Small Circular DNA Molecules as Triangular Scaffolds for the Growth of 3D Single Crystals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and DNA Strands
2.2. Preparation of Circular DNA
2.3. Crystallisation
2.4. TEM Imaging
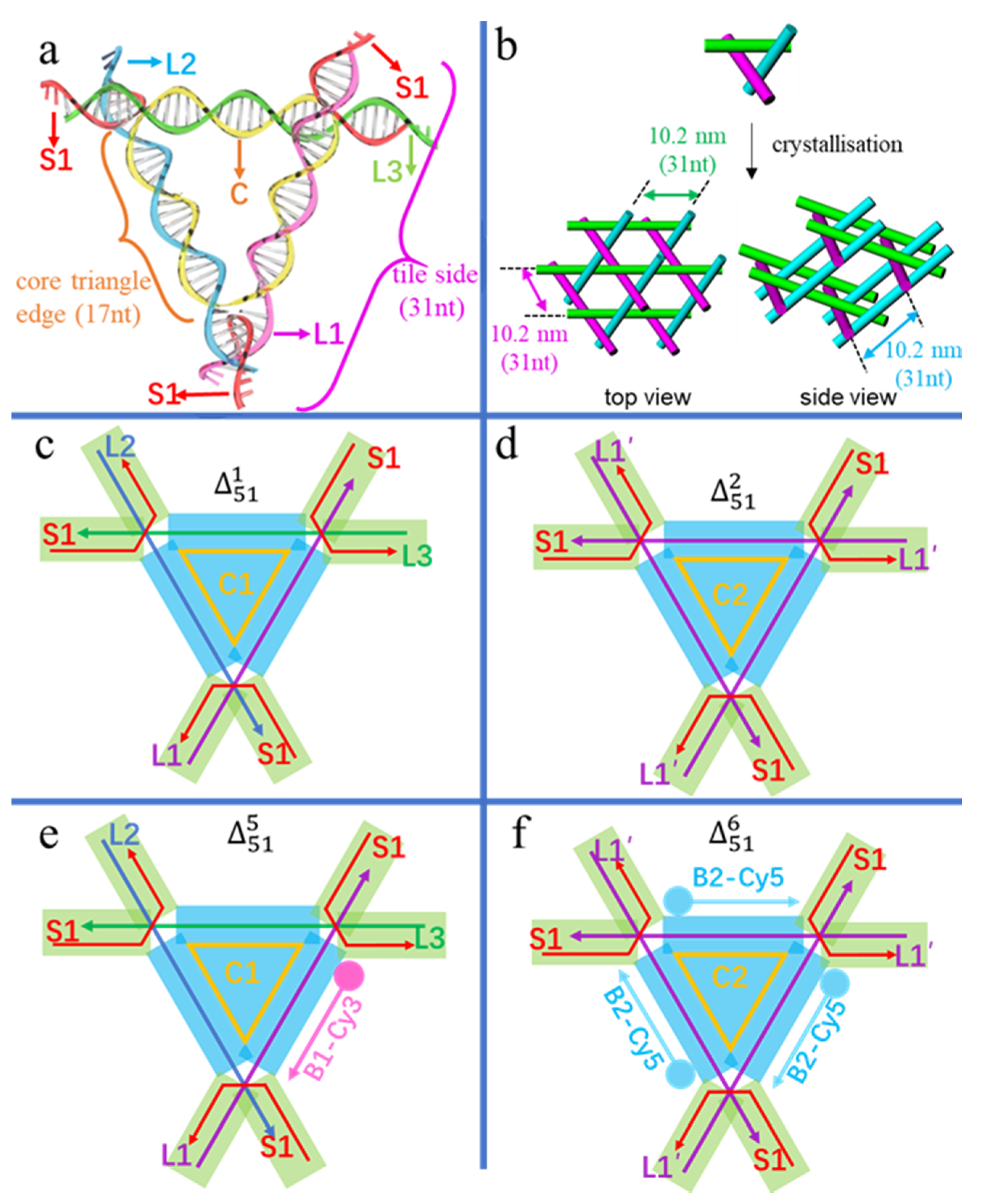
2.5. Overall Crystal Designs
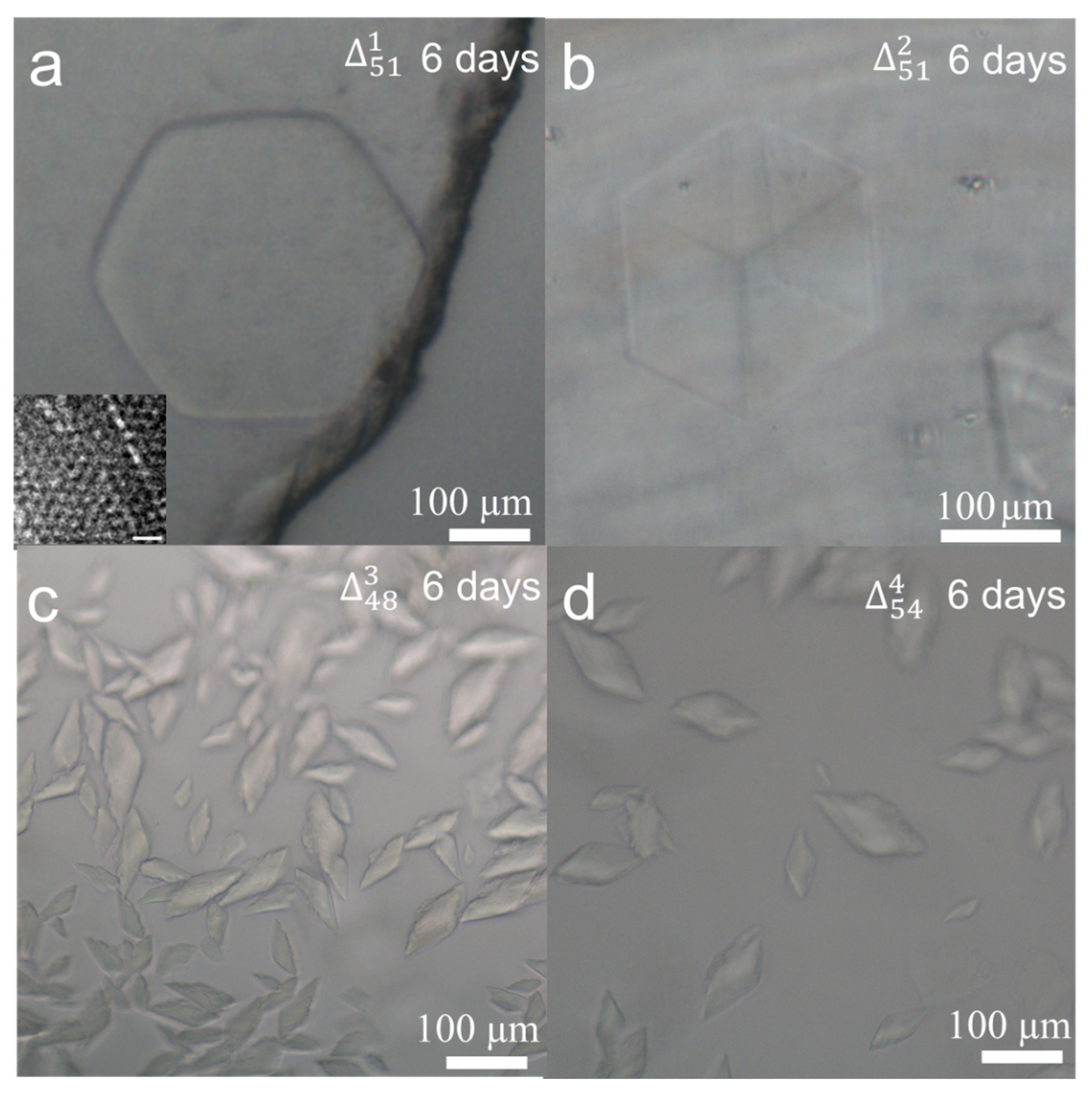
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wei, B.; Dai, M.; Yin, P. Complex shapes self-assembled from single-stranded DNA tiles. Nature 2012, 485, 623–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Li, Z.; Wang, P.; Meyer, T.; Mao, C.; Ke, Y. Reconfiguration of DNA molecular arrays driven by information relay. Science 2017, 357, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, S.M.; Dietz, H.; Liedl, T.; Hogberg, B.; Graf, F.; Shih, W.M. Self-assembly of DNA into nanoscale three-dimensional shapes. Nature 2009, 459, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Han, D.R.; Pal, S.; Yang, Y.; Jiang, S.X.; Nangreave, J.; Liu, Y.; Yan, H. DNA gridiron nanostructures based on four-arm junctions. Science 2013, 339, 1412–1415. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Pal, S.; Nangreave, J.; Deng, Z.; Liu, Y.; Yan, H. DNA origami with complex curvatures in three-dimensional space. Science 2011, 332, 342–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Jiang, S.; Wu, S.; Li, Y.; Mao, C.; Liu, Y.; Yan, H. Complex wireframe DNA origami nanostructures with multi-arm junction vertices. Nat. Nanotech. 2015, 10, 779–784. [Google Scholar] [CrossRef]
- Matthies, M.; Agarwal, N.P.; Schmidt, T.L. Design and synthesis of triangulated DNA origami trusses. Nano Lett. 2016, 16, 2108–2113. [Google Scholar] [CrossRef]
- Macfarlane, R.J.; Lee, B.; Jones, M.R.; Harris, N.; Schatz, G.C.; Mirkin, C.A. Nanoparticle superlattice engineering with DNA. Science 2011, 334, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.R.; Osberg, K.D.; Macfarlane, R.J.; Langille, M.R.; Mirkin, C.A. Templated techniques for the synthesis and assembly of plasmonic Nanostructures. Chem. Rev. 2011, 111, 3736–3827. [Google Scholar] [CrossRef]
- Stebe, K.J.; Lewandowski, E.; Ghosh, M. Oriented assembly of metamaterials. Science 2009, 325, 159–160. [Google Scholar] [CrossRef]
- Geng, C.; Paukstelis, P.J. DNA crystals as vehicles for biocatalysis. J. Am. Chem. Soc. 2014, 136, 7817–7820. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Chen, Z.G.; Shin, D.M. Nanoparticle therapeutics: An emerging treatment modality for cancer. Nat. Rev. Drug Discov. 2008, 7, 771–782. [Google Scholar] [CrossRef]
- Bell, A.T. The impact of nanoscience on heterogeneous catalysis. Science 2003, 299, 1688–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.F.; Yu, L.; Shen, W.Q. DNA Nanotechnology and its applications in biomedical research. J. Biomed. Nanotechnol. 2014, 10, 2350–2370. [Google Scholar] [CrossRef]
- Zheng, J.; Birktoft, J.J.; Chen, Y.; Wang, T.; Sha, R.; Constantinou, P.E.; Ginell, S.L.; Mao, C.; Seeman, N.C. From molecular to macroscopic via the rational design of a self-assembled 3D DNA crystal. Nature 2009, 461, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Rusling, D.A.; Chandrasekaran, A.R.; Ohayon, Y.P.; Brown, T.; Fox, K.R.; Sha, R.; Mao, C.; Seeman, N.C. Functionalizing designer DNA crystals with a triple-helical veneer. Angew. Chem. Int. Ed. Engl. 2014, 53, 3979–3982. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.; Birktoft, J.J.; Sha, R.J.; Wang, T.; Zheng, J.P.; Constantinou, P.E.; Ginell, S.L.; Chen, Y.; Mao, C.D.; Seeman, N.C. The absence of tertiary interactions in a self-assembled DNA crystal structure. J. Mol. Recognit. 2012, 25, 234–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durland, R.H.; Kessler, D.J.; Gunnell, S.; Duvic, M.; Pettitt, B.M.; Hogan, M.E. Binding of triple helix forming oligonucleotides to sites in gene promoters. Biochemistry 1991, 30, 9246–9255. [Google Scholar] [CrossRef]
- Li, N.; Wang, J.; Ma, K.; Liang, L.; Mi, L.; Huang, W.; Ma, X.; Wang, Z.; Zheng, W.; Xu, L.; et al. The dynamics of forming a triplex in an artificial telomere inferred by DNA mechanics. Nucleic Acids Res. 2019, 47, e86. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.K.; Adaku, N.; Peart, N.; Rogers, F.A. Triplex structures induce DNA double strand breaks via replication fork collapse in NER deficient cells. Nucleic Acids Res. 2016, 44, 7742–7754. [Google Scholar]
- Huo, S.D.; Gong, N.Q.; Jiang, Y.; Chen, F.; Guo, H.B.; Gan, Y.L.; Wang, Z.S.; Herrmann, A.; Liang, X.J. Gold-DNA nanosunflowers for efficient gene silencing with controllable transformation. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.M.; Chandrasekaran, A.R.; Li, Q.; Li, X.; Sha, R.J.; Seeman, N.C.; Mao, C.D. Post-assembly stabilization of rationally designed DNA crystals. Angew. Chem. Int. Ed. Engl. 2015, 54, 9936–9939. [Google Scholar] [CrossRef]
- Abdallah, H.O.; Ohayon, Y.P.; Chandrasekaran, A.R.; Sha, R.J.; Fox, K.R.; Brown, T.; Rusling, D.A.; Mao, C.; Seeman, N.C. Stabilisation of self-assembled DNA crystals by triplex-directed photo-cross-linking. Chem. Commun. 2016, 52, 8014–8017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, E.; Praetorius, F.; de Oliveira Mann, C.C.; Hopfner, K.P.; Dietz, H. Impact of heterogeneity and lattice bond strength on DNA triangle crystal growth. ACS Nano 2016, 10, 9156–9164. [Google Scholar] [CrossRef] [PubMed]
- Ohayon, Y.P.; Hernandez, C.; Chandrasekaran, A.R.; Wang, X.Y.; Abdallah, H.O.; Jong, M.A.; Mohsen, M.G.; Sha, R.J.; Birktoft, J.J.; Lukeman, P.S.; et al. Designing higher resolution self-assembled 3D DNA crystals via strand terminus modifications. ACS Nano 2019, 13, 7957–7965. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.M.; Zhao, Y.; Li, Z.; Wang, Y.; Sha, R.J.; Seeman, N.C.; Mao, C.D. Modulating self-assembly of DNA crystals with rationally designed agents. Angew. Chem. Int. Ed. Engl. 2018, 57, 16529–16532. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.F.; Zheng, M.X.; Zhao, J.M.; Seeman, N.C.; Mao, C.D. Making engineered 3D DNA crystals robust. J. Am. Chem. Soc. 2019, 141, 15850–15855. [Google Scholar] [CrossRef]
- Zheng, H.; Xiao, M.; Yan, Q.; Ma, Y.; Xiao, S.J. Small circular DNA molecules act as rigid motifs to build DNA nanotubes. J. Am. Chem. Soc. 2014, 136, 10194–10197. [Google Scholar] [CrossRef]
- Wang, M.; Guo, X.; Jiang, C.; Wang, X.; Xiao, S.-J. In-phase assembly of slim DNA lattices with small circular DNA motifs via short connections of 11 and 16 base pairs. ChemBioChem 2016, 17, 1132–1137. [Google Scholar] [CrossRef]
- Wang, M.; Huang, H.; Zhang, Z.; Xiao, S.J. 2D DNA lattices constructed from two-tile DAE-O systems possessing circular central strands. Nanoscale 2016, 8, 18870–18875. [Google Scholar] [CrossRef]
- Afshan, N.; Ali, M.; Wang, M.; Baig, M.; Xiao, S.J. DNA nanotubes assembled from tensegrity triangle tiles with circular DNA scaffolds. Nanoscale 2017, 9, 17181–17185. [Google Scholar] [CrossRef]
- Giannoni, G.; Padden, F.; Keith, H.D. Crystallization of DNA from dilute solution. Proc. Natl. Acad. Sci. USA 1968, 62, 964–971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Szostak, J.W.; Huang, Z. Nucleic acid crystallization and X-ray crystallography facilitated by single selenium atom. Front. Chem. Sci. Eng. 2016, 10, 196–202. [Google Scholar] [CrossRef]
- Rusling, D.A.; Powers, V.E.; Ranasinghe, R.T.; Wang, Y.; Osborne, S.D.; Brown, T.; Fox, K.R. Four base recognition by triplex-forming oligonucleotides at physiological pH. Nucleic Acids Res. 2005, 33, 3025–3032. [Google Scholar] [CrossRef] [Green Version]
- Foltman, B. On the crystallisation, stability and proteolytic activity of rennin. Acta Chem. Scand. 1959, 13, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Chayen, N.E. A novel technique to control the rate of vapour diffusion, giving larger protein crystals. J. Appl. Cryst. 1997, 30, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Kristiansen, M.; Sha, R.; Birktoft, J.J.; Hernandez, C.; Mao, C.; Seeman, N.C. A device that operates within a self-assembled 3D DNA crystal. Nat. Chem. 2017, 9, 824–827. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tile Components | C1, L1, L2, L3, 3 × S1 | C2, 3×L1′, 3 × S1 | C3, 3 × L4, 3 × S1 | C4, 3 × L5, 3 × S1 | C1, L1, L2, L3, 3 × S1, B1-Cy3 | C2, 3 × L1′, 3 × S1, 3 × (B2-Cy5) |
| Crystal Shapes and Estimated Sizes | Hexagonal prism, a(= b) = 100–450 µm, c = 20–40 µm | Cube, a(= b = c) = 100–150 µm | Oblique triangular prism, a(= b) = 20~30 µm, c = 10~30 µm | Oblique triangular prism, a(= b) = 30~50 µm, c = 10~30 µm | Hexagonal prism, a(= b) = 100~150 µm, c = 10~20 µm | Oblique triangular prism, a(= b) = 60~110 µm, c = 40~60 µm |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Guo, X.; Kou, B.; Zhang, L.; Xiao, S.-J. Small Circular DNA Molecules as Triangular Scaffolds for the Growth of 3D Single Crystals. Biomolecules 2020, 10, 814. https://doi.org/10.3390/biom10060814
Wang Y, Guo X, Kou B, Zhang L, Xiao S-J. Small Circular DNA Molecules as Triangular Scaffolds for the Growth of 3D Single Crystals. Biomolecules. 2020; 10(6):814. https://doi.org/10.3390/biom10060814
Chicago/Turabian StyleWang, Yu, Xin Guo, Bo Kou, Ling Zhang, and Shou-Jun Xiao. 2020. "Small Circular DNA Molecules as Triangular Scaffolds for the Growth of 3D Single Crystals" Biomolecules 10, no. 6: 814. https://doi.org/10.3390/biom10060814
APA StyleWang, Y., Guo, X., Kou, B., Zhang, L., & Xiao, S. -J. (2020). Small Circular DNA Molecules as Triangular Scaffolds for the Growth of 3D Single Crystals. Biomolecules, 10(6), 814. https://doi.org/10.3390/biom10060814