuclear RNA can be psoralen-cross-linked in vivo to the 5’ external transcribed spacer of pre-ribosomal-RNA. Proc. Natl. Acad. Sci. USA 1989, 86, 6523–6527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrame, M.; Tollervey, D. Identification and functional analysis of two U3 binding sites on yeast pre-ribosomal RNA. EMBO J. 1992, 11, 1531–1542. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhu, X.; Qi, J.; An, W.; Lan, P.; Tan, D.; Chen, R.; Wang, B.; Zheng, S.; Zhang, C. Molecular architecture of the 90S small subunit pre-ribosome. Elife 2017, 6, e22086. [Google Scholar] [CrossRef]
- Crouch, R.J.; Kanaya, S.; Earl, P.L. A model for the involvement of the small nucleolar RNA (U3) in processing eukaryotic ribosomal RNA. Mol. Biol. Rep. 1983, 9, 75–78. [Google Scholar] [CrossRef]
- Bachellerie, J.P.; Michot, B.; Raynal, F. Recognition signals for mouse pre-rRNA processing. A potential role for U3 nucleolar RNA. Mol. Biol. Rep. 1983, 9, 79–86. [Google Scholar] [CrossRef]
- Tague, B.W.; Gerbi, S.A. Processing of the large rRNA precursor: Two proposed categories of RNA-RNA interactions in eukaryotes. J. Mol. Evol 1984, 20, 362–367. [Google Scholar] [CrossRef]
- Parker, K.A.; Steitz, J.A. Structural analysis of the human U3 ribonucleoprotein particle reveal a conserved sequence available for base pairing with pre-rRNA. Mol. Cell Biol. 1987, 7, 2899–2913. [Google Scholar] [CrossRef] [Green Version]
- Kass, S.; Tyc, K.; Steitz, J.A.; Sollner-Webb, B. The U3 small nucleolar ribonucleoprotein functions in the first step of preribosomal RNA processing. Cell 1990, 60, 897–908. [Google Scholar] [CrossRef]
- Savino, R.; Gerbi, S.A. In vivo disruption of Xenopus U3 snRNA affects ribosomal RNA processing. EMBO J. 1990, 9, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.M.; Ares, M., Jr. Depletion of U3 small nucleolar RNA inhibits cleavage in the 5’ external transcribed spacer of yeast pre-ribosomal RNA and impairs formation of 18S ribosomal RNA. EMBO J. 1991, 10, 4231–4239. [Google Scholar] [CrossRef] [PubMed]
- Tollervey, D.; Lehtonen, H.; Carmo-Fonseca, M.; Hurt, E.C. The small nucleolar RNP protein NOP1 (fibrillarin) is required for pre-rRNA processing in yeast. EMBO J. 1991, 10, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Lübben, B.; Marshallsay, C.; Rottmann, N.; Lührmann, R. Isolation of U3 snoRNP from CHO cells: A novel 55 kDa protein binds to the central part of U3 snoRNA. Nucleic Acids Res. 1993, 21, 5377–5385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venema, J.; Vos, H.; Faber, A.W.; Van Venrooij, W.J.; Raué, H.A. Yeast Rrp9p is an evolutionarily conserved U3 snoRNP protein essential for early pre-rRNA processing cleavages and requires box C for its association. RNA 2000, 6, 1660–1671. [Google Scholar] [CrossRef] [Green Version]
- Clerget, G.; Bourguignon-Igel, V.; Marmier-Gourrier, N.; Rolland, N.; Wacheul, L.; Manival, X.; Charron, C.; Kufel, J.; Méreau, A.; Senty-Ségault, V.; et al. Synergistic defects in pre-rRNA processing from mutations in the U3-specific protein Rrp9 and U3 snoRNA. Nucleic Acids Res. 2020, 48, 3848–3868. [Google Scholar] [CrossRef]
- Granneman, S.; Kudla, G.; Petfalski, E.; Tollervey, D. Identification of protein binding sites on U3 snoRNA and pre-rRNA by UV cross-linking and high-throughput analysis of cDNAs. Proc. Natl. Acad. Sci. USA 2009, 106, 9613–9618. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, D.A.; Wormsley, S.; Agentis, T.M.; Baserga, S.J. Mpp10p, a U3 small nucleolar ribonucleoprotein component required for pre-18S rRNA processing in yeast. Mol. Cell Biol. 1997, 17, 5803–5812. [Google Scholar] [CrossRef] [Green Version]
- Dragon, F.; Gallagher, J.E.G.; Compagnone-Post, P.A.; Mitchell, B.M.; Porwancher, K.A.; Wehner, K.A.; Wormsley, S.; Settlage, R.E.; Shabanowitz, J.; Osheim, Y.; et al. A large nucleolar U3 ribonucleoprotein required for 18S ribosomal RNA biogenesis. Nature 2002, 417, 967–970. [Google Scholar] [CrossRef]
- Miller, O.L., Jr.; Beatty, B.R. Visualization of nucleolar genes. Science 1969, 164, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Mougey, E.B.; O’Reilly, M.; Osheim, Y.; Sollner-Webb, B.; Miller, O.L.; Beyer, A. The terminal balls characteristic of eukaryotic rRNA transcription units in chromatin spreads are rRNA processing complexes. Genes Dev. 1993, 7, 1609–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornprobst, M.; Turk, M.; Kellner, N.; Cheng, J.; Flemming, D.; Koš-Braun, I.; Koš, M.; Thoms, M.; Berninghausen, O.; Beckmann, R.; et al. Architecture of the 90S Pre-ribosome: A Structural View on the Birth of the Eukaryotic Ribosome. Cell 2016, 166, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaker-Margot, M.; Barandun, J.; Hunziker, M.; Klinge, S. Architecture of the yeast small subunit processome. Science 2017, 355, eaal1880. [Google Scholar] [CrossRef]
- Barandun, J.; Chaker-Margot, M.; Hunziker, M.; Molloy, K.R.; Chait, B.T.; Klinge, S. The complete structure of the small-subunit processome. Nat. Struct. Mol. Biol. 2017, 24, 944–953. [Google Scholar] [CrossRef]
- Cheng, J.; Kellner, N.; Berninghausen, O.; Hurt, E.; Beckmann, R. 3.2-A-resolution structure of the 90S preribosome before A1 pre-rRNA cleavage. Nat. Struct. Mol. Biol. 2017, 24, 954–964. [Google Scholar] [CrossRef]
- Goldfarb, K.C.; Cech, T.R. Targeted CRISPR disruption reveals a role for RNase MRP RNA in human preribosomal RNA processing. Genes Dev. 2017, 31, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.D.; Clayton, D.A. A novel endoribonuclease cleaves at a priming site of mouse mitochondrial DNA replication. EMBO J. 1987, 6, 409–417. [Google Scholar] [CrossRef]
- Chang, D.D.; Clayton, D.A. A mammalian mitochondrial RNA processing activity contains nucleus-encoded RNA. Science 1987, 235, 1178–1184. [Google Scholar] [CrossRef]
- Chang, D.D.; Clayton, D.A. Mouse RNAase MRP RNA is encoded by a nuclear gene and contains a decamer sequence complementary to a conserved region of mitochondrial RNA substrate. Cell 1989, 56, 131–139. [Google Scholar] [CrossRef]
- Clayton, D.A. A nuclear function for RNase MRP. Proc. Natl. Acad. Sci. USA 1994, 91, 4615–4617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topper, J.N.; Clayton, D.A. Characterization of human MRP/Th RNA and its nuclear gene: Full length MRP/Th RNA is an active endoribonuclease when assembled as an RNP. Nucleic Acids Res. 1990, 18, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, M.E.; Bennett, J.L.; Dairaghi, D.J.; Clayton, D.A. Secondary structure of RNase MRP RNA as predicted by phylogenetic comparison. FASEB J. 1993, 7, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.C.; Avis, J.M. A conserved element in the yeast RNase MRP RNA subunit can participate in a long-range base-pairing interaction. J. Mol. Biol. 2004, 341, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, L.; Fretz, S.; Epps, N.; Zengel, J.M. Functional equivalence of hairpins in the RNA subunits of RNase MRP and RNase P in Saccharomyces cerevisiae. RNA 2000, 6, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Gold, H.; Topper, J.; Clayton, D.; Craft, J. The RNA processing enzyme RNase MRP is identical to the Th RNP and related to RNase P. Science 1989, 245, 1377–1380. [Google Scholar] [CrossRef]
- Reddy, R.; Li, W.Y.; Henning, D.; Choi, Y.C.; Nohga, K.; Busch, H. Characterization and subcellular localization of 7-8 S RNAs of Novikoff hepatoma. J. Biol. Chem. 1981, 256, 8452–8457. [Google Scholar]
- Hashimoto, C.; Steitz, J.A. Sequential association of nucleolar 7-2 RNA with two different autoantigens. J. Biol. Chem. 1983, 258, 1379–1382. [Google Scholar]
- Reimer, G.; Raška, I.; Scheer, U.; Tan, E.M. Immunolocalization of 7-2-ribonucleoprotein in the granular component of the nucleolus. Exp. Cell Res. 1988, 176, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Lygerou, Z.; Allmang, C.; Tollervey, D.; Séraphin, B. Accurate processing of a eukaryotic precursor ribosomal RNA by ribonuclease MRP in vitro. Science 1996, 272, 268–270. [Google Scholar] [CrossRef]
- Schmitt, M.E.; Clayton, D.A. Nuclear RNase MRP is required for correct processing of pre-5.8S rRNA in Saccharomyces cerevisiae. Mol. Cell Biol. 1993, 13, 7935–7941. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Archer, R.H.; Zengel, J.M.; Lindahl, L. The RNA of RNase MRP is required for normal processing of ribosomal RNA. Proc. Natl. Acad. Sci. USA 1994, 91, 659–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, L.; Archer, R.H.; Zengel, J.M. A new rRNA processing mutant of Saccharomyces cerevisiae. Nucleic Acids Res. 1992, 20, 295–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, M.E.; Clayton, D.A. Characterization of a unique protein component of yeast RNase MRP: An RNA-binding protein with a zinc-cluster domain. Genes Dev. 1994, 8, 2617–2628. [Google Scholar] [CrossRef] [Green Version]
- Salinas, K.; Wierzbicki, S.; Zhou, L.; Schmitt, M.E. Characterization and purification of Saccharomyces cerevisiae RNase MRP reveals a new unique protein component. J. Biol. Chem. 2005, 280, 11352–11360. [Google Scholar] [CrossRef] [Green Version]
- Reich, C.; Olsen, G.; Pace, B.; Pace, N. Role of the protein moiety of ribonuclease P, a ribonucleoprotein enzyme. Science 1988, 239, 178–181. [Google Scholar] [CrossRef]
- Cai, T.; Aulds, J.; Gill, T.; Cerio, M.; E Schmitt, M. The Saccharomyces cerevisiae RNase mitochondrial RNA processing is critical for cell cycle progression at the end of mitosis. Genetics 2002, 161, 1029–1042. [Google Scholar]
- Gill, T.; Cai, T.; Aulds, J.; Wierzbicki, S.; Schmitt, M.E. RNase MRP cleaves the CLB2 mRNA to promote cell cycle progression: Novel method of mRNA degradation. Mol. Cell Biol. 2004, 24, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Peculis, B.A.; Steitz, J.A. Disruption of U8 nucleolar snRNA inhibits 5.8S and 28S rRNA processing in the Xenopus oocyte. Cell 1993, 73, 1233–1245. [Google Scholar] [CrossRef]
- Srivastava, L.; Lapik, Y.R.; Wang, M.; Pestov, D.G. Mammalian DEAD box protein Ddx51 acts in 3’ end maturation of 28S rRNA by promoting the release of U8 snoRNA. Mol. Cell Biol. 2010, 30, 2947–2956. [Google Scholar] [CrossRef] [Green Version]
- Langhendries, J.-L.; Nicolas, E.; Doumont, G.; Goldman, S.; Lafontaine, D.L. The human box C/D snoRNAs U3 and U8 are required for pre-rRNA processing and tumorigenesis. Oncotarget 2016, 7, 59519–59534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cote, C.A.; Greer, C.L.; Peculis, B.A. Dynamic conformational model for the role of ITS2 in pre-rRNA processing in yeast. RNA 2002, 8, 786–797. [Google Scholar] [PubMed] [Green Version]
- Cavaille, J.; Hadjiolov, A.A.; Bachellerie, J.P. Processing of mammalian rRNA precursors at the 3’ end of 18S rRNA. Identification of cis-acting signals suggests the involvement of U13 small nucleolar RNA. Eur. J. Biochem. 1996, 242, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Horikawa, S.; Suzuki, T.; Kawauchi, H.; Tanaka, Y.; Suzuki, T.; Suzuki, T. Human NAT10 is an ATP-dependent RNA acetyltransferase responsible for N4-acetylcytidine formation in 18 S ribosomal RNA (rRNA). J. Biol. Chem. 2014, 289, 35724–35730. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Langhendries, J.L.; Watzinger, P.; Kötter, P.; Entian, K.D.; Lafontaine, D.L. Yeast Kre33 and human NAT10 are conserved 18S rRNA cytosine acetyltransferases that modify tRNAs assisted by the adaptor Tan1/THUMPD1. Nucleic Acids Res. 2015, 43, 2242–2258. [Google Scholar] [CrossRef] [Green Version]
- Li, H.D.; Zagorski, J.; Fournier, M.J. Depletion of U14 small nuclear RNA (snR128) disrupts production of 18S rRNA in Saccharomyces cerevisiae. Mol. Cell Biol. 1990, 10, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.Q.; Fournier, M.J. U14 base-pairs with 18S rRNA: A novel snoRNA interaction required for rRNA processing. Genes Dev. 1995, 9, 2433–2443. [Google Scholar] [CrossRef] [Green Version]
- Samarsky, D.A.; Schneider, G.S.; Fournier, M.J. An essential domain in Saccharomyces cerevisiae U14 snoRNA is absent in vertebrates, but conserved in other yeasts. Nucleic Acids Res. 1996, 24, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Jarmolowski, A.; Zagorski, J.; Li, H.V.; Fournier, M.J. Identification of essential elements in U14 RNA of Saccharomyces cerevisiae. EMBO J. 1990, 9, 4503–4509. [Google Scholar] [CrossRef]
- Dunbar, D.A.; Baserga, S.J. The U14 snoRNA is required for 2’-O-methylation of the pre-18S rRNA in Xenopus oocytes. RNA 1998, 4, 195–204. [Google Scholar]
- Tycowski, K.T.; Shu, M.D.; Steitz, J.A. Requirement for intron-encoded U22 small nucleolar RNA in 18S ribosomal RNA maturation. Science 1994, 266, 1558–1561. [Google Scholar] [CrossRef] [PubMed]
- Enright, C.A.; Maxwell, E.S.; Eliceiri, G.L.; Sollner-Webb, B. 5’ETS rRNA processing facilitated by four small RNAs: U14, E3, U17, and U3. RNA 1996, 2, 1094–1099. [Google Scholar]
- Atzorn, V.; Fragapane, P.; Kiss, T. U17/snR30 is a ubiquitous snoRNA with two conserved sequence motifs essential for 18S rRNA production. Mol. Cell Biol. 2004, 24, 1769–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, R.K.; Eliceiri, G.L. Three small nucleolar RNAs that are involved in ribosomal RNA precursor processing. Proc. Natl. Acad. Sci. USA 1997, 94, 4972–4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, M.H.; Soeiro, R.; Warner, J.R.; Darnell, J.E. The effects of methionine deprivation on ribosome synthesis in HeLa cells. Proc. Natl. Acad. Sci. USA 1967, 58, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swann, P.F. The effect of ethionine on ribonucleic acid synthesis in rat liver. Biochem. J. 1975, 150, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Caboche, M.; Bachellerie, J.P. RNA methylation and control of eukaryotic RNA biosynthesis. Effects of cycloleucine, a specific inhibitor of methylation, on ribosomal RNA maturation. Eur. J. Biochem. 1977, 74, 19–29. [Google Scholar] [CrossRef]
- Chelbi-Alix, M.K.; Expert-Bezançon, A.; Hayes, F.; Alix, J.H.; Branlant, C. Properties of ribosomes and ribosomal RNAs synthesized by Escherichia coli grown in the presence of ethionine. Normal maturation of ribosomal RNA in the absence of methylation. Eur. J. Biochem. 1981, 115, 627–634. [Google Scholar] [CrossRef]
- Jiang, W.; Middleton, K.; Yoon, H.J.; Fouquet, C.; Carbon, J. An essential yeast protein, CBF5p, binds in vitro to centromeres and microtubules. Mol. Cell Biol. 1993, 13, 4884–4893. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Navarrete, S.; Jasinski, M.; Vulliamy, T.; Dokal, I.; Bessler, M.; Mason, P.J. Targeted disruption of Dkc1, the gene mutated in X-linked dyskeratosis congenita, causes embryonic lethality in mice. Oncogene 2002, 21, 7740–7744. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.-W.; Ge, J.; Fan, J.-M.; Bessler, M.; Mason, P.J. Slow growth and unstable ribosomal RNA lacking pseudouridine in mouse embryonic fibroblast cells expressing catalytically inactive dyskerin. FEBS Lett. 2013, 587, 2112–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, R.C.; Klootwijk, J.; Planta, R.J.; Maden, B.E.H. Biosynthesis of a hypermodified nucleotide in Saccharomyces carlsbergensis 17S and HeLa-cell 18S ribosomal ribonucleic acid. Biochem. J. 1978, 169, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, B.A.; Ho, H.-T.; Ranganathan, S.V.; Vangaveti, S.; Ilkayeva, O.; Assi, H.A.; Choi, A.K.; Agris, P.F.; Holley, C. Modification of messenger RNA by 2’-O-methylation regulates gene expression in vivo. Nat. Commun. 2019, 10, 3401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nostramo, R.T.; Hopper, A.K. Beyond rRNA and snRNA: tRNA as a 2’-O-methylation target for nucleolar and Cajal body box C/D RNPs. Genes Dev. 2019, 33, 739–740. [Google Scholar] [CrossRef] [PubMed]
- A McKusick, V.; Eldridge, R.; A Hostetler, J.; Ruangwit, U.; A Egeland, J. Dwarfism in the Amish. Ii. Cartilage-Hair Hypoplasia. Bull. Johns. Hopkins. Hosp. 1965, 116, 285–326. [Google Scholar]
- Ridanpää, M.; Van Eenennaam, H.; Pelin, K.; Chadwick, R.; Johnson, C.; Yuan, B.; Vanvenrooij, W.; Pruijn, G.; Salmela, R.; Rockas, S.; et al. Mutations in the RNA component of RNase MRP cause a pleiotropic human disease, cartilage-hair hypoplasia. Cell 2001, 104, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Mattijssen, S.; Welting, T.J.; Pruijn, G.J. RNase MRP and disease. Wiley Interdiscip. Rev. RNA 2010, 1, 102–116. [Google Scholar] [CrossRef]
- Hermanns, P.; Bertuch, A.A.; Bertin, T.K.; Dawson, B.; Schmitt, M.E.; Shaw, C.A.; Zabel, B.; Lee, B. Consequences of mutations in the non-coding RMRP RNA in cartilage-hair hypoplasia. Hum. Mol. Genet. 2005, 14, 3723–3740. [Google Scholar] [CrossRef] [Green Version]
- Dokal, I. Dyskeratosis congenita. Hematol. Am. Soc. Hematol. Educ. Program 2011, 2011, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Walne, A.J.; Vulliamy, T.; Marrone, A.; Beswick, R.; Kirwan, M.; Masunari, Y.; Al-Qurashi, F.-H.; Aljurf, M.; Dokal, I. Genetic heterogeneity in autosomal recessive dyskeratosis congenita with one subtype due to mutations in the telomerase-associated protein NOP10. Hum. Mol. Genet. 2007, 16, 1619–1629. [Google Scholar] [CrossRef] [Green Version]
- Benyelles, M.; O’Donohue, M.-F.; Kermasson, L.; Lainey, E.; Borie, R.; Lagresle-Peyrou, C.; Nunes, H.; Cazelles, C.; Fourrage, C.; Ollivier, E.; et al. NHP2 deficiency impairs rRNA biogenesis and causes pulmonary fibrosis and Hoyeraal-Hreidarsson syndrome. Hum. Mol. Genet. 2020, 29, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, T.; Beswick, R.; Kirwan, M.; Marrone, A.; Digweed, M.; Walne, A.; Dokal, I. Mutations in the telomerase component NHP2 cause the premature ageing syndrome dyskeratosis congenita. Proc. Natl. Acad. Sci. USA 2008, 105, 8073–8078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thumati, N.R.; Zeng, X.-L.; Au, H.H.T.; Jang, C.; Jan, E.; Wong, J.M. Severity of X-linked dyskeratosis congenita (DKCX) cellular defects is not directly related to dyskerin (DKC1) activity in ribosomal RNA biogenesis or mRNA translation. Hum. Mutat. 2013, 34, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Ruggero, D.; Shimamura, A. Marrow failure: A window into ribosome biology. Blood 2014, 124, 2784–2792. [Google Scholar] [CrossRef] [Green Version]
- Heiss, N.S.; Knight, S.W.; Vulliamy, T.; Klauck, S.M.; Wiemann, S.; Mason, P.J.; Poustka, A.; Dokal, I. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat. Genet. 1998, 19, 32–38. [Google Scholar] [CrossRef]
- Mitchell, J.R.; Wood, E.; Collins, K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature 1999, 402, 551–555. [Google Scholar] [CrossRef]
- Ruggero, D.; Grisendi, S.; Piazza, F.; Rego, E.M.; Mari, F.; Rao, P.H.; Cordon-Cardo, C.; Pandolfi, P.P. Dyskeratosis congenita and cancer in mice deficient in ribosomal RNA modification. Science 2003, 299, 259–262. [Google Scholar] [CrossRef]
- Yoon, A.; Peng, G.; Brandenburg, Y.; Zollo, O.; Xu, W.; Rego, E.; Ruggero, D. Impaired control of IRES-mediated translation in X-linked dyskeratosis congenita. Science 2006, 312, 902–906. [Google Scholar] [CrossRef]
- Bellodi, C.; Kopmar, N.; Ruggero, D. Deregulation of oncogene-induced senescence and p53 translational control in X-linked dyskeratosis congenita. EMBO J. 2010, 29, 1865–1876. [Google Scholar] [CrossRef] [Green Version]
- Bellodi, C.; McMahon, M.; Contreras, A.; Juliano, D.; Kopmar, N.; Nakamura, T.; Maltby, D.; Burlingame, A.; Savage, S.A.; Shimamura, A. H/ACA small RNA dysfunctions in disease reveal key roles for noncoding RNA modifications in hematopoietic stem cell differentiation. Cell Rep. 2013, 3, 1493–1502. [Google Scholar] [CrossRef] [Green Version]
- Mills, E.W.; Green, R. Ribosomopathies: There’s strength in numbers. Science 2017, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachmani, D.; Bothmer, A.H.; Grisendi, S.; Mele, A.; Bothmer, D.; Lee, J.D.; Monteleone, E.; Cheng, K.; Zhang, Y.; Bester, A.C.; et al. Germline NPM1 mutations lead to altered rRNA 2’-O-methylation and cause dyskeratosis congenita. Nat. Genet. 2019, 51, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.; Buffa, F.M.; Camps, C.; Ramachandran, A.; Leek, R.; Taylor, M.; Patil, M.; Sheldon, H.; Betts, G.; Homer, J.; et al. The small-nucleolar RNAs commonly used for microRNA normalisation correlate with tumour pathology and prognosis. Br. J. Cancer 2011, 104, 1168–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.-M.; Thompson, I.; Yuan, Z.-M. Elevated snoRNA biogenesis is essential in breast cancer. Oncogene 2014, 33, 1348–1358. [Google Scholar] [CrossRef]
- Valleron, W.; Laprevotte, E.; Gautier, E.-F.; Quelen, C.; Demur, C.; Delabesse, E.; Agirre, X.; Prosper, F.; Kiss, T.; Brousset, P. Specific small nucleolar RNA expression profiles in acute leukemia. Leukemia 2012, 26, 2052–2060. [Google Scholar] [CrossRef] [Green Version]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-Y.; Guo, P.; Boyd, J.; Sun, X.; Li, Q.; Zhou, W.; Dong, J.-T. Implication of snoRNA U50 in human breast cancer. J. Genet. Genom. 2009, 36, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-Y.; Rodriguez, C.; Guo, P.; Sun, X.; Talbot, J.T.; Zhou, W.; Petros, J.; Li, Q.; Vessella, R.L.; Kibel, A.S.; et al. SnoRNA U50 is a candidate tumor-suppressor gene at 6q14.3 with a mutation associated with clinically significant prostate cancer. Hum. Mol. Genet. 2008, 17, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Siprashvili, Z.; Webster, D.E.; Johnston, D.; Shenoy, R.M.; Ungewickell, A.J.; Bhaduri, A.; Flockhart, R.; Zarnegar, B.J.; Che, Y.; Meschi, F.; et al. The noncoding RNAs SNORD50A and SNORD50B bind K-Ras and are recurrently deleted in human cancer. Nat. Genet. 2016, 48, 53–58. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.; Contreras, A.; Holm, M.; Uechi, T.; Forester, C.M.; Pang, X.; Jackson, C.; Calvert, M.E.; Chen, B.; Quigley, D.A.; et al. A single H/ACA small nucleolar RNA mediates tumor suppression downstream of oncogenic RAS. Elife 2019, 8. [Google Scholar]
- Liang, J.; Wen, J.; Huang, Z.; Chen, X.-P.; Zhang, B.-X.; Chu, L. Small Nucleolar RNAs: Insight Into Their Function in Cancer. Front. Oncol. 2019, 9, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, D.M.; Lu, C.; Green, P.J.; Parker, R. tRNA cleavage is a conserved response to oxidative stress in eukaryotes. RNA 2008, 14, 2095–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, S.; Ivanov, P.; Hu, G.-F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Feng, J.; Liu, Q.; Sun, F.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Stress induces tRNA cleavage by angiogenin in mammalian cells. FEBS Lett. 2009, 583, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Haussecker, D.; Huang, Y.; Lau, A.; Parameswaran, P.; Fire, A.Z.; Kay, M.A. Human tRNA-derived small RNAs in the global regulation of RNA silencing. RNA 2010, 16, 673–695. [Google Scholar] [CrossRef] [Green Version]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J. Small RNAs derived from snoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Kishore, S.; Khanna, A.; Zhang, Z.; Hui, J.; Balwierz, P.; Stefan, M.; Beach, C.; Nicholls, R.D.; Zavolan, M.; Stamm, S. The snoRNA MBII-52 (SNORD 115) is processed into smaller RNAs and regulates alternative splicing. Hum. Mol. Genet. 2010, 19, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Mleczko, A.M.; Machtel, P.; Walkowiak, M.; Wasilewska, A.; Pietras, P.J.; Bąkowska-Żywicka, K. Levels of sdRNAs in cytoplasm and their association with ribosomes are dependent upon stress conditions but independent from snoRNA expression. Sci. Rep. 2019, 9, 18397. [Google Scholar] [CrossRef] [Green Version]
- Lyons, S.; Achorn, C.; Kedersha, N.L.; Anderson, P.; Ivanov, P. YB-1 regulates tiRNA-induced Stress Granule formation but not translational repression. Nucleic Acids Res. 2016, 44, 6949–6960. [Google Scholar] [CrossRef]
- Blenkiron, C.; Hurley, D.; Fitzgerald, S.; Print, C.; Lasham, A. Links between the oncoprotein YB-1 and small non-coding RNAs in breast cancer. PLoS ONE 2013, 8, e80171. [Google Scholar] [CrossRef] [PubMed]
- Mordovkina, D.; Lyabin, D.; Smolin, E.; Sogorina, E.; Ovchinnikov, L.; Eliseeva, I. Y-Box Binding Proteins in mRNP Assembly, Translation, and Stability Control. Biomolecules 2020, 10, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, S.M.; Fay, M.M.; Ivanov, P. The role of RNA modifications in the regulation of tRNA cleavage. FEBS Lett. 2018, 592, 2828–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondrashov, N.; Pusic, A.; Stumpf, C.R.; Shimizu, K.; Hsieh, A.C.; Ishijima, J.; Shiroishi, T.; Barna, M. Ribosome-mediated specificity in Hox mRNA translation and vertebrate tissue patterning. Cell 2011, 145, 383–397. [Google Scholar] [CrossRef] [Green Version]
- Birkedal, U.; Christensen-Dalsgaard, M.; Krogh, N.; Sabarinathan, R.; Gorodkin, J.; Nielsen, H. Profiling of ribose methylations in RNA by high-throughput sequencing. Angew. Chem. Int. Ed. Engl. 2015, 54, 451–455. [Google Scholar] [CrossRef]
- Maden, B.E. Identification of the locations of the methyl groups in 18 S ribosomal RNA from Xenopus laevis and man. J. Mol. Biol. 1986, 189, 681–699. [Google Scholar] [CrossRef]
- Maden, B.E. Locations of methyl groups in 28 S rRNA of Xenopus laevis and man. Clustering in the conserved core of molecule. J. Mol. Biol. 1988, 201, 289–314. [Google Scholar] [CrossRef]
- Stanley, J.; Vassilenko, S. A different approach to RNA sequencing. Nature 1978, 274, 87–89. [Google Scholar] [CrossRef]
- Bakin, A.; Ofengand, J. A high sensitivity method for sequencing RNA: Application to ribosomal RNA. Biotechniques 1992, 13, 682–684. [Google Scholar]
- Bakin, A.; Ofengand, J. Four newly located pseudouridylate residues in Escherichia coli 23S ribosomal RNA are all at the peptidyltransferase center: Analysis by the application of a new sequencing technique. Biochemistry 1993, 32, 9754–9762. [Google Scholar] [CrossRef]
- Schaefer, M.; Kapoor, U.; Jantsch, M.F. Understanding RNA modifications: The promises and technological bottlenecks of the ‘epitranscriptome’. Open. Biol. 2017, 7, 170077. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Motorin, Y. Next-generation sequencing technologies for detection of modified nucleotides in RNAs. RNA Biol. 2017, 14, 1124–1137. [Google Scholar] [CrossRef]
- Helm, M.; Motorin, Y. Detecting RNA modifications in the epitranscriptome: Predict and validate. Nat. Rev. Genet. 2017, 18, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiong, X.; Yi, C. Epitranscriptome sequencing technologies: Decoding RNA modifications. Nat. Methods 2016, 14, 23–31. [Google Scholar] [CrossRef]
- Vandivier, L.E.; Anderson, Z.D.; Gregory, B.D. HAMR: High-Throughput Annotation of Modified Ribonucleotides. Methods Mol. Biol. 2019, 1870, 51–67. [Google Scholar] [PubMed] [Green Version]
- Vandivier, L.E.; Gregory, B.D. Reading the Epitranscriptome: New Techniques and Perspectives. Enzymes 2017, 41, 269–298. [Google Scholar] [PubMed]
- Motorin, Y.; Marchand, V. Detection and Analysis of RNA Ribose 2’-O-Methylations: Challenges and Solutions. Genes (Basel) 2018, 9, 642. [Google Scholar] [CrossRef] [Green Version]
- Incarnato, D.; Anselmi, F.; Morandi, E.; Neri, F.; Maldotti, M.; Rapelli, S.; Parlato, C.; Basile, G.; Oliviero, S. High-throughput single-base resolution mapping of RNA 2-O-methylated residues. Nucleic Acids Res. 2017, 45, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Pirnie, S.P.; Carmichael, G.G. High-throughput and site-specific identification of 2’-O-methylation sites using ribose oxidation sequencing (RibOxi-seq). RNA 2017, 23, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Moshitch-Moshkovitz, S.; Han, D.; Kol, N.; Amariglio, N.; Rechavi, G.; Dominissini, D.; He, C. Nm-seq maps 2’-O-methylation sites in human mRNA with base precision. Nat. Methods 2017, 14, 695–698. [Google Scholar] [CrossRef]
- Schwartz, S.; Bernstein, U.A.; Mumbach, M.; Jovanovic, M.; Herbst, R.H.; León-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouridylation of ncRNA and mRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovejoy, A.F.; Riordan, D.P.; Brown, P.O. Transcriptome-wide mapping of pseudouridines: Pseudouridine synthases modify specific mRNAs in S. cerevisiae. PLoS ONE 2014, 9, e110799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Marchand, V.; Motorin, Y.; Lafontaine, D.L.J. Identification of sites of 2’-O-methylation vulnerability in human ribosomal RNAs by systematic mapping. Sci. Rep. 2017, 7, 11490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, N.; Jansson, M.; Häfner, S.; Tehler, D.E.; Birkedal, U.; Christensen-Dalsgaard, M.; Lund, A.H.; Nielsen, H. Profiling of 2’-O-Me in human rRNA reveals a subset of fractionally modified positions and provides evidence for ribosome heterogeneity. Nucleic Acids Res. 2016, 44, 7884–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebras, J.; Krogh, N.; Marty, V.; Nielsen, H.; Cavaillé, J. Developmental changes of rRNA ribose methylations in the mouse. RNA Biol. 2020, 17, 150–164. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojha, S.; Malla, S.; Lyons, S.M. snoRNPs: Functions in Ribosome Biogenesis. Biomolecules 2020, 10, 783. https://doi.org/10.3390/biom10050783
Ojha S, Malla S, Lyons SM. snoRNPs: Functions in Ribosome Biogenesis. Biomolecules. 2020; 10(5):783. https://doi.org/10.3390/biom10050783
Chicago/Turabian StyleOjha, Sandeep, Sulochan Malla, and Shawn M. Lyons. 2020. "snoRNPs: Functions in Ribosome Biogenesis" Biomolecules 10, no. 5: 783. https://doi.org/10.3390/biom10050783
APA StyleOjha, S., Malla, S., & Lyons, S. M. (2020). snoRNPs: Functions in Ribosome Biogenesis. Biomolecules, 10(5), 783. https://doi.org/10.3390/biom10050783