Rapid Ex-Vivo Ciliogenesis and Dose-Dependent Effect of Notch Inhibition on Ciliogenesis of Respiratory Epithelia



Abstract
:1. Introduction
2. Methods
2.1. Isolation of Mouse Trachea and Human Nasal Epithelia
Stationary Phase Culturing
2.2. Suspension Culture for Ciliogenesis
2.3. Assessing Ciliogenesis and Ciliary Motion
2.4. Statistical Analyses
2.5. Results
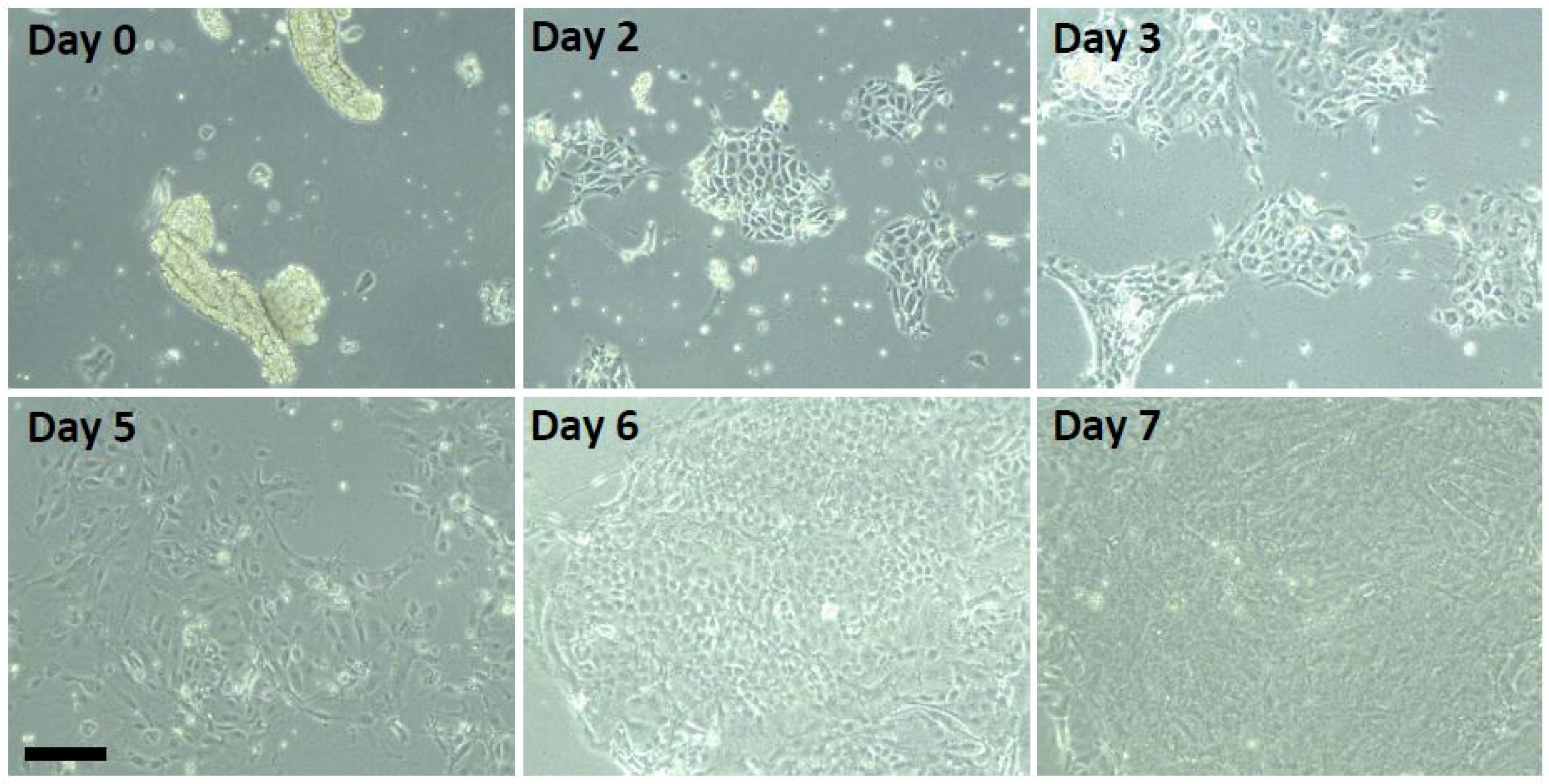
2.5.1. A Suspension Culture Method for Measuring Mouse Airway Cell Reciliation
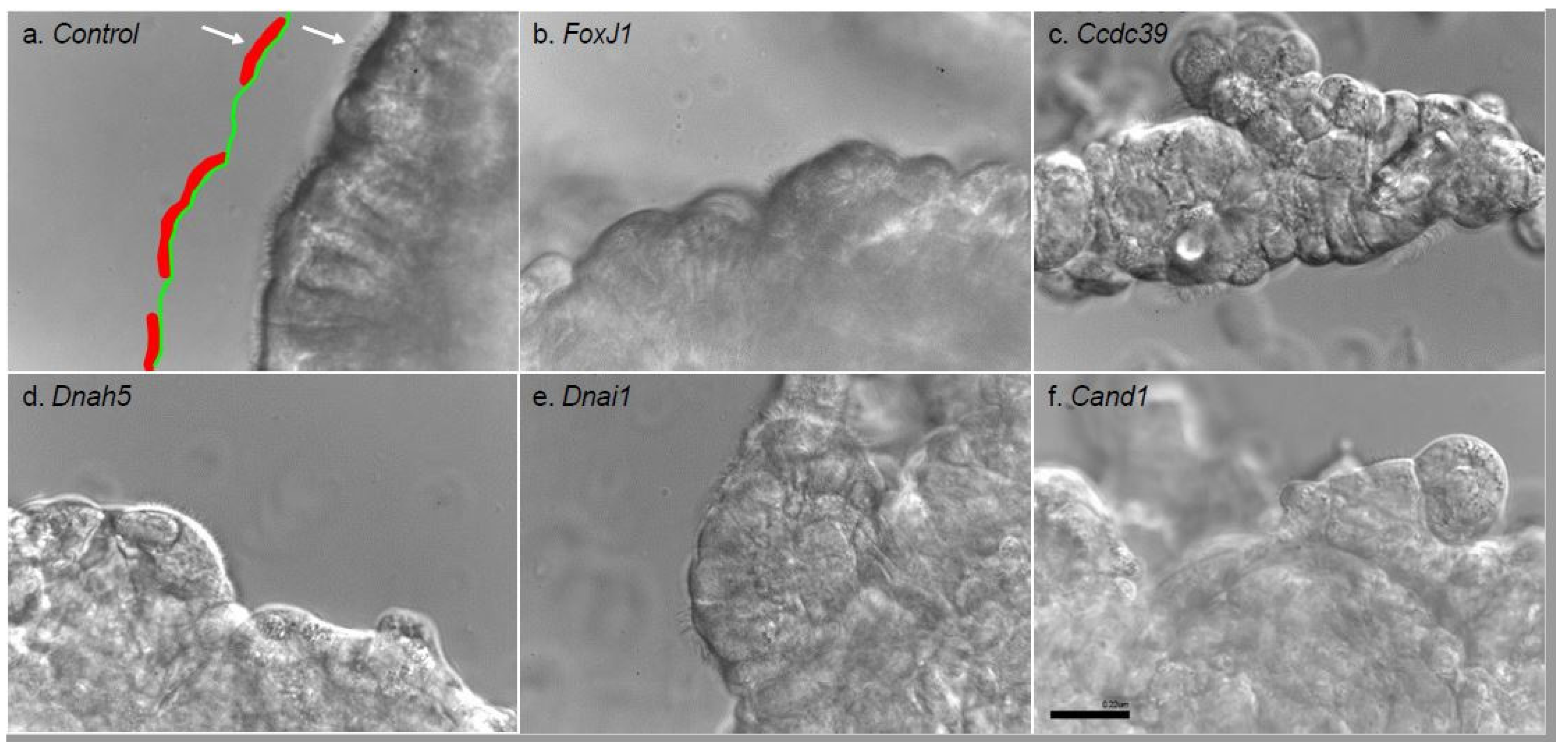
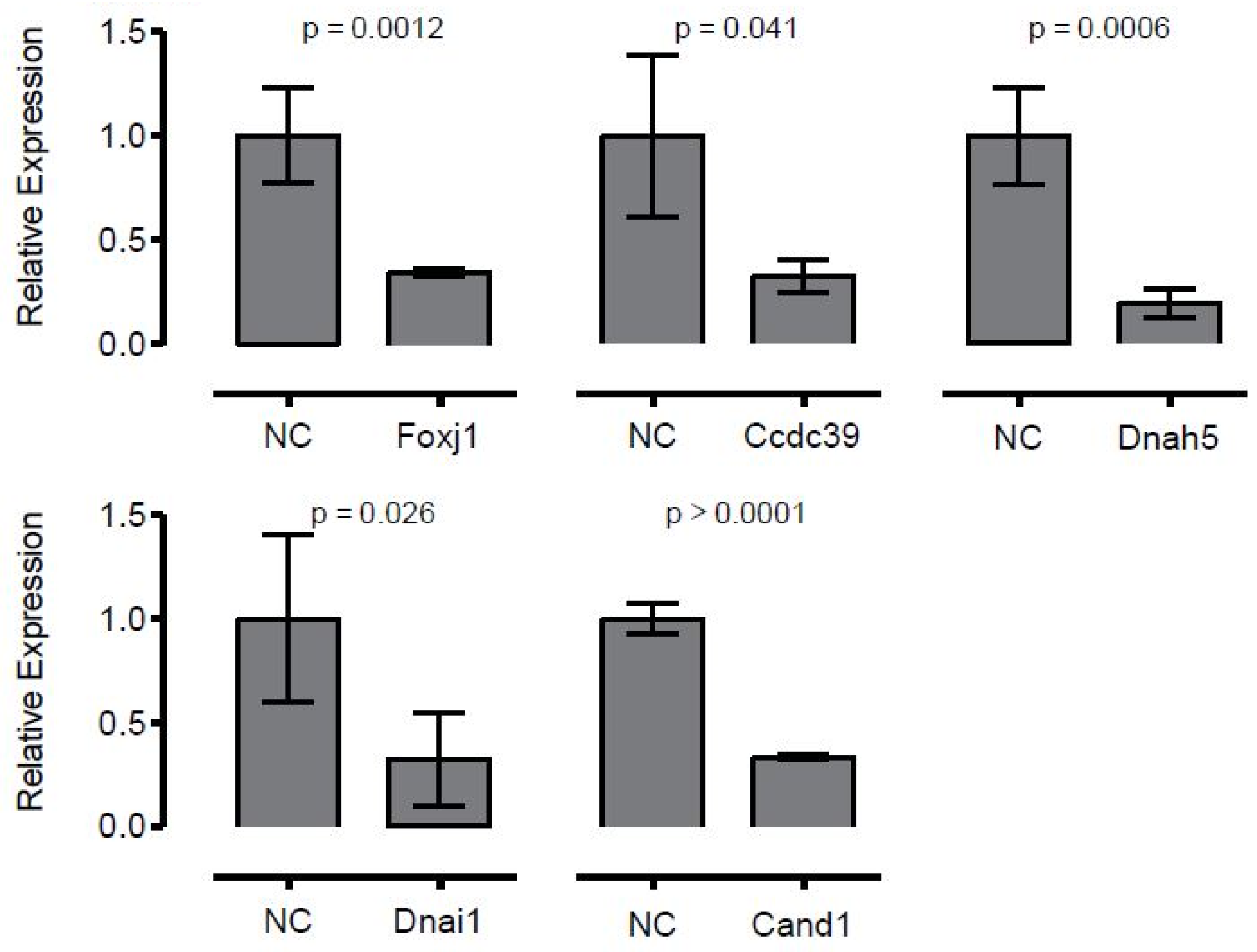
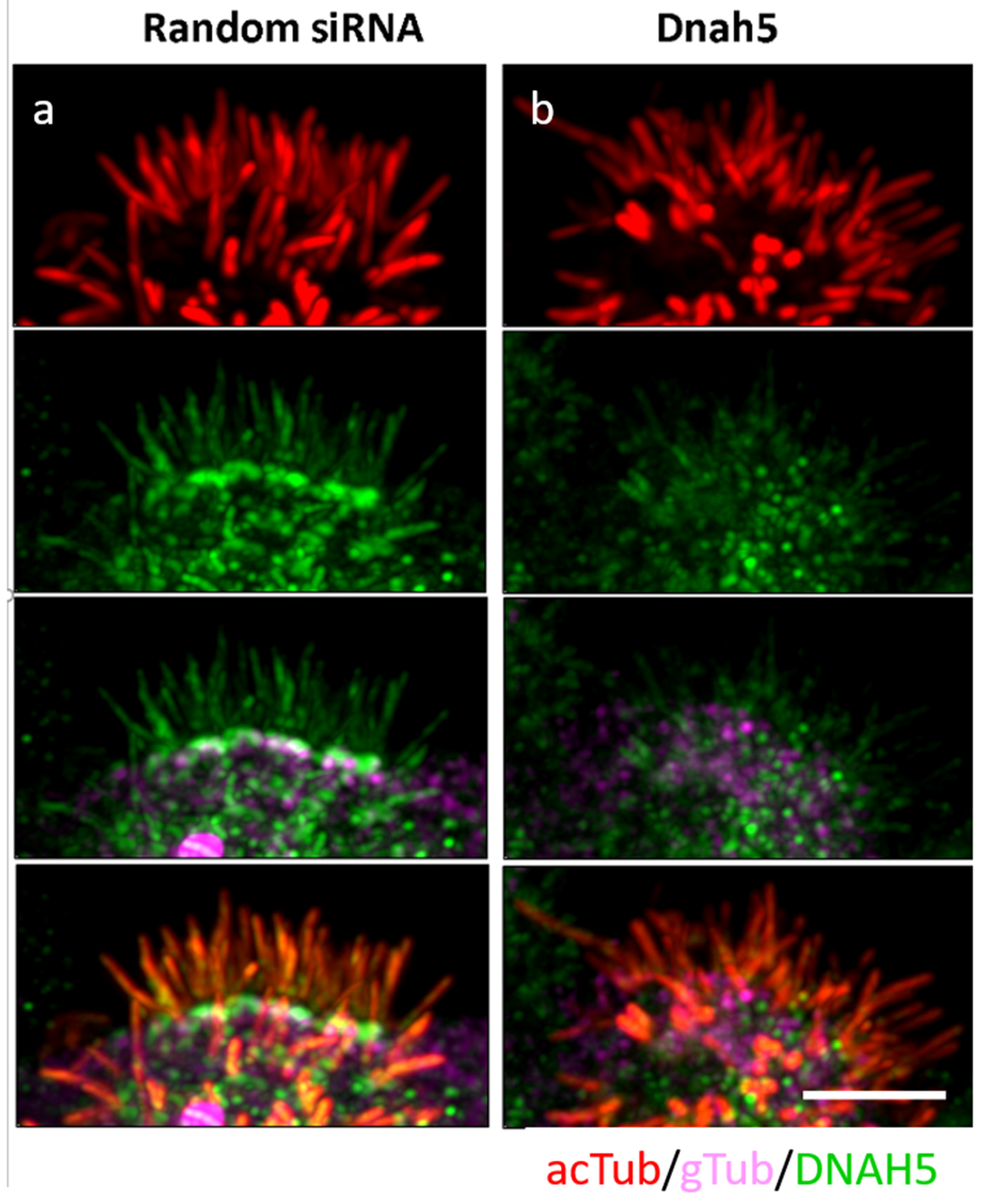
2.5.2. Evaluation of Genetic Contribution to Reciliation through siRNA Knockdown
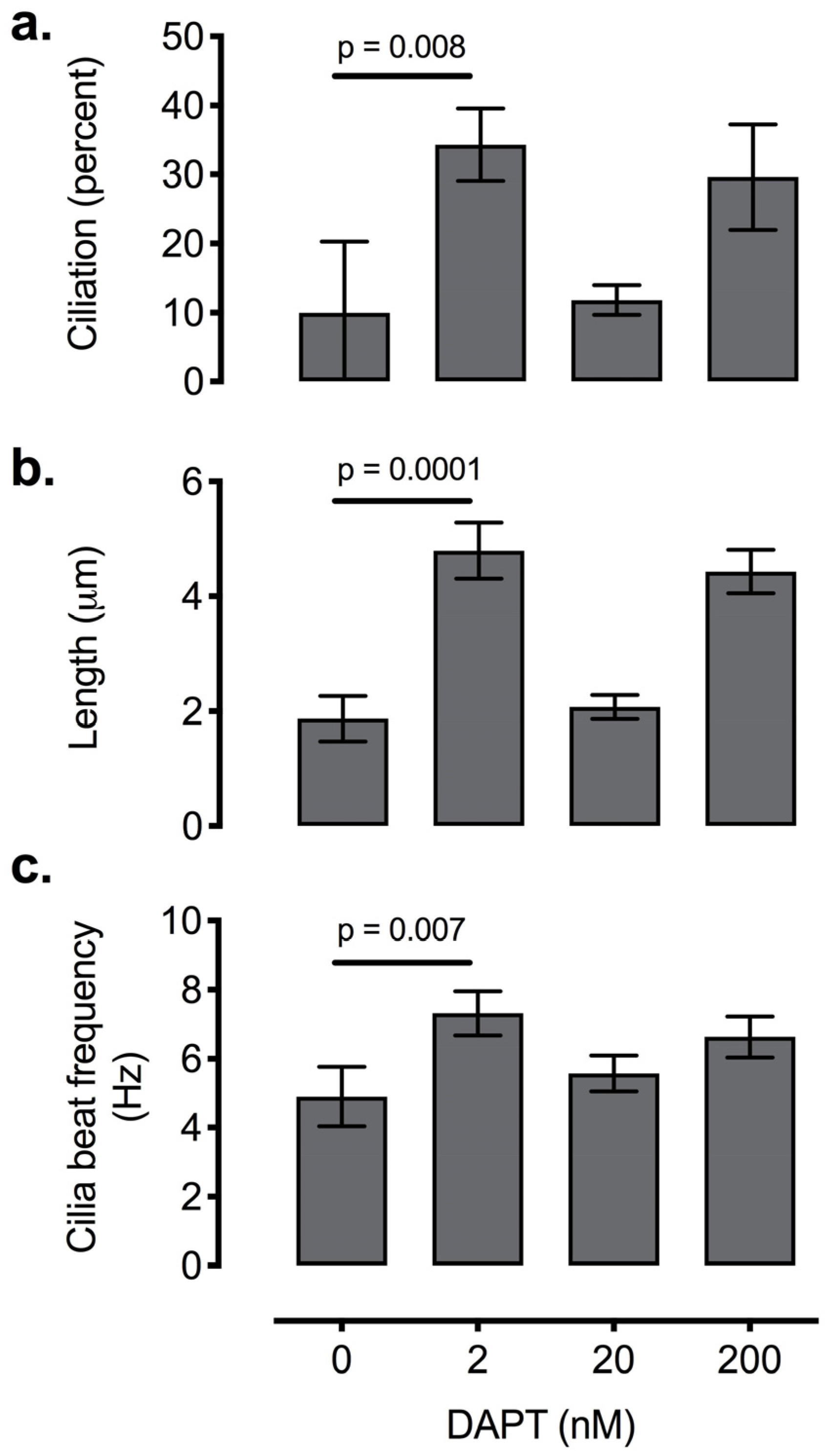
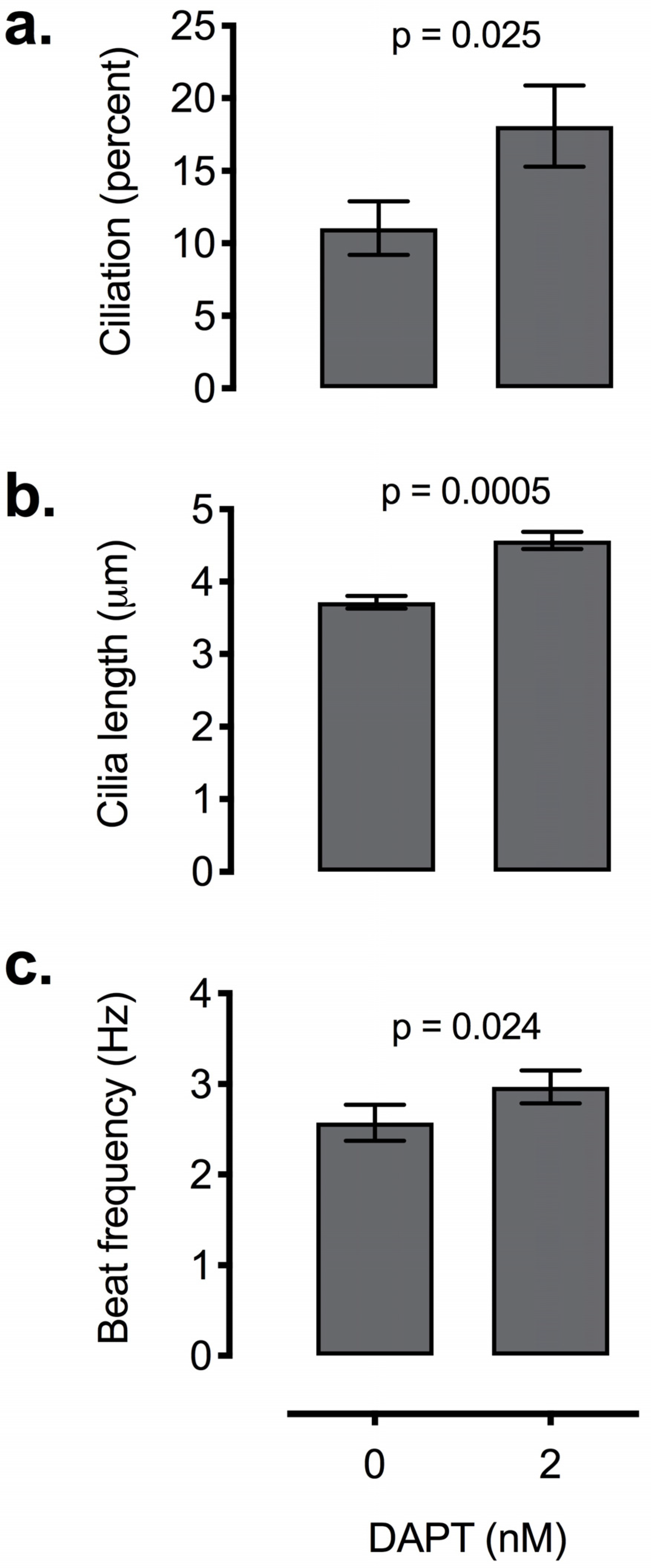
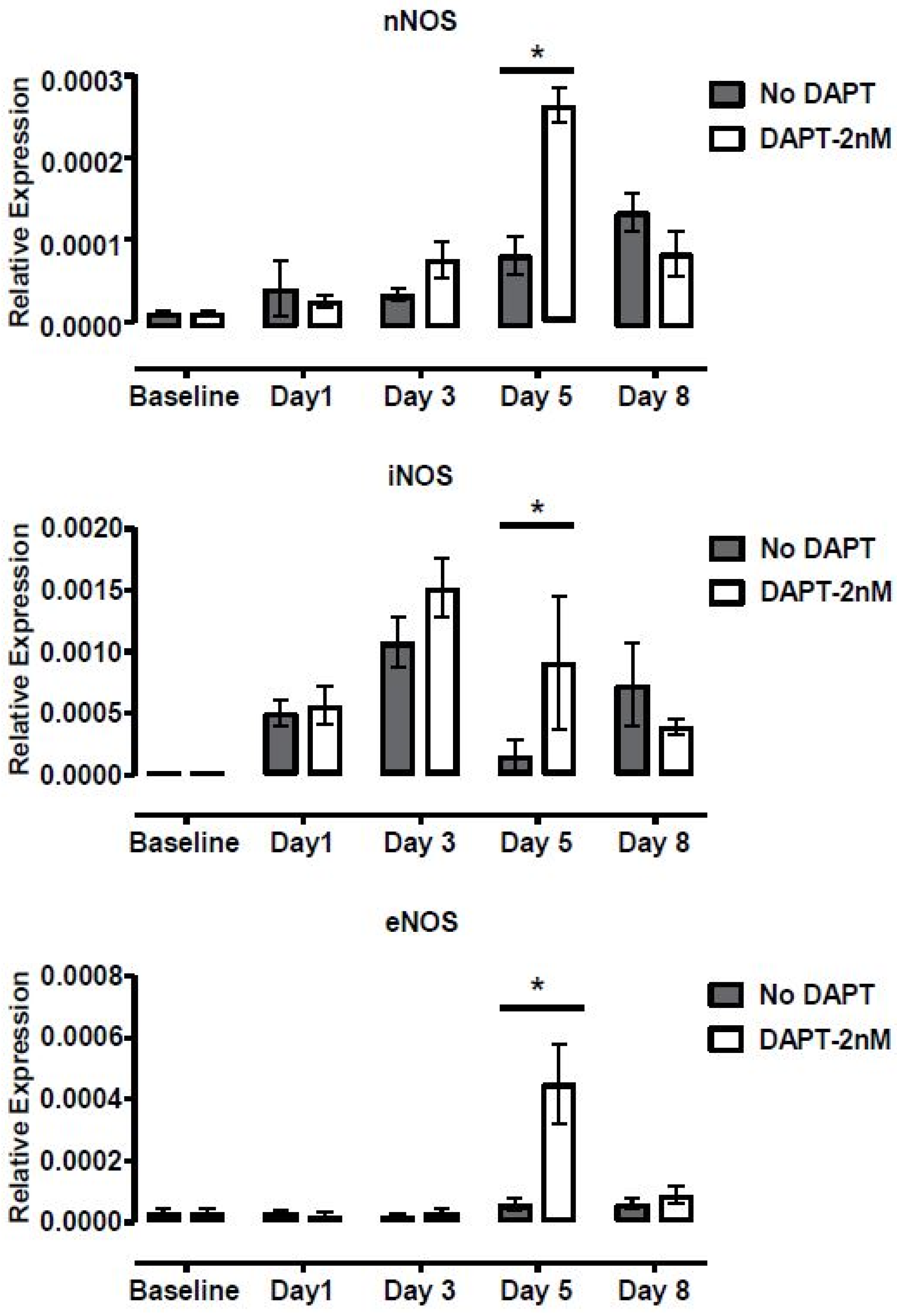
2.5.3. Enhancing Ciliogenesis with Notch Inhibition
3. Discussion
4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hessel, J.; Heldrich, J.; Fuller, J.; Staudt, M.R.; Radisch, S.; Hollmann, C.; Harvey, B.-G.; Kaner, R.J.; Salit, J.; Yee-Levin, J.; et al. Intraflagellar Transport Gene Expression Associated with Short Cilia in Smoking and COPD. PLoS ONE 2014, 9, e85453. [Google Scholar] [CrossRef] [Green Version]
- Yaghi, A.; Zaman, A.; Cox, G.; Dolovich, M.B. Ciliary beating is depressed in nasal cilia from chronic obstructive pulmonary disease subjects. Respir. Med. 2012, 106, 1139–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedzicha, J.A.; Singh, R.; Mackay, A.J. Acute COPD exacerbations. Clin. Chest Med. 2014, 35, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M.; Rogliani, P.; Puxeddu, E.; Ora, J.; Matera, M.G. An overview of the current management of chronic obstructive pulmonary disease: Can we go beyond the GOLD recommendations? Expert Rev. Respir. Med. 2017, 12, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Mathers, C.D.; Loncar, D. Projections of Global Mortality and Burden of Disease from 2002 to 2030. PLoS Med. 2006, 3, e442. [Google Scholar] [CrossRef] [Green Version]
- McLean, S.; Hoogendoorn, M.; Hoogenveen, R.T.; Feenstra, T.L.; Wild, S.; Simpson, C.R.; Mölken, M.R.-V.; Sheikh, A. Projecting the COPD population and costs in England and Scotland: 2011 to 2030. Sci. Rep. 2016, 6, 31893. [Google Scholar] [CrossRef] [Green Version]
- Hoogendoorn, M.; Mölken, M.R.-V.; Hoogenveen, R.T.; Al, M.J.; Feenstra, T.L. Developing and Applying a Stochastic Dynamic Population Model for Chronic Obstructive Pulmonary Disease. Value Health 2011, 14, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Garrod, A.S.; Madan-Khetarpal, S.; Sreedher, G.; McGuire, M.; Yagi, H.; Klena, N.T.; Gabriel, G.C.; Khalifa, O.; Zahid, M.; et al. Respiratory motile cilia dysfunction in a patient with cranioectodermal dysplasia. Am. J. Med. Genet. Part A 2015, 167, 2188–2196. [Google Scholar] [CrossRef]
- Frank, V.; Hollander, A.I.D.; Brüchle, N.O.; Zonneveld, M.N.; Nürnberg, G.; Becker, C.; Du Bois, G.; Kendziorra, H.; Roosing, S.; Senderek, J.; et al. Mutations of theCEP290 gene encoding a centrosomal protein cause Meckel-Gruber syndrome. Hum. Mutat. 2008, 29, 45–52. [Google Scholar] [CrossRef]
- Garrod, A.S.; Zahid, M.; Tian, X.; Francis, R.O.; Khalifa, O.; Devine, W.; Gabriel, G.C.; Leatherbury, L.; Lo, C.W. Airway Ciliary Dysfunction and Sinopulmonary Symptoms in Patients with Congenital Heart Disease. Ann. Am. Thorac. Soc. 2014, 11, 1426–1432. [Google Scholar] [CrossRef]
- Zahid, M.; Bais, A.; Tian, X.; Devine, W.; Lee, D.M.; Yau, C.; Sonnenberg, D.; Beerman, L.; Khalifa, O.; Lo, C.W. Airway ciliary dysfunction and respiratory symptoms in patients with transposition of the great arteries. PLoS ONE 2018, 13, e0191605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trost, M.J.; Shimada, H. Hospitalists Should Have Increased Suspicion for Primary Ciliary Dyskinesia in Patients with Congenital Heart Disease. Hosp. Pediatr. 2014, 4, 109–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swisher, M.; Jonas, R.; Tian, X.; Lee, E.S.; Lo, C.W.; Leatherbury, L. Increased postoperative and respiratory complications in patients with congenital heart disease associated with heterotaxy. J. Thorac. Cardiovasc. Surg. 2011, 141, 637–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, E.; Adams, P.S.; Tian, X.; Khalifa, O.; Wearden, P.; Zahid, M.; Lo, C. Airway ciliary dysfunction: Association with adverse postoperative outcomes in nonheterotaxy congenital heart disease patients. J. Thorac. Cardiovasc. Surg. 2018, 155, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Jorissen, M.; Van Der Schueren, B.; Berghe, H.V.D.; Cassiman, J.-J. The preservation and regeneration of cilia on human nasal epithelial cells cultured in vitro. Eur. Arch. Otorhinolaryngol. 1989, 246, 308–314. [Google Scholar] [CrossRef]
- Jorissen, M.; Bessems, A. Normal Ciliary Beat Frequency after Ciliogenesis in Nasal Epithelial Cells Cultured Sequentially as Monolayer and in Suspension. Acta Otolaryngol. 1995, 115, 66–70. [Google Scholar] [CrossRef]
- Jorissen, M.; Van Der Schueren, B.; Tyberghein, J.; Van Der Berghe, H.; Cassiman, J.J. Ciliogenesis and coordinated ciliary beating in human nasal epithelial cells cultured in vitro. Acta Otolaryngol. Belg. 1989, 43, 67–73. [Google Scholar]
- Schindelin, J. Fiji: An open-source platform for biological-image analysis. Nat. Method. 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Doube, M.; Kłosowski, M.M.; Arganda-Carreras, I.; Cordelieres, F.P.; Dougherty, R.P.; Jackson, J.S.; Schmid, B.; Hutchinson, J.R.; Shefelbine, S.J. BoneJ: Free and extensible bone image analysis in Image. J. Bone 2010, 47, 1076–1079. [Google Scholar] [CrossRef] [Green Version]
- Strutz, F.; Okada, H.; Lo, C.W.; Danoff, T.; Carone, R.L.; Tomaszewski, J.E.; Neilson, E.G. Identification and characterization of a fibroblast marker: FSP1. J. Cell Boil. 1995, 130, 393–405. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Huang, T.; Richer, E.J.; Schmidt, J.-E.H.; Zabner, J.; Borok, Z.; Brody, S.L. Role of f-box factor foxj1 in differentiation of ciliated airway epithelial cells. Am. J. Physiol. Cell. Mol. Physiol. 2004, 286, L650–L657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merveille, A.; Davis, E.E.; Becker-Heck, A.; Legendre, M.; Amirav, I.; Bataille, G.; Belmont, J.W.; Beydon, N.; Billen, F.; Clément, A.; et al. CCDC39 is required for assembly of inner dynein arms and the dynein regulatory complex and for normal ciliary motility in humans and dogs. Nat. Genet. 2010, 43, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, N.; Hohenfellner, M.; Duensing, S. CAND1 Promotes PLK4-Mediated Centriole Overduplication and Is Frequently Disrupted in Prostate Cancer. Neoplasia 2012, 14, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Guseh, J.S.; Bores, S.A.; Stanger, B.Z.; Zhou, Q.; Anderson, W.J.; Melton, U.A.; Rajagopal, J. Notch signaling promotes airway mucous metaplasia and inhibits alveolar development. Development 2009, 136, 1751–1759. [Google Scholar] [CrossRef] [Green Version]
- Tsao, P.-N.; Vasconcelos, M.; Izvolsky, K.I.; Qian, J.; Lu, J.; Cardoso, W.V. Notch signaling controls the balance of ciliated and secretory cell fates in developing airways. Development 2009, 136, 2297–2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eenjes, E.; Mertens, T.C.J.; Kempen, M.J.B.-V.; Van Wijck, Y.; Taube, C.; Rottier, R.J.; Hiemstra, P.S. A novel method for expansion and differentiation of mouse tracheal epithelial cells in culture. Sci. Rep. 2018, 8, 7349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerovac, B.J.; Valencia, M.; Baumlin, N.; Salathe, M.; Conner, G.E.; Fregien, N.L. Submersion and Hypoxia Inhibit Ciliated Cell Differentiation in a Notch-Dependent Manner. Am. J. Respir. Cell Mol. Boil. 2014, 51, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Collaborators, T.U.B.O.D.; Mokdad, A.H.; Ballestros, K.; Echko, M.; Glenn, S.; Olsen, H.E.; Mullany, E.; Lee, A.; Khan, A.R.; Ahmadi, A.; et al. The State of US Health, 1990–2016: Burden of Diseases, Injuries, and Risk Factors Among US States. JAMA 2018, 319, 1444–1472. [Google Scholar] [CrossRef] [Green Version]
- Lam, H.C.; Cloonan, S.M.; Bhashyam, A.R.; Haspel, J.A.; Singh, A.; Sathirapongsasuti, J.F.; Cervo, M.; Yao, H.; Chung, A.L.; Mizumura, K.; et al. Histone deacetylase 6–mediated selective autophagy regulates COPD-associated cilia dysfunction. J. Clin. Investig. 2013, 123, 5212–5230. [Google Scholar] [CrossRef]
- Boon, M.; Wallmeier, J.; Ma, L.; Loges, N.T.; Jaspers, M.; Olbrich, H.; Dougherty, G.W.; Raidt, J.; Werner, C.; Amirav, I.; et al. MCIDAS mutations result in a mucociliary clearance disorder with reduced generation of multiple motile cilia. Nat. Commun. 2014, 5, 4418. [Google Scholar] [CrossRef]
- Willems, T.; Jorissen, M. Sequential monolayer-suspension culture of human airway epithelial cells. J. Cyst. Fibros. 2004, 3, 53–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Brody, S.L. Culture and differentiation of mouse tracheal epithelial cells. Method. Mol. Biol. 2013, 945, 123–143. [Google Scholar]
- Horani, A.; Dickinson, J.D.; Brody, S.L. Applications of Mouse Airway Epithelial Cell Culture for Asthma Research. Method. Mol. Biol. 2013, 1032, 91–107. [Google Scholar]
- Vladar, E.; Brody, S.L. Analysis of Ciliogenesis in Primary Culture Mouse Tracheal Epithelial Cells. Method. Enzymol. 2013, 525, 285–309. [Google Scholar]
- Radder, J.E.; Gregory, A.D.; Leme, A.S.; Cho, M.H.; Chu, Y.; Kelly, N.J.; Bakke, P.; Gulsvik, A.; Litonjua, A.A.; Sparrow, D.; et al. Variable Susceptibility to Cigarette Smoke–Induced Emphysema in 34 Inbred Strains of Mice Implicates Abi3bp in Emphysema Susceptibility. Am. J. Respir. Cell Mol. Boil. 2017, 57, 367–375. [Google Scholar] [CrossRef]
- Karp, P.H.; Moninger, T.O.; Weber, S.P.; Nesselhauf, T.S.; Launspach, J.L.; Zabner, J.; Welsh, M.J.; Wise, C. An in vitro Model of Differentiated Human Airway Epithelia: Methods for Establishing Primary Cultures. Method. Mol. Biol. 2002, 188, 115–137. [Google Scholar]
- You, Y.; Richer, E.J.; Huang, T.; Brody, S.L. Growth and differentiation of mouse tracheal epithelial cells: Selection of a proliferative population. Am. J. Physiol. Cell. Mol. Physiol. 2002, 283. [Google Scholar] [CrossRef]
- Gras, D.; Petit, A.; Charriot, J.; Knabe, L.; Alagha, K.; Gamez, A.S.; Garulli, C.; Bourdin, A.; Chanez, P.; Molinari, N.; et al. Epithelial ciliated beating cells essential for ex vivo ALI culture growth. BMC Pulm. Med. 2017, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Olbrich, H.; Häffner, K.; Kispert, A.; Völkel, A.; Volz, A.; Sasmaz, G.; Reinhardt, R.; Hennig, S.; Lehrach, H.; Konietzko, N.; et al. Mutations in DNAH5 cause primary ciliary dyskinesia and randomization of left–right asymmetry. Nat. Genet. 2002, 30, 143–144. [Google Scholar] [CrossRef]
- Guichard, C.; Harricane, M.-C.; Lafitte, J.-J.; Godard, P.; Zaegel, M.; Tack, V.; Lalau, G.; Bouvagnet, P. Axonemal Dynein Intermediate-Chain Gene (DNAI1) Mutations Result in Situs Inversus and Primary Ciliary Dyskinesia (Kartagener Syndrome). Am. J. Hum. Genet. 2001, 68, 1030–1035. [Google Scholar] [CrossRef] [Green Version]
- Francis, R.O.; Chatterjee, B.; Loges, N.T.; Zentgraf, H.; Omran, H.; Lo, C.W. Initiation and maturation of cilia-generated flow in newborn and postnatal mouse airway. Am. J. Physiol. Cell. Mol. Physiol. 2009, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchon, S. Delineation of CCDC39/CCDC40 mutation spectrum and associated phenotypes in primary ciliary dyskinesia. J. Med. Genet. 2012, 49, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Antony, D.; Becker-Heck, A.; Zariwala, M.A.; Schmidts, M.; Onoufriadis, A.; Forouhan, M.; Wilson, R.; Taylor-Cox, T.; Dewar, A.; Jackson, C.L.; et al. Mutations in CCDC39 and CCDC40 are the major cause of primary ciliary dyskinesia with axonemal disorganization and absent inner dynein arms. Hum. Mutat. 2013, 34, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhamed, Z.; Vuong, S.M.; Hill, L.; Shula, C.; Timms, A.; Beier, D.; Campbell, K.; Mangano, F.T.; Stottmann, R.W.; Goto, J. A mutation in Ccdc39 causes neonatal hydrocephalus with abnormal motile cilia development in mice. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Lafkas, D.; Shelton, A.; Chiu, C.; Boenig, G.D.L.; Chen, Y.; Stawicki, S.S.; Siltanen, C.; Reichelt, M.; Zhou, M.; Wu, X.; et al. Therapeutic antibodies reveal Notch control of transdifferentiation in the adult lung. Nature 2015, 528, 127–131. [Google Scholar] [CrossRef]
- Annaert, W.G. Presenilin 1 controls gamma-secretase processing of amyloid precursor protein in pre-golgi compartments of hippocampal neurons. J. Cell Biol. 1999, 147, 277–294. [Google Scholar] [CrossRef] [Green Version]
- McCauley, K.; Hawkins, F.; Serra, M.; Thomas, D.C.; Jacob, A.; Kotton, D.N. Efficient Derivation of Functional Human Airway Epithelium from Pluripotent Stem Cells via Temporal Regulation of Wnt Signaling. Cell Stem Cell 2017, 20, 844–857. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Shu, B.; Yang, R.; Xu, Y.; Xing, B.; Liu, J.; Chen, L.; Qi, S.; Liu, X.; Wang, P.; et al. Wnt and Notch signaling pathway involved in wound healing by targeting c-Myc and Hes1 separately. Stem Cell Res. Ther. 2015, 6, 120. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, A.J.; Josephson, M.; Rosenfeld, M.; Yilmaz, O.; Davis, S.D.; Polineni, D.; Guadagno, E.; Leigh, M.W.; Lavergne, V. Accuracy of Nasal Nitric Oxide Measurement as a Diagnostic Test for Primary Ciliary Dyskinesia. A Systematic Review and Meta-analysis. Ann. Am. Thorac. Soc. 2017, 14, 1184–1196. [Google Scholar] [CrossRef] [Green Version]
- Charles, N.; Ozawa, T.; Squatrito, M.; Bleau, A.-M.; Brennan, C.W.; Hambardzumyan, L.; Holland, E.C. Perivascular nitric oxide activates notch signaling and promotes stem-like character in PDGF-induced glioma cells. Cell Stem Cell 2010, 6, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-Y.; Park, J.-H.; Mo, J.-S.; Ann, E.-J.; Han, S.-O.; Baek, S.-H.; Kim, K.-J.; Im, S.-Y.; Choi, E.-J.; Park, H.-S.; et al. Downregulation by lipopolysaccharide of Notch signaling, via nitric oxide. J. Cell Sci. 2008, 121, 1466–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welchering, N.; Ochoa, S.; Tian, X.; Francis, R.O.; Zahid, M.; Lo, C.; Muñoz, R. Dexmedetomidine and Fentanyl Exhibit Temperature Dependent Effects on Human Respiratory Cilia. Front. Pediatr. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopher, A.B.; Ochoa, S.; Krushansky, E.; Francis, R.O.; Tian, X.; Zahid, M.; Lo, C.; Muñoz, R. The Effects of Temperature and Anesthetic Agents on Ciliary Function in Murine Respiratory Epithelia. Front. Pediatr. 2014, 2, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, B.M.; Tsifansky, M.D.; Yang, Y.; Yeo, Y. Challenges and advances in the development of inhalable drug formulations for cystic fibrosis lung disease. Expert Opin. Drug Deliv. 2011, 8, 451–466. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Knockdown | Ciliation (%) | Cilia Beat Frequency (Beats/sec) | Cilia Length (μm) | Cilia Motion |
|---|---|---|---|---|
| Control | 16.3 ± 8.7 | 7.4 ± 1.1 | 2.6 ± 0.7 | Normal |
| Foxj1 | 2.4 ± 3.4 * | 7.0 ± 0.6 | 3.3 ± 0.2 | Mostly immotile |
| Ccdc39 | 3.8 ± 7.6 * | 0 | 1.5 | Immotile |
| Dnah5 | 15.0 ± 8.2 | 7.1 ± 1.1 | 3.2 ± 1.1 | 50% immotile |
| Dnai1 | 10.3 ± 4.0 | 7.2 ± 1.0 | 2.1 ± 0.7 | 50% immotile |
| Cand1 | 4.0 ± 5.6 * | 7.6 ± 1.4 | 1.7 ± 0.3 | Normal |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahid, M.; Feinstein, T.N.; Oro, A.; Schwartz, M.; Lee, A.D.; Lo, C.W. Rapid Ex-Vivo Ciliogenesis and Dose-Dependent Effect of Notch Inhibition on Ciliogenesis of Respiratory Epithelia. Biomolecules 2020, 10, 1182. https://doi.org/10.3390/biom10081182
Zahid M, Feinstein TN, Oro A, Schwartz M, Lee AD, Lo CW. Rapid Ex-Vivo Ciliogenesis and Dose-Dependent Effect of Notch Inhibition on Ciliogenesis of Respiratory Epithelia. Biomolecules. 2020; 10(8):1182. https://doi.org/10.3390/biom10081182
Chicago/Turabian StyleZahid, Maliha, Timothy N. Feinstein, Anthony Oro, Molly Schwartz, Alex D. Lee, and Cecilia W. Lo. 2020. "Rapid Ex-Vivo Ciliogenesis and Dose-Dependent Effect of Notch Inhibition on Ciliogenesis of Respiratory Epithelia" Biomolecules 10, no. 8: 1182. https://doi.org/10.3390/biom10081182
APA StyleZahid, M., Feinstein, T. N., Oro, A., Schwartz, M., Lee, A. D., & Lo, C. W. (2020). Rapid Ex-Vivo Ciliogenesis and Dose-Dependent Effect of Notch Inhibition on Ciliogenesis of Respiratory Epithelia. Biomolecules, 10(8), 1182. https://doi.org/10.3390/biom10081182