Is Memantine Effective as an NMDA Receptor Antagonist in Adjunctive Therapy for Schizophrenia?

Abstract
:1. Introduction
2. NMDA Receptor Hypofunction in Schizophrenia
3. Memantine as Adjunctive Therapy in Schizophrenia
4. Why is Memantine Free of the Severe Side Effects of NMDA Receptor Antagonists, such as PCP and Ketamine?
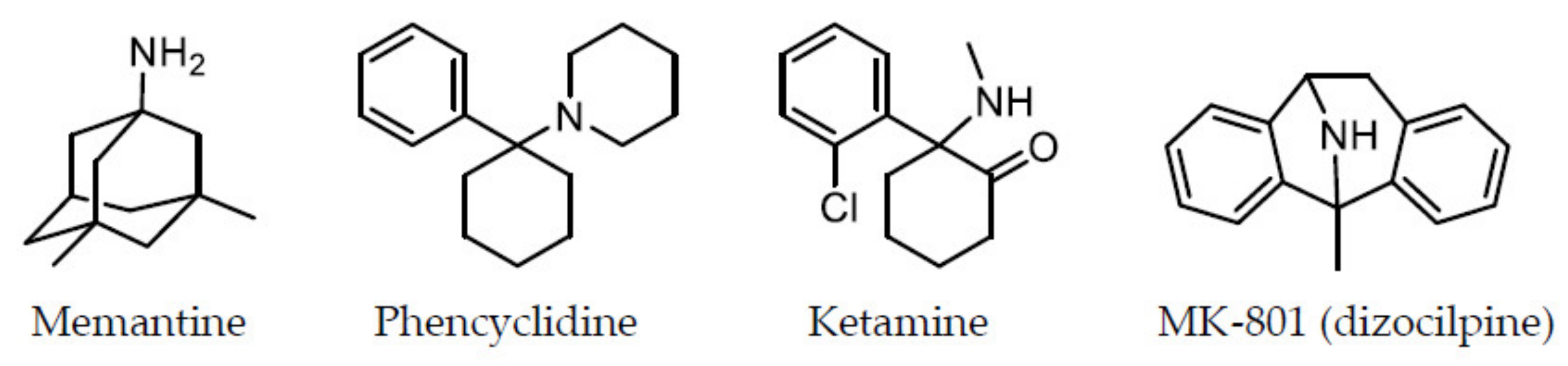
4.1. Affinity for the PCP-Binding Site
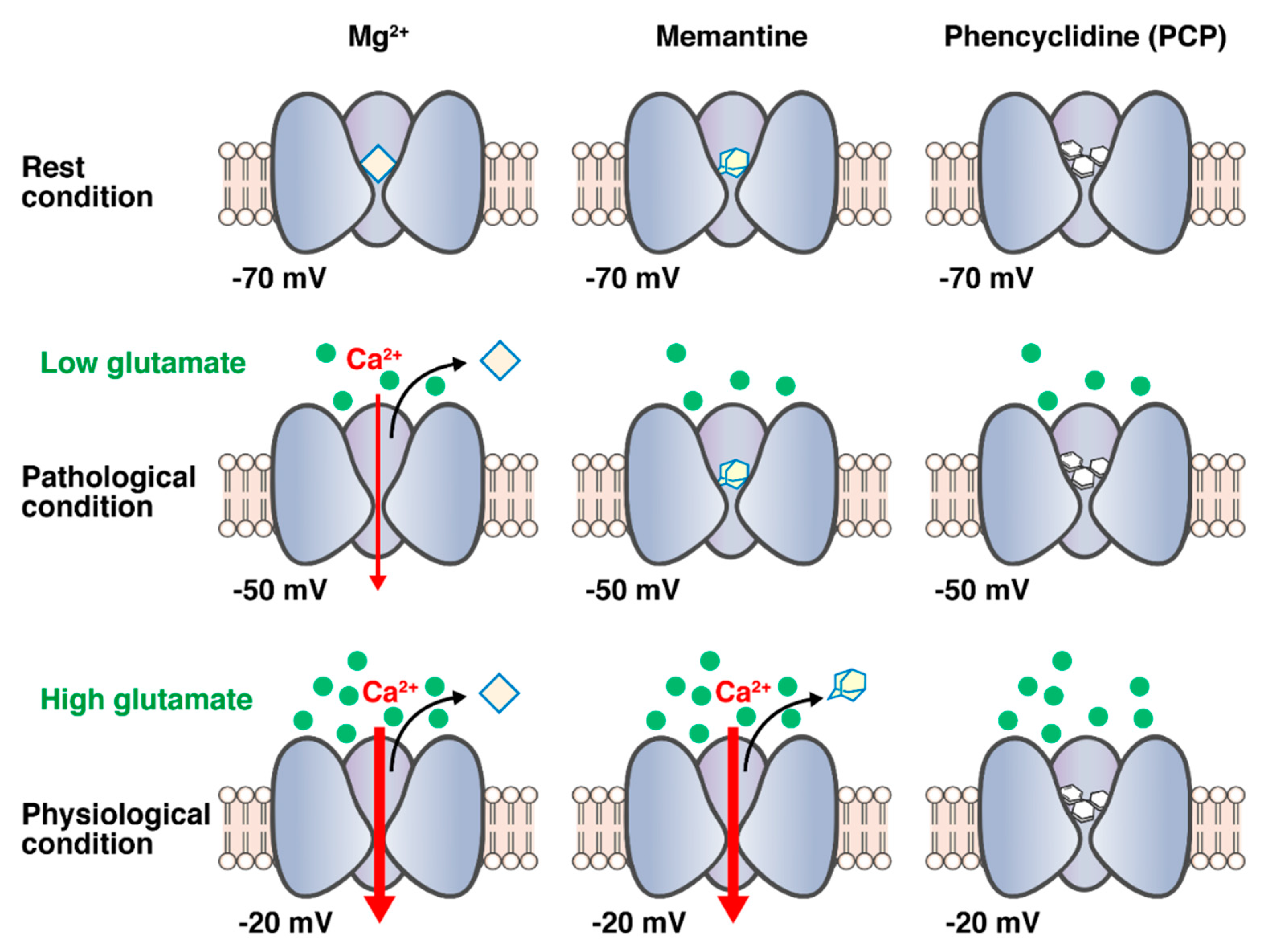
4.2. Mode of Fast Off-Rate
4.3. Voltage Dependency
4.4. Uncompetitive Antagonist
4.5. Preferential Inhibition of Extrasynaptic Receptors
5. Can Memantine Ameliorate Abnormalities Caused by Other NMDA Receptor Antagonists or Caused by Other Manipulations?
6. Can the NMDA Receptor Antagonist Activity of Memantine Be Exerted at Clinical Therapeutic Concentrations?
7. Conclusions
Funding
Conflicts of Interest
References
- Conley, R.R.; Kelly, D.L. Current status of antipsychotic treatment. Curr. Drug Targets CNS Neurol. Disord. 2002, 37, 123–128. [Google Scholar] [CrossRef]
- Huhn, M.; Nikolakopoulou, A.; Schneider-Thoma, J.; Krause, M.; Samara, M.; Peter, N.; Arndt, T.; Bäckers, L.; Rothe, P.; Cipriani, A.; et al. Comparative efficacy and tolerability of 32 oral antipsychotics for the acute treatment of adults with multi-episode schizophrenia: A systematic review and network meta-analysis. Lancet 2019, 394, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.C.; Tsai, S.-J. New target for schizophrenia treatment beyond the dopamine hypothesis. Int. J. Mol. Sci. 2017, 18, 1689. [Google Scholar] [CrossRef]
- Atri, A. The Alzheimer’s disease clinical spectrum: Diagnosis and management. Med. Clin. N. Am. 2019, 103, 263–293. [Google Scholar] [CrossRef]
- Balu, D.T. The NMDA receptor and schizophrenia: From pathophysiology to treatment. Adv. Pharmacol. 2016, 76, 351–382. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Coyle, J.T. Glutamate hypothesis in schizophrenia. Psychiatry Clin. Neurosci. 2019, 73, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Javitt, D.C.; Zukin, S.R. Recent advances in the phencyclidine model of schizophrenia. Am. J. Psychiatry 1991, 148, 1301–1308. [Google Scholar] [CrossRef]
- Kristal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B.; Charney, D.S. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Arch. Gen. Psychiatry 1994, 51, 199–214. [Google Scholar] [CrossRef]
- Lahti, A.C.; Holcomb, H.H.; Medoff, D.R.; Tamminga, C.A. Ketamine activates psychosis and alters limbic blood flow in schizophrenia. Neuroreport 1995, 6, 869–872. [Google Scholar] [CrossRef]
- Malhotra, A.K.; Pinals, D.A.; Adler, C.M.; Elman, I.; Clifton, A.; Pickar, D.; Breier, A. Ketamine-induced exacerbation of psychotic symptoms and cognitive impairment in neuroleptic-free schizophrenia. Neuropsychopharmacology 1997, 17, 141–150. [Google Scholar] [CrossRef]
- Wong, E.H.F.; Kemp, J.A.; Priestley, T.; Knight, A.R.; Woodruff, G.N. The anticonvulsant MK-801 is a potent N-methyl-D-aspartate antagonist. Proc. Natl. Acad. Sci. USA 1986, 83, 7104–7108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domino, E.F. Chemical dissociation of human awareness: Focus on non-competitive NMDA receptor antagonists. J. Psychopharmacol. 1992, 6, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Coyle, J.T. The glutamatergic dysfunction hypothesis for schizophrenia. Harv. Rev. Psychiatry 1996, 3, 241–253. [Google Scholar] [CrossRef]
- Snyder, M.A.; Gao, W.-J. NMDA receptor hypofunction for schizophrenia revisited: Perspectives from epigenetic mechanisms. Schizophr. Res. 2020, 217, 60–70. [Google Scholar] [CrossRef]
- Nakazawa, K.; Sapkota, K. The origin of NMDA receptor hypofunction in schizophrenia. Pharmacol. Ther. 2020, 205, 107426. [Google Scholar] [CrossRef]
- Hu, W.; MacDonald, M.L.; Elswick, D.E.; Sweet, R.A. The glutamate hypothesis of schizophrenia: Evidence from human brain tissue studies. Ann. N. Y. Acad. Sci. 2015, 1338, 38–57. [Google Scholar] [CrossRef] [Green Version]
- Pilowsky, L.S.; Bressan, R.A.; Stone, J.M.; Erlandsson, K.; Mulligan, R.S.; Krystal, J.H.; Ell, P.J. First in vivo evidence of an NMDA receptor deficit in medication-free schizophrenic patients. Mol. Psychiatry 2006, 11, 118–119. [Google Scholar] [CrossRef] [Green Version]
- Poels, E.M.P.; Kegeles, L.S.; Kantrowitz, J.T.; Slifstein, M.; Javitt, D.C.; Lieberman, J.A.; Abi-Dargham, A.; Girgis, R.R. Imaging glutamate in schizophrenia: Review of findings and implications for drug discovery. Mol. Psychiatry 2014, 19, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.J.; Owen, M.J. Genes for schizophrenia? Recent findings and their pathophysiological implications. Lancet 2003, 361, 417–419. [Google Scholar] [CrossRef]
- Schwartz, T.L.; Sachdeva, S.; Stahl, S.M. Genetic data supporting the NMDA glutamate receptor hypothesis for schizophrenia. Curr. Pharm. Des. 2012, 18, 1580–1592. [Google Scholar] [CrossRef]
- Kayser, M.S.; Dalmau, J. Anti-NMDA receptor encephalitis, autoimmunity, and psychosis. Schizophr. Res. 2016, 176, 36–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellaithy, A.; Younkin, J.; González-Maeso, J.; Logothetis, D.E. Positive allosteric modulators of metabotropic glutamate 2 receptors in schizophrenia treatment. Trends Neurosci. 2015, 38, 506–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, K. Targeting of NMDA receptors in new treatments for schizophrenia. Expert Opin. Ther. Targets 2014, 18, 1049–1063. [Google Scholar] [CrossRef]
- Kishi, T.; Iwata, N. NMDA receptor antagonists interventions in schizophrenia: Meta-analysis of randomized, placebo-controlled trials. J. Psychiatr. Res. 2013, 47, 1143–1149. [Google Scholar] [CrossRef]
- Rezaei, F.; Mohammad-karimi, M.; Seddighi, S.; Modabbernia, A.; Ashrafi, M.; Salehi, B.; Hammidi, S.; Motasami, H.; Hajiaghaee, R.; Tabrizi, M.; et al. Memantine add-on to risperidone for treatment of negative symptoms in patients with stable schizophrenia: Randomized, double-blind, placebo-controlled study. J. Clin. Psychopharmacol. 2013, 33, 336–342. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kishi, T.; Iwata, N. Efficacy and safety of NMDA receptor antagonists augmentation therapy for schizophrenia: An updated meta-analysis of randomized placebo-controlled trials. J. Psychiatr. Res. 2013, 47, 2018–2020. [Google Scholar] [CrossRef]
- Kishi, T.; Matsuda, Y.; Iwata, N. Memantine add-on to antipsychotic treatment for residual negative and cognitive symptoms of schizophrenia: A meta-analysis. Psychopharmacology 2017, 234, 2113–2125. [Google Scholar] [CrossRef]
- Di Iorio, G.; Baroni, G.; Lorusso, M.; Montemitro, C.; Spano, M.C.; di Giannantonio, M. Efficacy of memantine in schizophrenic patients: A systematic review. J. Amino Acids 2017, 7021071. [Google Scholar] [CrossRef] [Green Version]
- Fakhri, A.; Pakseresht, S.; Haghdoost, M.R.; Hekmatkhah, N.; Torkashvand, M.; Ghorbanzadeh, B. Memantine enhances the effect of olanzapine in patients with schizophrenia: A randomized, placebo-controlled study. Acta Med. Iran. 2016, 54, 696–703. [Google Scholar] [PubMed]
- Mazinani, R.; Nejati, S.; Khodaei, M. Effects of memantine added to risperidone on the symptoms of schizophrenia: A randomized double-blind, placebo-controlled clinical trial. Psychiatry Res. 2017, 247, 291–295. [Google Scholar] [CrossRef]
- Omranifard, V.; Rajabi, F.; Mohammadian-Sichani, M.; Maracy, M.R. The effect of add-on memantine on positive, negative and depressive symptoms of schizophrenia: A double-blind, randomized, controlled trial. Actas Esp. Psiquiatr. 2017, 45, 107–115. [Google Scholar] [PubMed]
- Hassanpour, F.; Zarghami, M.; Mouodi, S.; Moosazadeh, M.; Barzegar, F.; Bagheri, M.; Hendouei, N. Adjunctive memantine treatment of schizophrenia: A double-blind, randomized placebo-controlled study. J Clin. Psychopharmacol. 2019, 39, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Li, X.-H.; Yang, X.-H.; Cai, D.-B.; Ungvari, G.S.; Ng, C.H.; Wang, S.-B.; Wang, Y.-Y.; Ning, Y.-P.; Xiang, Y.-T. Adjunctive memantine for schizophrenia: A meta-analysis of randomized, double-blind, placebo-controlled trials. Psychol. Med. 2018, 48, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhu, X.-M.; Zhang, Q.-E.; Cai, D.-B.; Yang, X.-H.; Zhou, Y.-L.; Ungvari, G.S.; Ng, C.H.; He, S.-H.; Peng, X.-J.; et al. Adjunctive memantine for major mental disorders; A systematic review and meta-analysis of randomized double-blind controlled trials. Schizophr. Res. 2019, 209, 12–21. [Google Scholar] [CrossRef]
- Schaefer, M.; Sarkar, S.; Theophil, I.; Leopold, K.; Heinz, A.; Gallinat, J. Acute and long-term memantine add-on treatment to risperidone improves cognitive dysfunction in patients with acute and chronic schizophrenia. Pharmacopsychiatry 2020, 53, 21–29. [Google Scholar] [CrossRef]
- Gray, L.; van den Buuse, M.; Scarr, E.; Dean, B.; Hannan, A.J. Clozapine reverses schizophrenia-related behaviours in the metabotropic glutamate receptor 5 knockout mouse: Association with N-methyl-D-aspartic acid receptor up-regulation. Int. J. Neuropsychopharmacol. 2009, 12, 45–60. [Google Scholar] [CrossRef] [Green Version]
- de Bartolomeis, A.; Sarappa, C.; Magara, S.; Iasevoli, F. Targeting glutamate system for novel antipsychotic approaches: Relevance for residual psychotic symptoms and treatment resistant schizophrenia. Eur. J. Pharmacol. 2012, 682, 1–11. [Google Scholar] [CrossRef]
- de Lucena, D.; Fernandes, B.S.; Berk, M.; Dodd, S.; Medeiros, D.W.; Pedrini, M.; Kunz, M.; Gomes, F.A.; Giglio, L.F.; Lobato, M.I.; et al. Improvement of negative and positive symptoms in treatment-refractory schizophrenia: A double-blind, randomized, placebo-controlled trial with memantine as add-on therapy to clozapine. J. Clin. Psychiatry 2009, 70, 1416–1423. [Google Scholar] [CrossRef] [Green Version]
- Veerman, S.R.T.; Schulte, P.F.J.; Smith, J.D.; de Haan, L. Memantine augmentation in clozapine-refractory schizophrenia: A randomized, double-blind, placebo-controlled crossover study. Psychol. Med. 2016, 46, 1909–1921. [Google Scholar] [CrossRef] [Green Version]
- Veerman, S.R.T.; Schulte, P.F.J.; Deijen, J.B.; de Haan, L. Adjunctive memantine in clozapine-treated refractory schizophrenia: An open-label 1-year extension study. Psychol. Med. 2017, 47, 363–375. [Google Scholar] [CrossRef]
- Omranifard, V.; Rajabi, F.; Mohammadian-Sichani, M.; Maracy, M. The effect of add-on memantine on global function and quality of life in schizophrenia: A randomized, double-blind, controlled, clinical trial. Adv. Biomed. Res. 2015, 4, 211. [Google Scholar] [PubMed]
- Merritt, K.; Egerton, A.; Kempton, M.J.; Taylor, M.J.; McGuire, P.K. Nature of glutamate alterations in schizophrenia: A meta-analysis of proton magnetic resonance spectroscopy studies. JAMA Psychiatry 2016, 73, 665–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sani, G.; Serra, G.; Kotzalidis, G.D.; Romano, S.; Tamorri, S.M.; Manfredi, G.; Caloro, M.; Telesforo, C.L.; Caltagirone, S.S.; Panaccione, I.; et al. The role of memantine in the treatment of psychiatric disorders other than the dementias: A review of current preclinical and clinical evidence. CNS Drugs 2012, 26, 663–690. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Nasrallah, H.A. The use of memantine in neuropsychiatric disorders: An overview. Ann. Clin. Psychiatry 2018, 30, 234–248. [Google Scholar] [PubMed]
- Temme, L.; Schepmann, D.; Schreiber, J.A.; Frehland, B.; Wünsch, B. Comparative pharmacological study of common NMDA receptor open channel blockers regarding their affinity and functional activity toward GluN2A and GluN2B NMDA receptors. ChemMedChem 2018, 13, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Sax, M.; Wünsch, B. Relationship between the structure of dexoxadrol and etoxadrol analogues and their NMDA receptor affinity. Curr. Top. Med. Chem. 2006, 6, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, F.X.; Leenders, K.L.; Øye, I.; Hell, D.; Angst, J. Differential psychopathology and patterns of cerebral glucose utilisation produced by (S)- and (R)-ketamine in healthy volunteers using positron emission tomography (PET). Eur. Neuropsychopharmacol. 1997, 7, 25–38. [Google Scholar] [CrossRef]
- Angus, S.; Sugars, J.; Boltezar, R.; Koskewich, S.; Schneider, N.M. A controlled trial of amantadine hydrochloride and neuroleptics in the treatment of tardive dyskinesia. J. Clin. Psychopharmacol. 1997, 17, 88–91. [Google Scholar] [CrossRef]
- Nguyen, L.; Thomas, K.L.; Lucke-Wold, B.P.; Cavendish, J.Z.; Crowe, M.S.; Matsumoto, R.R. Dextromethorphan: An update on its utility for neurological and neuropsychiatric disorders. Pharmacol. Therapeut. 2016, 159, 1–22. [Google Scholar] [CrossRef]
- Parsons, C.G.; Danysz, W.; Quack, G. Memantine is a clinically well tolerated N-methyl-D-aspartate (NMDA) receptor antagonist—A review of preclinical data. Neuropharmacology 1999, 38, 735–767. [Google Scholar] [CrossRef]
- Parsons, C.G.; Danysz, W.; Quack, G. Memantine and the amino-alkyl-cyclohexane MRZ 2/579 are moderate affinity uncompetitive NMDA receptor antagonists—In vitro characterization. Amino Acids 2000, 19, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Heusler, P.; Tourette, A.; Cussac, D. Potencies and unblocking kinetic properties of antagonists at recombinant human NMDA receptors in a Xenopus oocytes model. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Lipton, S.A. Paradigm shift in neuroprotection by NMDA receptor blockade: Memantine and beyond. Nat. Rev. Drug Discov. 2006, 5, 160–170. [Google Scholar] [CrossRef]
- Parsons, C.G.; Stöffler, A.; Danysz, W. Memantine: A NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system—Too little activation is bad, too much is even worse. Neuropharmacology 2007, 53, 699–723. [Google Scholar] [CrossRef]
- Rammes, G.; Danysz, W.; Parsons, C.G. Pharmacodynamics of memantine: An update. Curr. Neuropharmacol. 2008, 6, 55–78. [Google Scholar] [CrossRef] [Green Version]
- Blanpied, T.A.; Boeckman, F.A.; Aizenman, E.; Johnson, J.W. Tapping Channel block of NMDA-activated responses by amantadine and memantine. J. Neurophysiol. 1997, 77, 309–323. [Google Scholar] [CrossRef] [Green Version]
- Lipton, S.A. Paradigm shift in NMDA receptor antagonist drug development: Molecular mechanism of uncompetitive inhibition by memantine in the treatment of Alzheimer’s disease and other neurologic disorders. J. Alzheimers Dis. 2004, 6, S61–S74. [Google Scholar] [CrossRef]
- Chen, H.-S.V.; Pellegrini, J.W.; Aggarwal, S.K.; Lei, S.Z.; Warach, S.; Jensen, F.E.; Lipton, S.A. Open-channel block of N-methyl-D-aspartate (NMDA) responses by memantine: Therapeutic advantage against NMDA receptor-mediated neurotoxicity. J. Neurosci. 1992, 12, 4427–4436. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-S.V.; Lipton, S.A. Mechanism of memantine block of NMDA-activated channels in rat retinal ganglion cells: Uncompetitive antagonism. J. Physiol. 1997, 499, 27–46. [Google Scholar] [CrossRef]
- Gilling, K.E.; Jatzke, C.; Hechenberger, M.; Parsons, C.G. Potency, voltage-dependency, agonist concentration-dependency, blocking kinetics and partial untrapping of the uncompetitive N-methyl-D-aspartate (NMDA) channel blocker memantine at human NMDA (GluN1/GluN2A) receptors. Neuropharmacology 2009, 56, 866–875. [Google Scholar] [CrossRef]
- Rogawski, M.A.; Yamaguchi, S.; Jones, S.M.; Rice, K.C.; Thurkauf, A.; Monn, J.A. Anticonvulsant activity of the low-affinity uncompetitive N-methyl-D-aspartate antagonist (±)-5-aminocarbonyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine (ADCI): Comparison with the structural analogs dizocilpine (MK-801) and carbamazepine. J. Pharmacol. Exp. Ther. 1991, 259, 30–37. [Google Scholar] [PubMed]
- Seidleck, B.K.; Thurkauf, A.; Witkin, J.M. Evaluation of ADCI against convulsant and locomotor stimulant effects of cocaine: Comparison with the structural analogs dizocilpine and carbamazepine. Pharmacol. Biochem. Behav. 1994, 47, 839–844. [Google Scholar] [CrossRef]
- Song, X.; Jensen, M.Ø.; Jogini, V.; Stein, R.A.; Lee, C.-H.; Mchaourab, H.S.; Shaw, D.E.; Gouaux, E. Mechanism of NMDA receptor channel block by MK-801 and memantine. Nature 2018, 566, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Groc, L.; Bard, L.; Choquet, D. Surface trafficking of N-methyl-D-aspartate receptors: Physiological and pathological perspectives. Neuroscience 2009, 158, 4–18. [Google Scholar] [CrossRef]
- Xia, P.; Chen, H.-S.V.; Zhang, D.; Lipton, S.A. Memantine preferentially blocks extrasynaptic over synaptic NMDA receptor currents in hippocampal autapses. J. Neurosci. 2010, 30, 11246–11250. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-N.; Johnson, S.W. Memantine selectively blocks extrasynaptic NMDA receptors in rat substantia nigra dopamine neurons. Brain Res. 2015, 1603, 1–7. [Google Scholar] [CrossRef]
- Kaufman, A.M.; Milnerwood, A.J.; Sepers, M.D.; Coquinco, A.; She, K.; Wang, L.; Lee, H.; Craig, A.M.; Cynader, M.; Raymond, L.A. Opposing roles of synaptic and extrasynaptic NMDA receptor signaling in cocultured striatal and cortical neurons. J. Neurosci. 2012, 32, 3992–4003. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, S.; Pouladi, M.A.; Talantova, M.; Yao, D.; Xia, P.; Ehrnhoefer, D.E.; Zaidi, R.; Clemente, A.; Kaul, M.; Graham, R.K.; et al. Balance between synaptic versus extrasynaptic NMDA receptor activity influences inclusions and neurotoxicity of mutant huntingtin. Nat. Med. 2009, 15, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Savchenko, A.; Braun, G.B.; Molokanova, E. Nanostructured antagonist of extrasynaptic NMDA receptors. Nano Lett. 2016, 16, 5495–5502. [Google Scholar] [CrossRef]
- Johnson, J.W.; Glasgow, N.G.; Povysheva, N.V. Recent insights into the mode of action of memantine and ketamine. Curr. Opin. Pharmacol. 2015, 20, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Acker, T.M.; Yuan, H.; Hansen, K.B.; Vance, K.M.; Ogden, K.K.; Jensen, H.S.; Burger, P.B.; Mullasseril, P.; Snyder, J.P.; Liotta, D.C.; et al. Mechanism for noncompetitive inhibition by novel GluN2C/D N-methyl-D-aspartate receptor subunit-selective modulators. Mol. Pharmacol. 2011, 80, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Uribe, E.; Landaeta, J.; Wix, R.; Eblen, A. Memantine reverses social withdrawal induced by ketamine in rats. Exp. Neurobiol. 2013, 22, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Kakefuda, K.; Ishisaka, M.; Tsuruma, K.; Shimazawa, M.; Hara, H. Memantine, an NMDA receptor antagonist, improves working memory deficits in DGKβ knockout mice. Neurosci. Lett. 2016, 630, 228–232. [Google Scholar] [CrossRef]
- Owczarek, S.; Hou, J.; Secher, T.; Kristiansen, L.V. Phencyclidine treatment increases NR2A and NR2B N-methyl-D-aspartate receptor subunit expression in rats. NeuroReport 2011, 22, 935–938. [Google Scholar] [CrossRef]
- Fukuyama, K.; Hasegawa, T.; Okada, M. Cystine/glutamate antiporter and aripiprazole compensate NMDA antagonist-induced dysfunction of thalamocortical L-glutamatergic transmission. Int. J. Mol. Sci. 2018, 19, 3645. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Fukuyama, K.; Kawano, Y.; Shiroyama, T.; Ueda, Y. Memantine protects thalamocortical hyper-glutamatergic transmission induced by NMDA receptor antagonism via activation of system xc−. Pharmacol. Res. Perspect. 2019, 7, e00457. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.S.; Hatchett, A.D.; Fuller, M.A. Catatonic schizophrenia and the use of memantine. Ann. Pharmacother. 2006, 40, 344–346. [Google Scholar] [CrossRef]
- Obregon, D.F.; Velasco, R.M.; Wuerz, T.P.; Catalano, M.C.; Catalano, G.; Kahn, D. Memantine and Catatonia: A case report and literature review. J. Psychiatr. Pract. 2011, 17, 292–299. [Google Scholar] [CrossRef]
- Roy, K.; Warnick, S.J.; Balon, R. Catatonia delirium: 3 cases treated with memantine. Psychosomatics 2016, 57, 645–650. [Google Scholar] [CrossRef]
- Inta, D.; Sartorius, A.; Gass, P. NMDA receptor blockade and catatonia: A complex relationship. Schizophr. Res. 2015, 168, 581–582. [Google Scholar] [CrossRef]
- Ramirez-Bermudez, J.; Restrepo-Martinez, M.; Diaz-Victoria, A.R.; Espinola-Nadurille, M. Memantine as adjunctive therapy in a patient with anti-N-methyl-D-aspartate receptor encephalitis. J. Clin. Psychopharmacol. 2020, 40, 92–93. [Google Scholar] [CrossRef] [PubMed]
- Harms, L. Mismatch responses and deviance detection in N-methyl-D-aspartate (NMDA) receptor hypofunction and development models of schizophrenia. Biol. Psychiatry 2016, 116, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Bhakta, S.; Chou, H.-H.; Talledo, J.A.; Balvaneda, B.; Light, G.A. Memantine effects on sensorimotor gating and mismatch negativity in patients with chronic psychosis. Neuropsychopharmacology 2016, 41, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, L.; Parras, G.G.; Michie, P.T.; Malmierca, M.S. The role of glutamate neurotransmission in mismatch negativity (MMN), a measure of auditory synaptic plasticity and change-detection. Neuroscience 2020, 32045628. [Google Scholar] [CrossRef] [PubMed]
- Sivarao, D.V.; Frenkel, M.; Chen, P.; Healy, F.L.; Lodge, N.J.; Zaczek, R. MK-801 disrups and nicotine augments 40 Hz auditory steady state responses in the auditory cortex of the urethane-anesthetized rat. Neuropharmacology 2013, 73, 1–9. [Google Scholar] [CrossRef]
- Light, G.A.; Zhang, W.; Joshi, Y.B.; Bhakta, S.; Talledo, J.A.; Swerdlow, N.R. Single-dose memantine improves cortical oscillatory response dynamics in patients with schizophrenia. Neuropsychopharmacology 2017, 42, 2633–2639. [Google Scholar] [CrossRef]
- Molina, J.L.; Voytek, B.; Thomas, M.L.; Joshi, Y.B.; Bhakta, S.G.; Talledo, J.A.; Swerdlow, N.R.; Light, G.A. Memantine effects on electroencephalographic measures of putative excitatory/inhibitory balance in schizophrenia. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 562–568. [Google Scholar] [CrossRef]
- Hesselink, M.B.; De Boer, B.G.; Breimer, D.D.; Danysz, W. Brain penetration and in vivo recovery of NMDA receptor antagonists amantadine and memantine: A quantitative microdialysis study. Pharm. Res. 1999, 16, 637–642. [Google Scholar] [CrossRef]
- Kornhuber, J.; Quack, G. Cerebrospinal fluid and serum concentrations of the N-methyl-D-aspartate (NMDA) receptor antagonist memantine in man. Neurosci. Lett. 1995, 137–139. [Google Scholar] [CrossRef]
- Andrade, C. Memantine as an augmentation treatment for schizophrenia: Limitation of meta-analysis for evidence-based evaluation of research. J. Clin. Psychiatry 2017, 78, e1307–e1309. [Google Scholar] [CrossRef] [Green Version]
- Rammes, G.; Rupprecht, R.; Ferrari, U.; Zieglgänsberger, W.; Parsons, C.G. The N-methyl-D-aspartate receptor channel blockers memantine, MRZ 2/579 and other amino-alkyl-cyclohexanes antagonise 5-HT3 receptor currents in cultured HEK-293 and N1E-115 cell systems in a non-competitive manner. Neurosci. Lett. 2001, 306, 81–84. [Google Scholar] [CrossRef]
- Burke, D.A.; Heshmati, P.; Kholdebarin, E.; Levin, E.D. Decreasing nicotinic receptor activity and the spatial learning impairment caused by the NMDA glutamate antagonist dizocilpine in rats. Eur. J. Pharmacol. 2014, 741, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maskell, P.D.; Speder, P.; Newberry, N.R.; Bermudez, I. Inhibition of human α7 nicotinic acetylcholine receptors by open channel blockers of N-methyl-D-aspartate receptors. Br. J. Pharmacol. 2003, 140, 1313–1319. [Google Scholar] [CrossRef] [Green Version]
- Seeman, P.; Caruso, C.; Lasaga, M. Memantine agonist action at dopamine D2High receptors. Synapse 2008, 62, 149–153. [Google Scholar] [CrossRef]
- Hergovich, N.; Singer, E.; Agneter, E.; Eichler, H.G.; Graselli, U.; Simhandle, C.; Jilma, B. Comparison of the effects of ketamine and memantine on prolactin and cortisol release in men: A randomized, double-blind, placebo-controlled trial. Neuropsychopharmacology 2001, 24, 590–593. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Studies | Number of Trials | Patients | Drugs | Outcomes |
|---|---|---|---|---|
| Kishi et al. 2013 [24] | 8 (n = 406) | Schizophrenia 85.5%, Bipolar disorder 14.5% | Memantine (3 trials, n = 186), Amantadine (5 trials, n = 220) | Not significant compared to placebo: overall symptoms (p = 0.31), positive symptoms (p = 0.44), negative symptoms (p = 0.16), and Clinical Global Impression Severity scale (p = 0.56). Memantine was favorable compared to placebo in Mini-Mental State Examination score in schizophrenia (p = 0.002). |
| Matsuda et al. (Kishi’s group) 2013 [26] | 9 (n = 446) | Schizophrenia, Bipolar disorder | Memantine (4 trials, n = 226), Amantadine (5 trials, n = 220) | Not significant compared to placebo: overall symptoms (p = 0.08), positive symptoms (p = 0.43), and negative symptoms (p = 0.05). Individually, only memantine adjunctive therapy showed a trend towards superior efficacy of overall symptoms (p = 0.06) and negative symptoms (p = 0.06) compared to placebo. |
| Kishi et al. 2017 [27] | 8 (n = 448) | Schizophrenia | Memantine | Significant compared to placebo: negative symptoms (p = 0.006), Positive and Negative Syndrome Scale general subscale (p = 0.002), and Mini-Mental Status Examination score (p < 0.0001). Not significant compared to placebo: overall (p = 0.06), positive (p = 0.07), and depressive symptoms (p = 0.326). Patient age was associated with memantine-associated amelioration of negative symptoms (slope = 0.171, p = 0.0206). The effect on negative symptoms may be greater in younger adult patients. |
| Zheng et al. 2018 [33] | 8 (n = 452) | Schizophrenia | Memantine | Significant compared to placebo: Positive and Negative Syndrome Scale and Brief Psychiatric Rating Scale negative symptoms (p = 0.009) and Mini-Mental State Examination score (p < 0.00001). Not significant compared to placebo: total, positive and general symptoms (p = 0.06–0.60) and Clinical Global Impression Severity Scale (p = 0.78). There was no significant difference in the discontinuation rate (p = 0.31) and adverse drug reactions between the two groups. |
| Zheng et al. 2019 [34] | 15 (n = 988) | Schizophrenia (9 trials, n = 512), Bipolar disorder (3 trials, n = 319), Major depressive disorder (3 trials, n = 157) | Memantine | In schizophrenia, significant compared to placebo: total psychopathology (p = 0.01), negative symptoms (p = 0.0003), and Mini-Mental State Examination score (p < 0.0001). Not significant compared to placebo: positive symptoms (p = 0.05) and general psychopathology (p = 0.14) in schizophrenia, depressive (p = 0.37) and manic (p = 0.09) symptoms in bipolar disorder, and depressive symptoms (p = 0.75) in major depressive disorder. |
| Studies | Patients/ Antipsychotics | Duration | Drug | Number | Outcomes |
|---|---|---|---|---|---|
| Rezaei et al. 2013 [25] | Schizophrenia (risperidone 6 mg/day) | 8 weeks | Memantine (20 mg/day) | Memantine n = 20 Placebo n = 20 | Significant compared to placebo: PANSS negative subscale score (p < 0.001), PANSS total score (p < 0.001), and general psychopathology subscale score (p = 0.002). Not significant compared to placebo: PANSS positive subscale score (p = 0.759). Changes in the Hamilton Depression Rating Scale score and the Extrapyramidal Symptom Rating Scale score and frequency of adverse effects did not differ between the 2 groups. |
| Fakhri et al. 2016 [29] | Schizophrenia (olanzapine 15–20 mg/day) | 6 weeks | Memantine (week 1: 10 mg/day; weeks 2–6: 20 mg/day) | Memantine n = 30 Placebo n = 30 | Significant compared to placebo: PANSS negative subscale score (p < 0.001) and PANSS positive subscale score (p < 0.001). Females showed significantly better response than males, especially in PANSS positive subscale score. No significant changes in extrapyramidal symptoms were observed. |
| Mazinani et al. 2017 [30] | Schizophrenia (risperidone 4–6 mg/day) | 12 weeks | Memantine (titration weeks 1–4: 5 to 20 mg/day; weeks 5–12: 20 mg/day) | Memantine n = 23 Placebo n = 23 | Significant compared to placebo: PANSS negative subscale score (p = 0.003) and Mini-Mental State Examination score (p < 0.001). Not significant compared to placebo: PANSS positive subscale score (p = 0.09) and general psychopathology score (p = 0.9). |
| Omranifard et al. 2017 [31] | Schizophrenia (atypical antipsychotic regimen) | 12 weeks | Memantine (titration weeks 1–4: 5 to 20 mg/day; weeks 5–12: 20 mg/day) | Memantine n = 30 Placebo n = 30 | Significant compared to placebo: PANSS negative subscale score (p = 0.004), PANSS positive subscale score (p = 0.028), general psychopathology score (p < 0.001), depressive symptom score (p < 0.001), and total symptom severity score (p < 0.001). No serious or severe adverse effects were reported during the study. |
| Hassanpour et al. 2019 [32] | Schizophrenia (antipsychotic regimen) | 8 weeks | Memantine (week 1: 10 mg/day; weeks 2–8: 20 mg/day) | Memantine n = 20 Placebo n = 20 | Significant compared to placebo: verbal memory (p = 0.01), working memory (p = 0.007), verbal fluency letter (p = 0.002), and verbal fluency total (p = 0.013) scores of the Brief Assessment of Cognition Scale. No improvement on psychotic symptoms was observed. There was no significant difference in the Abnormal Involuntary Movement Scale and Barnes Akathisia Rating Scale scores between the 2 groups. |
| Schaefer et al. 2020 [35] | Acute schizophrenia with predominant positive symptoms (risperidone 2–8 mg/day) | 6 weeks | Memantine (20 mg/day) | Memantine n = 5 Placebo n = 6 | Significant compared to placebo: attention intensity (p = 0.005), verbal learning (p = 0.050), problem solving (p = 0.043), and flexibility (p = 0.049). Not significant compared to placebo: PANSS total score and PANSS negative subscale score |
| Chronic schizophrenia with negative symptoms (risperidone 2–8 mg/day) | 24 weeks | Memantine (20 mg/day) | Memantine n = 7 Placebo n = 6 | Significant compared to placebo: immediate memory (p = 0.033) and PANSS total score (p = 0.026). Not significant compared to placebo: PANSS negative subscale score. | |
| de Lucena et al. 2009 [38] | Treatment- refractory schizophrenia (mean doses of clozapine memantine: 540 mg/day; placebo: 659 mg/day) | 12 weeks | Memantine (20 mg/day) | Memantine n = 10 Placebo n = 11 | Significant compared to placebo: Brief Psychiatric Rating Scale total score (effect size = −2.75, p = 0.001), positive symptoms score (effect size = −1.38, p = 0.007), negative symptoms score (effect size = −3.33, p = 0.001), Clinical Global Impression score (effect size = −1.56, p = 0.001), and the Mini-Mental State Examination score (effect size = 1.32, p = 0.005). No significant changes in extrapyramidal symptoms were observed. |
| Veerman et al. 2016 [39] | Treatment- refractory schizophrenia (clozapine dosage was remained as much unaltered as possible) | 12 weeks | Memantine (week 1: 10 mg/day; weeks 2–12: 20 mg/day) | Memantine n = 26 Placebo n = 26 (each group has 52 cases based on two cross-overs) | This study was a 26-week single-center, double-blind, placebo-controlled crossover trial. The trial consisted of two crossover, 12-week treatment phases and a placebo wash-out period of 2 weeks in the 13th and 14th week to avoid carryover effects. Significant compared to placebo: PANSS negative subscale score (effect size = 0.29, p = 0.043) and memory composite (effect size = 0.30, p = 0.032), including verbal recognition memory and paired associates learning task scores on the Cambridge Neuropsychological Test Automated Battery. Not significant compared to placebo: PANSS positive subscale score (effect size = 0.15, p = 0.299) and PANSS total score (effect size = 0.19, p = 0.174). Side effects were mild and transient. |
| Omranifard et al. 2015 [41] | Schizophrenia (stable-dose atypical antipsychotic) | 12 weeks | Memantine (titration weeks 1–4: 5 to 20 mg/day; weeks 5–12: 20 mg/day) | Memantine n = 32 Placebo n = 32 | Global Assessment of Functioning scale and quality of life scale scores increased in both groups, with significantly higher scores in the memantine add-on group than the placebo group (p < 0.001 and p < 0.001, respectively). Memantine was well tolerated with no significant side effects. |
| Mode of Actions | Main References |
|---|---|
| Low affinity/fast off-rate | [45,50,51,53] |
| Voltage dependency/fast unblocking kinetics | [50,51,52,53,54,55] |
| Partial trapping | [54,55,56] |
| Uncompetitive antagonism/ agonist concentration dependency | [53,57,58,59,60] |
| Preferential inhibition of extrasynaptic receptors | [65,66,67,68,69,70] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikuchi, T. Is Memantine Effective as an NMDA Receptor Antagonist in Adjunctive Therapy for Schizophrenia? Biomolecules 2020, 10, 1134. https://doi.org/10.3390/biom10081134
Kikuchi T. Is Memantine Effective as an NMDA Receptor Antagonist in Adjunctive Therapy for Schizophrenia? Biomolecules. 2020; 10(8):1134. https://doi.org/10.3390/biom10081134
Chicago/Turabian StyleKikuchi, Tetsuro. 2020. "Is Memantine Effective as an NMDA Receptor Antagonist in Adjunctive Therapy for Schizophrenia?" Biomolecules 10, no. 8: 1134. https://doi.org/10.3390/biom10081134
APA StyleKikuchi, T. (2020). Is Memantine Effective as an NMDA Receptor Antagonist in Adjunctive Therapy for Schizophrenia? Biomolecules, 10(8), 1134. https://doi.org/10.3390/biom10081134